










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Mesoamericana
On-line version ISSN 2215-3608Print version ISSN 1659-1321
Agron. Mesoam vol.21 n.1 San Pedro Jun. 2010
Comportamiento reproductivo de vacas criollas con amamantamiento restringido y sincronización del estro 1
Juan Prisciliano Zárate-Martínez2, José Alejandro Ramírez-Godínez3, Felipe Alonso Rodríguez-Almeida3
1 Proyecto de Investigación del Rescate de Bovinos Criollos.
2 Campo Experimental La Posta. Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP). Km 22.5 carretera Veracruz- Córdoba, Col. Paso del Toro, mpio. de Medellín de Bravo, Veracruz, México. Apdo. Postal 429, 91700, Veracruz, Ver., México. zarate.juan@inifap.gob.mx
3 Departamento de Postgrado. Facultad de Zootecnia. Universidad Autónoma de Chihuahua Periférico Francisco. R. Almada km. 1 Chihuahua,Chih., México. aramirez@uach.mx; frodrigu@uach.mx
Resumen
Comportamiento reproductivo de vacas criollas con amamantamiento restringido y sincronización del estro. Los objetivos fueron conocer el comportamiento reproductivo posparto de vacas criollas de rodeo con amamantamiento restringido, la respuesta a la sincronización del estro con protocolos hormonales, los niveles séricos de progesterona y estradiol, la actividad ovárica y la tasa de concepción, con el empleo de inseminación artificial a tiempo fijo. En el Municipio de Soto Maynes, Chihuahua, México durante el verano de 2008, se observó el comportamiento de 27 vacas amamantando durante 95 días y se determinó el inicio de su actividad ovárica posparto por detección de estros, ultrasonografía y niveles séricos de progesterona. En el primer tratamiento, once vacas recibieron un dispositivo intravaginal con 1,9 g de progesterona, inyección intramuscular con 1 mg de β-estradiol + 50 mg de P4. Se retiró el dispositivo intravaginal el día siete más inyección intramuscular de 30 mg de prostaglandina. A las 24 h se aplicó 1 mg de β- estradiol y la IATF se realizó a las 54 h posteriores al retiro del dispositivo intravaginal. El segundo tratamiento con once vacas fue similar, pero la inyección intramuscular al recibir el dispositivo intravaginal sólo contenía β-estradiol. Se observó un comportamiento agresivo durante el amamantamiento restringido. El reinicio de actividad ovárica, fue a los 11,66 días de haber expuesto a vacas criollas a amamantamiento restringido, 11,11% mostraron estro durante este periodo, todas presentaron estro después de los protocolos hormonales. Hubo interacción T*DS (P=0,06) para niveles séricos de progesterona y estradiol. La tasa de concepción fue de 18,8%. Adicionar progesterona con un dispositivo intravaginal no incrementaron los niveles séricos de progesterona.
Palabras clave: Bovinos, progesterona, estradiol, cuerpo lúteo.
Abstract
Reproductive performance of Creole cows with restricted suckling and estrus synchronization. The objectives were to study the reproductive performance, initiation of post-partum ovarian activity, response to restricted suckling, estrus synchronization, and conception rate using artificial insemination at a fixed time of Creole rodeo cows (C). In Chihuahua, México, during the summer 2008, twenty-seven cows were used over 95 d, and the beginning of post-partum ovarian activity was detected by ultrasonography. Hormone treatment for estrus synchronization was administered to 22 cows (n=11 per group). In treatment1 (T1) the cows (n=11) received a CIDR with 1,9 g of progesterone and one IM injection with 1 mg of β-estradiol + 50 mg of P4. The CIDR was removed after d 7 and an IM injection of 30 mg of PGF2α was applie d. After 24 h, 1 mg of β-estradiol was applied, and the cows were inseminated 54 h after removing the CIDR. T2 (n=11) was similar, but the IM injection contained β-estradiol only. The C showed aggressive behavior (AB) during the restricted suckling. The average days from the beginning of the ovarian activity were 11,66 d and only 11,11% of the cows exhibited estrus during initial ovarian activity. One hundred percent of the animals underwent estrus on d 9 post synchronization, there were not differences (P>0,05) among the levels of progesterone of the T1 (4,94±0,331 ng/ml) vs the T2 (4,87±0,30 ng/ml. There was a T*DS interaction for concentrations of progesterone and E2 (P=0.06). There were no differences (P>0.05) in conception rate (18,8%). Adding P4 with a CIDR did not increase the levels of progesterone in C.
Key words: Bovine, progesterone, estradiol, corpora lutea.
