











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Uno de los principales objetivos en los estudios ecológicos es determinar los factores que limitan la distribución y abundancia de las especies de plantas (Godínez-Alvarez & Valiente-Banuet 2004). Los factores limitantes son una combinación de condiciones bióticas y abióticas que operan a diferentes escalas tanto espaciales como temporales, y que definen en gran medida la distribución de las especies (Maldonado & Mondragón-Chapano 2007). La comprensión de estos factores limitantes permite identificar las condiciones necesarias para la supervivencia de las especies a escala de paisaje (Münzbergová & Herben 2005), y en consecuencia determinar los hábitats donde son más efectivas las medidas de manejo. Todo esto puede ser crítico para el mantenimiento de la diversidad (Gowland et al. 2007). La disponibilidad de agua, luz, el rango de temperatura y el tipo de suelo son algunos de los factores que probablemente limiten la distribución de muchas especies (Guo et al. 2011, Song et al. 2013). En general, la preferencia del hábitat es asumida como adaptativa, y si las condiciones son buenas en estos sitios, se produce una selección natural que mantiene tales preferencias (Martin 1998).
La distribución de las plantas epífitas está fuertemente influenciada por las características de su forófito, que provee el sustrato necesario para el establecimiento (Hirata et al. 2009). Entre las características del forófito de mayor influencia está la arquitectura arbórea (Flores- Palacios & Ortíz-Pulido 2005, Otero et al. 2007, Winkler & Hietz 2001), las características físicas y químicas de la corteza, su estabilidad, la disponibilidad de área colonizable y la cobertura vegetal (Callaway et al. 2002, Maldonado & Mondragón-Chapano 2007). Estas características junto a las condiciones microclimáticas y los requerimientos ecofisiológicos de las epífitas puede limitar la distribución y abundancia de las mismas (Maldonado & Mondragón-Chapano 2007, McCormick et al. 2012).
Los factores limitantes no actúan de manera independiente, según Hirata et al. (2009) estos pueden interactuar entre sí, por lo que para entenderlos es necesario un análisis simultáneo de los mismos. Estudios en la última década han examinado la influencia del tamaño del hospedero sobre la diversidad de epífitas, encontrando una fuerte relación entre ambas variables (ver Hirata et al. 2009). En el caso de las orquídeas sus microhábitats preferenciales están más limitados aún, debido a que sus semillas dependen de la disponibilidad de hongos micorrízicos para germinar y establecerse (Flores-Palacios & Ortíz- Pulido 2005).
Las orquídeas constituyen una de las familias más numerosas de la flora de Cuba, con un alto grado de especies endémicas y un 71% de epifitismo (García- González et al. 2016). Una de estas especies endémicas es Encyclia pyriformis (Lindl.) Schltr., característica de las arenas blancas del Occidente de Cuba (Ackerman 2014, Vale et al. 2014). Esta orquídea tiene hojas cortas e inflorescencias con pocas flores (con respecto al resto de las especie del género), labelo obcordado y crestas del labelo que terminan en cuernos. Hasta el momento se considera una epífita específica del tronco de la palma Acoelorrhaphe wrightii (Griseb. & H.Wendl.) H.Wendl. ex Becc. (Vale et al. 2014). Su escasa y restringida distribución dificultan los estudios de autoecología, lo cual provoca que se desconozca el efecto que puedan tener ciertas variables ambientales sobre el desarrollo de los individuos de E. pyriformis. Esta información podría ser clave en el desarrollo de planes para su manejo exitoso, dado que es una especie que vive cerca de costas, la cual puede verse seriamente afectada por los efectos del cambio climático. Teniendo en cuenta estos aspectos el presente estudio tiene como objetivos caracterizar el uso del microhábitat de E. pyriformis y analizar la relación entre las variables morfológicas y el microhábitat usado en la Reserva Ecológica Los Pretiles, Cuba.