Introducción
El ganado bovino criollo de rodeo es descendiente del ganado traído de la península Ibérica a América en el siglo XVI y a lo largo de 500 años se adaptó su constitución genética a las distintas condiciones ambientales. Originándose como consecuencia poblaciones que se caracterizan por su adaptación y calidad biológica, como son, la gran resistencia a enfermedades y facilidad de parto entre otras, que le han dado su adaptación a las condiciones ambientales preponderantes de las distintas regiones del norte de México donde se desarrollaron. Sin embargo, por considerarlas poco productivas, las poblaciones de animales llamadas criollas, están siendo poco a poco sustituidas o absorbidas por otras razas. Como en el caso del ganado criollo de rodeo, que son los preferidos para las suertes de lazo (deporte de rodeo, donde son utilizados para el lazo en equipo y derribarlos en la lucha con el novillo) por su longevidad, bajo tamaño, fortaleza y crecimiento rápido de cuernos. El problema en el diseño de estrategias de conservación y mejoramiento genético para el ganado criollo de rodeo, es que las vacas no responden favorablemente a los protocolos hormonales de sincronización del estro para poder utilizarlos comercialmente y realizar programas de inseminación artificial (IA) (Ramírez et al. 1998).
El estudio del comportamiento social jerárquico reproductivo durante el inicio de la actividad ovárica del ciclo estral de animales que no están acostumbrados a un manejo constante, puede esclarecer los fenómenos que interfieren en la sincronización de su estro, ya que el conocimiento de la conducta social durante el amamantamiento y la jerarquía social del ganado criollo de rodeo que vive bajo condiciones muy rústicas y de muy poco manejo.
Se desconoce el comportamiento reproductivo posparto del ganado criollo de rodeo bajo manejo de amamantamiento restringido y su respuesta a los protocolos hormonales de sincronización del estro que incluyen progesterona (P4) natural. Con este conocimiento se podrían elucidar mejor los factores que afectan y controlan la dinámica folicular y con el control de los eventos periovulatorios lograr producir la sincronización de un estro fértil, utilizando inseminación artificial.
En el ganado comercial productor de carne o leche, además de someterlos a amamantamiento restringido, se realizan tratamientos hormonales que incluyen aplicaciones o periodos de suplementación con progesterona. Los animales bajo tratamiento han mostrado estros con una fase lútea subsecuente de duración normal y tasa de preñez mejorada, comparados con los no tratados (Rhodes et al. 2003). Sin embargo, las respuestas a cualquier tratamiento son variables y también están relacionadas con los demás factores que influyen en la duración del intervalo entre partos (Rhodes et al. 2003).
Es necesario reducir en vacas criollas de rodeo, los factores que influyen negativamente sobre los tratamientos hormonales de la sincronización del estro, tales como el reinicio tardío de la actividad ovárica causado por el amamantamiento y presencia del becerro las 24 horas del día, una inadecuada detección de estros que en el caso de los animales asilvestrados es mucho más factible que se presente este problema ya que no se conocen sus hábitos y/o comportamiento reproductivos tales como la frecuencia y horas del día en que muestran su estro (monta), así como los niveles séricos de progesterona y estradiol (E2) producidos por el tratamiento hormonal de sincronización del estro utilizado ya que pueden presentarse niveles subluteales de progesterona (niveles menores a un ng/ml) o concentraciones residuales de estradiol, que provocan pobres manifestaciones del estro y la formación de folículos ováricos persistentes (FOP), que son aquellos que continúan creciendo sin lograr ovular, lo cual provoca estros no fértiles. Por lo que si estos factores se presentan, no se podrá determinar el momento preciso para la inseminación artificial a tiempo fijo (IATF), ya que en el tiempo para realizar la inseminación artificial no habrá un óvulo a quien fecundar y la consecuente baja fertilidad. Esta técnica de inseminación no requiere la detección de celos, sino que se insemina a un número de horas fijado en el mismo protocolo hormonal de sincronización.
El objetivo de este trabajo fue determinar el comportamiento reproductivo posparto de vacas criollas de rodeo con amamantamiento restringido, la respuesta a sincronización del estro con protocolos hormonales, niveles séricos de progesterona y estradiol, la actividad ovárica y tasa de concepción, utilizando inseminación artificial a tiempo fijo.
Materiales y métodos
Área de estudio
El estudio se realizó a finales del verano y principios de otoño de 2008, en el rancho Teseachi en el estado de Chihuahua, México, ubicado en las coordenadas geográficas 28º 48’ de latitud norte y 107º 25’ de longitud oeste. La altitud varía entre 1950 y 2400 msnm, y la precipitación media annual es de 498 mm (Álvarez 1992). Se encuentra en Sierra Madre Occidental, dentro de la subprovincia Sierras y Llanuras Tarahumaras. El clima es un Bs1kw, con temperatura media anual entre 12 y 18 ºC con lluvias en verano y precipitación invernal menor de 5%. El tipo de vegetación predominante es el bosque de encino-pino y bosque de encino (INEGI 2002).
Animales y manejo
Durante el inicio del trabajo se utilizaron 27 vacas criollas de rodeo (C) multíparas, las cuales se trasladaron de su hábitat natural a 55 ha establecidas con un 60% de pasto navajita (Bouteloua gracilis y Bouteloua curtipendula). Las vacas se sometieron por primera vez a amamantamiento restringido (AR) dos veces al día, media hora en la mañana y media hora en la tarde (6:00 y 18:00 h) cuando se encontraban en los 76 días promedio posparto. Se realizó detección de estros durante el amamantamiento más 30 minutos de observación diaria por 22 días consecutivos.
Se observó la conducta social y el comportamiento reproductivo en tiempo y forma durante 45 días en el corral de manejo. A 22 vacas en las cuales se confirmó que iniciaron su actividad reproductiva posparto por medio de la detección de un cuerpo lúteo (CL), se les tomaron imágenes ultrasonográficas durante el AR cada tercer día con un transductor de 7,5 Mhz. A la detección del CL se tomaron muestras de sangre a partir de ese día y durante tres días más, para determinar los niveles séricos de progesterona (NSP).
Aplicación de tratamientos y muestreo
Se asignaron aleatoriamente a 22 vacas en dos tratamientos hormonales para sincronizar su estro, formando dos grupos de 11 animales cada uno. En el tratamiento 1 (T1), las vacas recibieron un dispositivo intravaginal (CIDR) con 1,9 g de progesterona, y una inyección intramuscular con 1 mg de β-estradiol más 50 mg de progesterona. Se retiró el dispositivo intravaginal a los siete días aplicando una inyección IM de Lutalyse con 30 mg de PGF2α. A las 24 horas se aplicó 1 mg de β-estradiol y se inseminaron a las 54 h posteriores al retiro del CIDR. El tratamiento dos (T2) fue similar al T1 pero la inyección IM al momento de aplicar el CIDR no contenía progesterona, sólo 1 mg de β-estradiol (Figura 1).
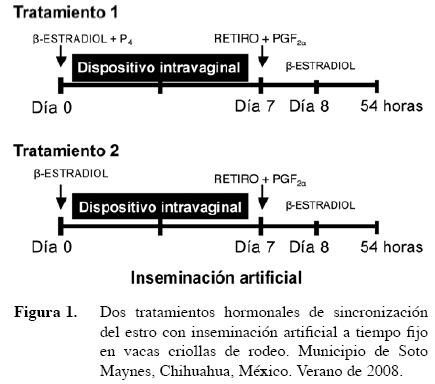
Para evaluar la dinámica folicular durante el tratamiento se tomaron imágenes desde el día cero del tratamiento hasta el día 10 postratamiento. Se grabaron las imágenes con el programa AUSKey para Windows 95. La dinámica folicular fue estudiada agrupando los folículos en cuatro rangos: de 2 a 3 mm, de 4 a 6 mm, de 7 a 10 mm, y > 10 mm de acuerdo con la metodología propuesta por Pierson y Ginther (1984).
Para evitar los problemas de la detección del celo, se han desarrollado protocolos de sincronización de la ovulación; que permiten inseminar animales en un período de tiempo preestablecido, para la aplicación de dicha técnica es necesario sincronizar la vida del cuerpo lúteo, sincronizar el surgimiento de una oleada folicular y la ovulación, así como considerar el tiempo de viabilidad tanto de los espermatozoides como del ovocito una vez realizada la inseminación artificial. Por lo que el tiempo óptimo con el protocolo hormonal utilizado fue establecido a las 54 h posteriores al retiro del CIDR.
El diagnóstico de preñez se determinó por medio de ultrasonografía transrectal a los 45 días posteriores al periodo de tiempo preestablecido para la inseminación artificial a tiempo fijo.
Se tomaron muestras de sangre cada tercer día por medio de punción de la vena coccígea a partir del día cero del tratamiento y diariamente desde el día siete hasta el día diez postratamiento, para comparar los niveles séricos de progesterona y estradiol entre tratamientos. Todas las muestras se transportaron en hielo inmediatamente después de obtenido y se mantuvieron por 24 h a 4 °C hasta su centrifugación. El suero se decantó y se mantuvo a una temperatura de -20 ºC durante dos semanas, hasta que se determinaron las concentraciones de E2 y P4, mediante un estuche comercial en fase sólida (DPC, Los Angeles Ca.). El coeficiente de variación dentro y entre ensayos fue de 3,9% y 15% para P4, y de 5,9% y 5,2% para E2, respectivamente. La medición de los perfiles hormonales se realizó en el laboratorio de radioinmunoanálisis (Schneider y Hallford 1996) de la Universidad Estatal de Nuevo México, localizado en Las Cruces, Nuevo México, EUA.
Variables medidas y análisis estadístico
Se evaluó la condición corporal cada 14 días. El sistema de evaluación fue el descrito por Richards et al. (1989), que especifica valores que van de uno (vaca emaciada) al nueve (vaca obesa). Para determinar si la condición corporal influía sobre el inicio de la actividad ovárica, se analizaron por el método de χ2 la presentación de ovulación y estros durante los 22 días con toma de imágenes ultrasonográficas con PROC FREQ (SAS 2002).