Materiales y métodos
Área de estudio.- El estudio se realizó en junio de 2015 en la Reserva Ecológica Los Pretiles, ubicada en la región noroeste del municipio Mantua, Pinar del Río (Figura 1). El área protegida tiene una extensión de 37,100 ha, de las cuales 451.8 ha son de zona terrestre (Márquez et al. 2015). La temperatura media anual se mantiene alrededor de los 25.7°C y las precipitaciones no superan los 1,100 mm anuales. El bosque de pino es una de las principales formaciones vegetales del área y está constituido por un bosque abierto cuyos árboles (Pinus tropicalis Morelet) ocupan entre el 20-40% de la cobertura. El estrato superior está compuesto, principalmente por Acoelorrhaphe wrightii, Tabebuia lepidophylla (A.Rich.) Greenm., Byrsonima pinetorum C.Wright ex Griseb., Lyonia ekmanii Urb., Lyonia lucida (Lam.) K.Koch y Morella cerifera (L.) Small.
Muestreo y análisis de datos.- El muestreo se realizó en el pinar Los Pretiles, perteneciente al sector 1 “La Isla”, donde se marcaron 39 parcelas de 25 m2, posicionadas en 4 recorridos lineales perpendiculares a la línea de costa. Para ubicar las parcelas se siguió un muestreo sistemático, con una distancia de 15 m entre cada parcela, con inicio en la línea de costa y con una separación de 50 m entre cada recorrido lineal. Debido al crecimiento cespitoso de esta y otras especies de Encyclia (Vale et al. 2014) delimitar un individuo genéticamente diferente de otro puede ser un tarea inviable en el campo. Por esta razón se siguió el criterio de Sanford (1968), quien define a un individuo como un grupo de rizomas y hojas pertenecientes a una misma especie, que forman una unidad claramente delimitada (Figura 2). Este mismo criterio fue utilizado por Zotz (2007) para varias especies de orquídeas y otras epífitas. Dentro de las parcelas se identificaron a todos los individuos de E. pyriformis y para cada uno se midieron seis variables externas (o ambientales): diámetro del tronco ocupado, cobertura vegetal, distancia a la costa, altura sobre el suelo, altura del forófito y orientación. El diámetro del tronco fue medido con un pie de rey en la misma zona donde estaba la orquídea. La altura del forófito se midió con una cinta métrica, y la cobertura vegetal mediante una fotografía tomada encima de cada individuo de E. pyriformis. Las fotos fueron procesadas en el programa GapLight Analysis, que permite calcular el porcentaje de cobertura de cada imagen. Para calcular la distancia a la costa se tomaron las coordenadas geográficas de cada parcela, las cuales se montaron sobre el mapa de línea de costa de Cuba, en el sistema de información geográfica ArcGis 10.1. La orientación de cada individuo en el forófito fue determinada con una brújula. Además, sobre cada individuo se midió: la altura de la orquídea, el diámetro del pseudobulbo (correspondiente a la hoja mayor), y el largo y ancho de la hoja mayor. Algunas mediciones fueron realizadas con una cinta métrica (error 0.1 cm) y otras con un pie de rey (error 0.01 cm). En las 39 parcelas fueron identificados y muestreados 138 individuos de E. pyriformis.

Tabla 1 Estadísticos descriptivos de las variables ambientales de los individuos de Encyclia pyriformis (n=138) en la Reserva Ecológica Los Pretiles, Cuba.
| Variable | Media | Dev. Std. | LC inf (95%) | LC sup (95%) | Min | Max |
|---|---|---|---|---|---|---|
| Diámetro del tronco (cm) | 9.42 | 1.84 | 9.11 | 9.73 | 6.2 | 13.9 |
| Cobertura vegetal (%) | 66.58 | 20.80 | 63.07 | 70.08 | 0 | 94.43 |
| Distancia a la costa (m) | 173.1 | 114.8 | 153.78 | 192.43 | 0 | 373 |
| Altura sobre el suelo (m) | 0.84 | 0.52 | 0.75 | 0.93 | 0 | 2.30 |
| Altura del forófito (m) | 2.37 | 0.84 | 2.22 | 2.51 | 0.69 | 5 |
Se calcularon los estadísticos descriptivos para las variables ambientales (media, desviación estándar de la media, límites de confianza, mínimo y máximo). Se realizó una prueba de Mantel (10,000 iteraciones), para determinar si existía relación entre las variables morfológicas y las variables ambientales (sin considerar la orientación por ser una variable nominal). Además, se relizaron pruebas de Mantel (10,000 iteraciones) entre las variables morfológicas y las variables ambientales por separado.