La distribución de horas al estro (HE) posterior a la aplicación de PGF2α se analizó por el método de χ2 por el procedimiento PROC FREQ (SAS 2002).
Para determinar los cambios en la dinámica folicular, se realizó un ANDEVA con el tratamiento y días de la sincronización (DS) como efectos fijos y su interacción T*DS y la condición corporal como covariable. Las variables dependientes fueron el número de folículos por categoría: 2 a 3 mm, 4 a 6 mm, 7 a 10 mm y >10 mm, analizándose por el procedimiento PROC MIXED (SAS 2002). Para determinar diferencias en los niveles séricos de P4 y E2 durante los tratamientos se realizó un ANDEVA, con el T como efecto fijo y su interacción T*DS, por el procedimiento GLM (SAS 2002).
Resultados y discusión
Se observó una relación de dominancia en el comportamiento animal entre las vacas criollas de rodeo ya que con una sola manifestación o actitud de desafío de las vacas con más rango jerárquico sobre las de menor rango, era suficiente para que esta última emprendiera la huida o cediera su espacio. Esta dominancia social no fue sólo marcada por la edad sino también por el tipo de cornamenta, ya que tanto las vacas adultas como jóvenes con alguna deformación o mala disposición de uno o los dos cuernos que les impidiera utilizarlos como una arma efectiva para amedrentar o golpear a las otras, principalmente por la lucha de un espacio o territorio que les proporcionara seguridad a ellas y a su cría durante el amamantamiento.
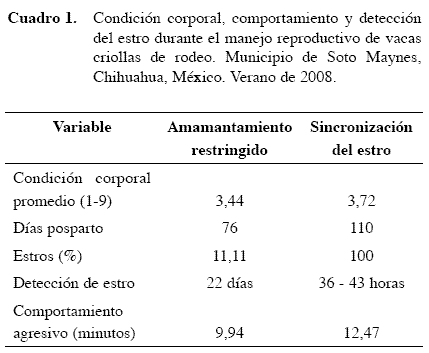
El promedio de comportamiento agresivo observado fue a los 9,94 min de haberse iniciado el manejo de amamantamiento y/o cuando el becerro terminaba de mamar. Este comportamiento agresivo fue retrasándose hasta los 12,47 min posteriores al inicio del manejo de amamantamiento, lo cual se interpretó como un acondicionamiento periódico de las vacas criollas a esta actividad. También se observó durante este período de tiempo que si las vacas permanecían amamantando a sus crías, les daba protección contra las vacas más dominantes del hato, pues si sus crías se alejaban de ellas el riesgo de sufrir agresión aumentaba notablemente. El comportamiento agresivo que resultó en encuentros físicos fue del 35%, todos fueron golpes o envestidas por los costados o por la parte posterior de las vacas y ninguno de frente a frente.
Se observó un comportamiento frecuente de monta en las crías de ambos sexos donde incluso algunas hicieron el intento de montar a sus madres. La literatura reporta ésto en diferentes especies como ovinos, caprinos y bovinos interpretándose como que las crías buscan atraer la atención de sus madres (Reinhardt et al. 1986).
La detección del primer cuerpo lúteo funcional posparto en vacas criollas de rodeo con amamantamiento restringido, fue a los 11,66 días y sólo el 11,11% de las vacas criollas mostraron estro durante este manejo (Cuadros 1 y 2).
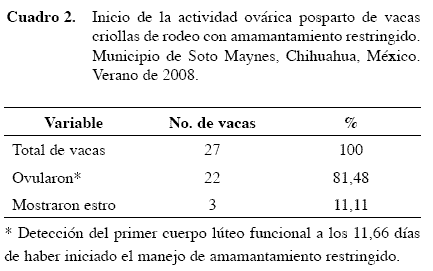
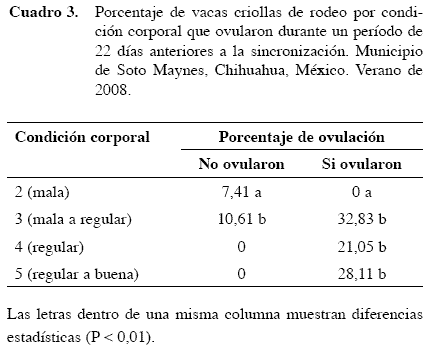
El mayor porcentaje de vacas criollas que mostraron estro (P<0,05; Cuadro 3) fue para las que tenían una condición corporal de tres (condición corporal mala a regular), pero no todas ovularon a diferencia de las vacas criollas con condición corporal cuatro y cinco (condición corporal de regular a buena), las cuales en su totalidad ovularon (Cuadro 4).
El diámetro de folículos ováricos que se detectaron por medio de ultrasonografía durante el inicio de la actividad ovárica posparto fue de 13 mm para el diámetro mayor del folículo dominante (Cuadro 5). El diámetro y área de los cuerpos lúteos al reinicio de la actividad ovárica fueron de 26,0 ± 0,43 mm y 4,72 ± 1.40 cm2 respectivamente.

La concentración media de progesterona durante la detección del primer cuerpo lúteo posparto y los siguientes tres días de muestreo de las 22 vacas que iniciaron su actividad ovárica fue de 8,3±1,66 ng/ml y éstos tuvieron una vida media de tres, 12 y 22 días los cuales correspondieron a ciclos cortos y de una a tres ondas foliculares. Se observó por medio de ultrasonografia que todas las vacas de los dos tratamientos llegaron con un cuerpo lúteo al día de la inserción del CIDR y posteriormente fue confirmado con los resultados de los niveles séricos de progesterona por medio de RIA. El 100% de las vacas criollas presentó estro a las 48 h posteriores a la aplicación de PGF2α y/o del retiro del CIDR.
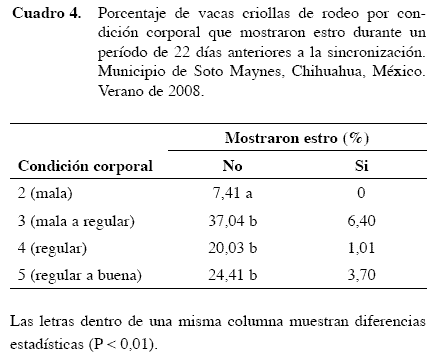
Durante la manifestación del estro sincronizado no se observó conducta agresiva a diferencia del periodo de reinicio de la actividad ovárica el cual se caracterizó por gran manifestación de conducta agresiva antes de mostrar signos de estro. En vez de ésto el comportamiento fue hacer grupos sexualmente activos principalmente de tres hembras por grupo en donde dos de ellas tenían un papel muy activo y la tercera un comportamiento menos pronunciado, pero todas montaron y aceptaron la monta, mostrando preferencia por una de ellas en particular ya que una de las vacas fue montada por todas. A diferencia del periodo de inicio de actividad ovárica se formaron grupos sexualmente activos que se mantenían en contacto estrecho y caminaban alrededor de todo el corral poniendo el mentón sobre la grupa de la otra. No se observaron actividades como lamer u olfatear la vulva así como tampoco topeteo; más bien, el comportamiento fue de un aumento de actividad que resultaba en la monta. No se encontraron diferencias (P>0,05) en el diámetro de folículos preovulatorios por T (T1= 10,15 ± 0,3499 vs T2= 10,62 ± 0,3192). Se observó una interacción T*DS para las concentraciones de progesterona (P=0,06). La concentración de progesterona en el día 0 fue mayor (P<0,05) para T1 comparado con T2 (7,15±0,91 vs 4,82±1,3 ng/ml; Figura 2). Las menores concentraciones de progesterona se registraron durante el estro 0,66±0,74 y 0,70±0,74 ng/ml para T1 y T2, respectivamente.
Se observó también una interacción (P=0,06) T*DS en las concentraciones séricas de estradiol (Figura 2). La concentración de estradiol fue mayor para T1 que para T2 día 0 (P = 0,06; 15,0±2,75 vs 10.0±2,75 pg/ml) y el día nueve (P=0,06; 30,5±17,85 y 26,72±17,85 pg/ml para T1 y T2, respectivamente).
Igual que en vacas de la raza Sahiwal (Mondal y Prakash 2003) y búfalas (Kaur y Arora 1984) las menores concentraciones de progesterona fueron encontradas en las vacas criollas durante el estro, mientras que las concentraciones anteriores al estro fueron similares a las de un cuerpo lúteo de vida media normal.
Los niveles séricos de progesterona y estradiol obtenidos en las vacas criollas indican que la ovulación debió ocurrir el día 10 o 48 h posteriores a la aplicación de 1 mg de β-estradiol en el octavo dνa del protocolo de sincronización (Figura 2). Si milar a vacas de raza comercial y búfalas, las concentraciones de estradiol fueron mayores durante el día del estro (Hafez y Hafez 1993). El número y diámetro de folículos ováricos por categoría que se detectaron por medio de ultrasonografía durante la actividad ovárica posparto y el diámetro medio de los cuerpos lúteos fueron similares a los reportados por Edmondson et al. (1986) en vacas de razas Holstein, Jersey y Ayrshire.