Resultados
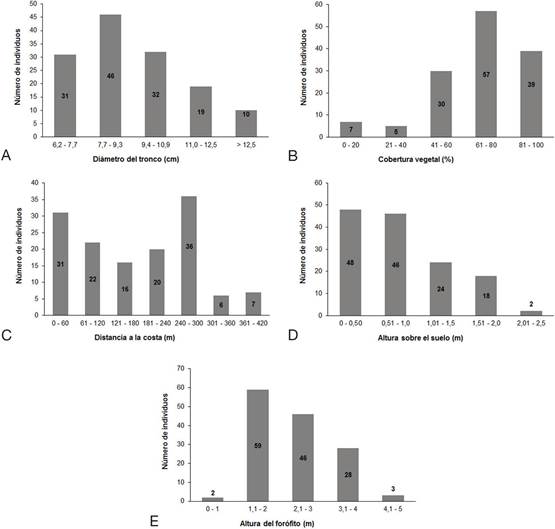
Los individuos de E. pyriformis en la Reserva Ecológica Los Pretiles siempre fueron encontrados sobre A. wrightii y se encontraban como promedio en troncos de 9.42±1.84 cm de diámetro, y con un valor máximo de 13.9 cm (Tabla 1). La mayor concentración de individuos de la orquídea estuvo entre los 7.3 y 9.3 cm de diámetro, con un total de 62. La segunda mayor concentración de individuos de E. pyriformis se encontró entre los 10.5 y los 12.5 cm de diámetro del tronco ocupado (Figura 3A). Seis individuos de E. pyriformis presentaron valores de cobertura de 0%, por lo que representan orquídeas que estaban complemente expuestas a la luz solar. El valor promedio de esta variable fue de 66.58±20.8%, siendo 94.43% el valor máximo de cobertura registrado para la especie (Tabla 1). El 91% de los individuos se encontraron en valores de cobertura entre el 40 y el 95% (Figura 3B). Por debajo de este rango solo exitían unos pocos individuos.
Encyclia pyriformis puede crecer desde la línea de costa hasta 373 m costa adentro (Tabla 1). La mayoría de los individuos fueron reportados en los intervalos de 0-60 m y 241-300 m (Figura 3C). Como promedio los individuos de E. pyriformis se encontraron a 0.84±0.52 m sobre el suelo, reportándose un individuo que estaba a nivel de suelo y otro a 2.3 m sobre el suelo (Tabla 1), lo que representan los valores extremos de la variable. Cerca del 8% de los individuos de E. pyriformis se encontraron en alturas inferiores a 1 m (Figura 3D), y solo 2 individuos fueron registrados en alturas superiores a los 2 m sobre el suelo.
El valor promedio de la altura del forófito (A. wrightii) fue 2.37±0.84 m (Tabla 1). El 96% de los individuos de E. pyriformis se encontraron en forófitos que presentaron alturas entre 1 y 4 m (Figura 3E). Al analizar las variables altura del forófito y altura sobre el suelo de la orquídea, se observa que el valor promedio de la primera fue 2.37 m y el de la segunda de 0.84 m. Esto demuestra que E. pyriformis solo aprovecha un 35 % del área disponible y siempre hacia el primer tercio del forófito. El valor máximo de altura sobre el suelo donde se encontraron individuos de E. pyriformis fue de 2.30 m mientras que la altura máxima de su forófito fue de 5 m.
La orientación cardinal encontrada con mayor frecuencia en la población de E. pyriformis fue la norte con un total de 32 individuos (Figura 4). La segunda orientación que más se presentó en la población fue la helicoidal, en la cual los individuos le dan la vuelta a todo el tronco en el cual se desarrollaban. Las exposiciones menos frecuentes fueron las sureste con solo seis individuos, la oeste con ocho y noroeste con nueve individuos (Figura 4). Los estadísticos descriptivos de las variables morfológicas de E. pyriformis se muestran en la Tabla 2.