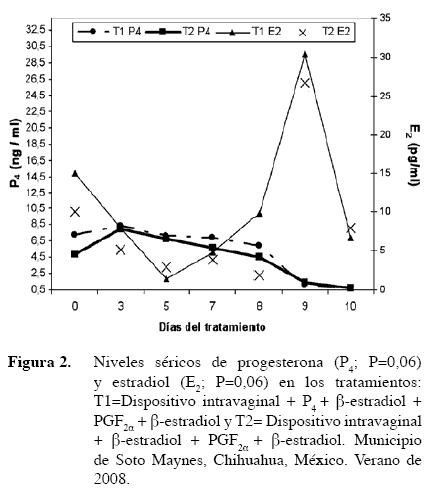
No se detectaron diferencias en el porcentaje de animales que presentaron estros por tratamiento (P>0,05). La distribución de estros mostró una tendencia de agruparse entre las 36 a 43 h posteriores a la aplicación de PGF2α (Figura 3). Estos resultados coinciden con lo reportado por Martνnez et al. (2002) quienes al comparar los tratamientos CIDR y acetato de melengestrol (MGA) encontraron la misma distribución de estros cuando sincronizaron con CIDR.
La expresión del comportamiento estral observada fue muy marcada y puede ser atribuida a buenos niveles promedio de progesterona obtenidos en ambos tratamientos y a que al momento de su aplicación, todos los animales tenían un cuerpo lúteo funcional. Esto se ha observado en los protocolos con aplicación de PGF2α en los cuales, concentraciones altas de progesterona anteriores a la luteσlisis están asociados con un pronunciado comportamiento estral (Stevenson et al. 1998). No se encontraron diferencias entre tratamientos (P>0,05) en el porcentaje de concepción (9,09% para ambos tratamientos), obtenido por medio de IATF. Quizás esto puede explicarse con lo reportado por Smith y Stevenson (1995) quienes concluyeron que la fertilidad no se incrementa cuando un cuerpo lúteo está presente tal como ocurrió en este trabajo ya que todas las vacas criollas asignadas a los dos tratamientos tenían un cuerpo lúteo funcional al momento de la aplicación del CIDR. Todas las vacas que resultaron preñadas se agruparon en las últimas 43 h de detección de estro; lo que podría sugerir que se dejó pasar más tiempo del recomendado para las vacas que se observaron en estro durante las primeras 36 a 37 h. Es tal vez necesario utilizar la técnica de AM-PM para inseminar a las 12 posteriores a la presentación del estro, lo que podría aumentar el porcentaje de concepción. Sin embargo, no puede descartarse la posibilidad de la presentación de estros anovulatorios. Todas las vacas mostraron estro sincronizado entre 36-43 h posteriores a la aplicación de PGF2α y no existieron diferencias entre tratamientos.
Una efectiva detección de estro durante el ciclo estral natural de las vacas criollas se verá afectada no sólo por las condiciones extensivas en las que son criadas sino también por su marcada jerarquía social, territorialidad y manifestación de monta en horas nocturnas entre las hembras en edad reproductiva sometidas a manejo de detección de estros. Sin embargo, al someterlas a este manejo y bajo un protocolo hormonal, facilitó la manifestación del estro y confirma lo encontrado por Rienhardt y Rienhardt (1982), quienes concluyeron que cuando un hato bajo condiciones asilvestradas se maneja bajo reducida agresión, se puede lograr un buen comportamiento reproductivo. El ganado criollo a pesar de pertenecer a las razas Bos taurus podría compartir algunas características con el ganado Bos indicus, ya que se caracterizan por una mayor rusticidad, con periodos interparto largos, asociados a un manejo tradicional de amamantamiento de su cría y deficiencias nutricionales. En otros trabajos también se ha reportado que como resultado de un mayor estrés en el ganado Bos indicus cuando es sometido a sistemas de manejo intenso se han observado diferencias en el tiempo de presentación de los eventos ováricos tales como la emergencia de las ondas foliculares y el diámetro del folículo preovulatorio (Bo et al. 2000). Una baja expresión del estro y pobres resultados en programas de sincronización de estros están asociados con el inconsistente crecimiento y maduración de los folículos ováricos debido a un balance negativo de energía (Stevenson et al. 1987).
Los perfiles séricos de progesterona y estradiol, junto con el comportamiento estral indican que ambos protocolos hormonales utilizados produjeron condiciones similares a las de un ciclo estral natural. La expresión del comportamiento estral observada en este trabajo fue muy marcada y son atribuidos a los niveles de progesterona obtenidos en ambos tratamientos. La contribución de progesterona para inhibir o estimular los signos de estro, dependen de su acción sobre diferentes sitios hipotalámicos y el subsiguiente cambio de receptores de progestágenos en cualquiera de estas áreas (Takahashi y Lisk 1988), por lo que las concentraciones preliminares de progesterona son determinantes para la presentación de un comportamiento estral adecuado (Caraty et al. 2002). Por otra parte otros trabajos han mostrado que al utilizar protocolos con estradiol éste induce la aparición de una nueva onda folicular por la supresión de FSH (Bo et al. 1995).
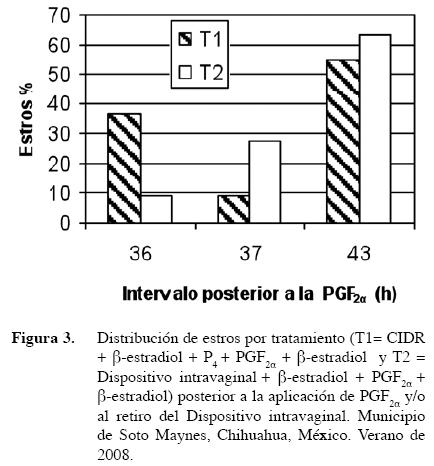
El estradiol y la progesterona aplicados al inicio del protocolo hormonal juntos con el CIDR en vaquillas productoras de carne fueron seguidos por atresia folicular y la sincronización de una nueva onda folicular a los 3,4±0,10 días posteriores a la aplicación (Martínez et al. 2002), lo cual coincidió con los resultados de los protocolos utilizados en el presente estudio que al remover el CIDR en el día siete el folículo dominante se encontraba en los días tres y cuatro de su fase de crecimiento correspondientes a un estro natural (7-10 mm), produciendo el desarrollo de una nueva onda folicular y permitiendo el crecimiento de folículos ováricos pequeños dependientes de FSH (Stock y Fortune 1993), por lo que no hubo formación de FOP debido a concentraciones subluteales de progesterona o niveles residuales de estradiol (Figura 4).
El mayor logro del presente estudio fue sincronizar al 100% de los animales experimentales utilizando dos protocolos hormonales con CIDR más (50 mg de progesterona + 1 mg de estradiol) y PGF2α, o CIDR mαs 1 mg de estradiol sin progesterona y PGF2α. Estos resultados no se habνan logrado en trabajos anteriores con vacas criollas utilizando protocolos hormonales con implantes de norgestomet. Sin embargo, debido a que no se esperaba provocar la manifestación del estro con tanta precisión así como una distribución de estros tan agrupada, se siguió el protocolo tal como se propuso y se inseminó a tiempo fijo lo que posiblemente produjo una reducción en la tasa de concepción (18,8%), ya que sólo las vacas que resultaron preñadas fueron las que se agruparon en las últimas 43 horas de detección de estro, lo que sugiere que se dejó pasar más tiempo del recomendado para las vacas que se observaron en estro durante las primeras 36 a 37 horas, por lo que el bajo porcentaje de concepción con IATF fue causado porque el tiempo de ovulación en las vacas criollas es muy variable al utilizar estos protocolos hormonales y no coincide con el tiempo de viabilidad que necesitan los espermatozoides para realizar la fecundación del ovocito. De acuerdo con estos resultados, se sugie re evaluar la inseminación artificial con base en la técnica AM-PM de manera que las vacas se inseminen aproximadamente a las 12 horas posteriores al estro, lo que podría aumentar el porcentaje de vacas preñadas. El 81,82% restante de las vacas sincronizadas quedaron preñadas con monta natural a los 50 días del empadre con toros criollos de rodeo.
La relación social jerárquica observada durante el manejo de amamantamiento en las vacas criollas de rodeo fue fuertemente marcada, caracterizándose por un comportamiento agresivo, movimientos intimidatorios, golpes y heridas principalmente en el tren posterior.
La actividad ovárica posparto dio inicio durante el manejo de amamantamiento restringido, pero la manifestación de comportamiento agresivo de las vacas criollas de rodeo en este período no permitió una eficiente detección del estro.
Todas las vacas que fueron tratadas con protocolo hormonal mostraron estro sincronizado, pero no existieron diferencias entre las concentraciones séricas de progesterona entre T1 y T2. En el caso de estradiol se observó una mayor concentración en el día del estro para T1 comparado con T2. No se pudo comprobar la presentación de folículos ováricos persistentes (FOP) debido a concentraciones subluteales de progesterona o niveles residuales de estradiol, ni diferencias en el porcentaje de concepción en ninguno de los dos tratamientos por lo que adicionar 50 mg de progesterona al momento de aplicar un CIDR no incrementó la tasa de concepción en vacas criollas de rodeo. Se observó que a mejor condición corporal de vacas criollas de rodeo mejora su eficiencia reproductiva.
Agradecimientos
A la Fundación Produce Chihuahua A.C., al Consejo Nacional de Ciencia y Tecnología y Fondos Mixtos del estado de Chihuahua, quiénes fueron fuente de financiamiento parcial del presente estudio.
Recibido: 23 de mayo, 2009. Aceptado: 17 de mayo, 2010.
Literatura citada
Álvarez, A. 1992. Compendio de datos climatológicos del Estado de Chihuahua. Gobierno del Estado de Chihuahua, México. p. 32. [ Links ]
Bo, GA; Adams GP; Pierson, RA; Mapletof, RJ. 1995. Exogenous control of follicular wave emergence in cattle. Theriogenology 43:31-40. [ Links ]
Bo, GA; Bergfelt, DR; Brogliatti, GM; Pierson, RA; Adams, GP; Mapletoft, RJ. 2000. Local versus systemic effect of exogenous estradiol-17β on follicular dynamics in heifers with progestogen implants. Anim Reprod Sci 59:141-157. [ Links ]
Caraty, A; Delaleum, B; Chesneau, D; Fabre-Nys, C. 2002. Se quential role of E2 and GnRH for the expression of estrous behavior in ewes. Endocrinology 143:139-145. [ Links ]
Edmondson, AJ; Fissore, RA; Pashen, RL; Bondurant, RH. 1986. The use of ultrasonography for the study of the bovine reproductive tract. I. Normal and Pathological Ovarian Structures. Anim. Reprod. Sci. 12:157-165. [ Links ]
Hafez, ESE; Hafez, B. 1993. Reproductive cycles: cattle and buffalo. Reproduction in farm animals. Williams and Wilkins Press. New York, USA. 563 p. [ Links ]
INEGI (Instituto Nacional de Estadística y Geografía). 2002. Síntesis de información geográfica del estado de Chihuahua. Anexo cartográfico. Aguascalientes, Ags. Mex p. 47. [ Links ]
Kaur, H; Arora, SP. 1984. Anual pattern of plasma progesterona in normal cycling buffaloes (Bubalus bubalis) fed two levels of nutrition. Anim. Reprod. Sci. 7:323-332. [ Links ]
Martínez, MF; Kastelic, JP; Adams GP; Mapletof, RJ. 2002. The use of a progesterone-releasing device (CIDR-B) or melengestrol acetate with GnRH, LH, or estradiol benzoate for fixed-time AI in beef heifers. J. Anim. Sci. 80:1746-1751. [ Links ]
Mondal, S; Prakash, BS. 2003. Periferal plasma progesterone concentrations in relation to estrus expression in Sahiwal cows. Ind. J. Physiol. Endocrinol. 47:111-114. [ Links ]
Pierson, RA; Ginther, OJ. 1984. Ultrasonography of the bovine ovary. Therioogenology 21 (3): 495-504. [ Links ]
Ramírez, JA; Torres Si mental, JF; Gómez Castañeda, JA; Flores-Marileñarena, A; Jiménez Castro, JA; Rodríguez Almeida, FA. 1998. Aspectos reproductivos del ganado Criollo (Rodeo). Memoria Segundo Foro de Análisis de los Recursos Genéticos: Ganado Criollo. Programa Nacional de Recursos Genéticos Pecuarios. Chihuahua, Chih. México. p. 23-32. [ Links ]
Reinhardt, V; Reinhardt A. 1982. Mock fighting in cattle. Applied Animal Behaviour Science 81:1-13. [ Links ]
Reinhardt, C; Reinhardt, A; Reinhardt, V. 1986. Social behaviour and reproductive performance in semi-wild Scottish Highland cattle. Applied Animal Behaviour Science 15:125-136. [ Links ]
Richards, MW; Wetteman, RP; Schoenemann, HM. 1989. Nutritional anestrus in beef cows: Body weight change, Body condition, luteinizing hormone in serum and ovarian activity. J. Anim. Sci. 67:1520-1525. [ Links ]
Rhodes, FM; McDougall, S; Burke, CR; Verkerk, GA; Macmillan, KL. 2003. Treatment of cows with an extended postpartum anestrous interval. J. Dairy. Sci. 86:1876-1894. [ Links ]
SAS (Statistical Analysis System). 2002. User’s guide. Version 9.0. SAS Institute Inc., Cary, NC, USA. [ Links ]
Schneider, FA; Hallford, DM. 1996. Use of rapid progesterone radioimmunoassay to predict pregnancy and fetal numbers in ewes. Sheep and Goat Research. J. 12:33- 38. [ Links ]
Smith, MW; Stevenson, JS. 1995. Fate of the dominant follicle, embryonal survival, and pregnancy rates in dairy cattle treated with prostaglandins F2 and progestins in the absence or presence of a functional corpus luteum. J. Anim. Sci. 73:3743-3751. [ Links ]
Stevenson JS; Lucy, MC; Call, EP. 1987. Failure of timed insemination and associated luteal function of PGF2α. Theriogenology 28:937-943. [ Links ]
Stevenson, JS; Lamb, GC ; Kobayashi, Y; Hoffman, DP. 1998. Luteolysis during two stages of estrous cycle: subsequent endocrine profiles associated with raditelemetrically detected estrus in heifers. J. D. Sci. 81:2897-2903. [ Links ]
Stock, AE; Fortune, JE. 1993. Ovarian follicular dominance in cattle: Re lationship between prolonged growth of the ovulatory follicle and endocrine parameters. Endocrinology 132:1108-114. [ Links ]
Takahashi, LK; Lisk, RD. 1988. Dual progesterone action in diencephalon facilitates the induction of sexual receptivity in estrogen-primed golden hamsters. Physiol. Behav. 44:741-747. [ Links ]