No se encontró correlación estadísticamente significativa entre las variables ambientales y las variables morfológicas de E. pyriformis (Figura 5). El valor de la prueba de Mantel para esta correlación múltiple fue de 0.047, con un valor de probabilidad del 8%. Al correlacionar las variables ambientales con las morfológicas de manera independiente (Tabla 3), se puede observar que solo existió correlación estadísticamente significactiva para las variables diámetro del tronco y distancia a la costa. Al parecer estas son las dos variables que presentaron algún tipo de influencia sobre la morfología vegetal de E. pyriformis.
Discusión
A una escala local como la del presente estudio, la distribución de las especies es, en teoría, limitada por la disponibilidad de micrositios apropiados y por la capacidad de dispersión de cada especie (Frei et al. 2012). En la mayoría de orquídeas la dispersión es mediada por el viento (González et al. 2007), lo que hace más azaroso el movimiento de las diásporas. La disponibilidad de micrositios como principal factor limitante ha sido reportado para otras especies de orquídeas como Oncidium poikilostalix (Kraenzl.) M.W.Chase & N.H.Williams (García-González et al. 2011), Ionopsis utricularioides (Sw.) Lindl. (García- González & Riverón-Giró 2014) y Telipogon helleri (L.O.Williams) N.H.Williams & Dressler (García- González & Damon 2013). Según Frei et al. (2012) las principales causas de disponibilidad de micrositios idóneos son factores ambientales como el clima, el suelo y la sucesión vegetal. Además de factores bióticos como la competencia y otras interacciones como es la presencia de hongos micorrízicos apropiados, sin los cuales la germinación de las semillas de orquídeas es imposible (Winkler et al. 2009).
Tabla 2 Estadísticos descriptivos de las variables morfológicas de los individuos de Encyclia pyriformis (n=138) en la Reserva Ecológica Los Pretiles, Cuba.
| Variable (cm) | Mean | Std. Dev. | LC inf (95%) | LC sup (95%) | Min | Max |
|---|---|---|---|---|---|---|
| Altura del individuo (cm) | 24.90 | 11.51 | 22.96 | 26.84 | 4 | 61 |
| Diámetro del pseudobulbo (cm) | 1.87 | 0.54 | 1.77 | 1.96 | 0.4 | 3.4 |
| Largo de la hoja mayor (cm) | 13.62 | 5.56 | 12.68 | 14.56 | 2 | 30 |
| Ancho de la hoja mayor (cm) | 2.06 | 0.58 | 1.96 | 2.15 | 0.3 | 3.8 |
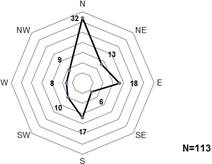
Figura 4 Diagrama de las orientaciones cardinales de Encyclia pyriformis en la Reserva Ecológica Los Pretiles, Cuba.
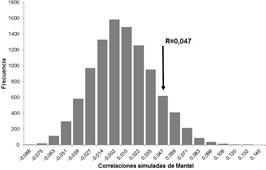
Figura 5 Histograma de frecuencia donde se muestran las correlaciones simuladas de Mantel (10,000 iteraciones) entre las variables ambientales (diámetro del tronco, altura sobre el suelo, cobertura vegetal, altura del forófito, distancia a la costa) y las variables morfológicas (altura de la orquídea, diámetro del pseudobulbo, largo y ancho de la hoja mayor) de Encyclia pyriformis. R de Mantel=0,047.
Otros factores no identificados relacionados con las condiciones del microhábitat pueden limitar la ditribución y la abundancia de las orquídeas (McCormick et al. 2012, Otero et al. 2013). Los valores mínimos y máximos de las variables analizadas representan las condiciones extremas en que se desarrollan los individuos de E. pyriformis muestreados en la Reserva Ecológica Los Pretiles. Los resultados obtenidos permiten asegurar que los individuos de E. pyriformis no usan de manera homogénea el área del forófito, de hecho usan menos del 40% del área disponible y siempre hacia la porción inferior del forófito. Esto podría estar influenciado por la alta intensidad lumínica que inside sobre las zonas más altas del forófito y por el hecho de que la humedad relativa en las zonas inferiores del forófito es mayor. Esta última causa cobra gran importancia si se tiene en cuenta que las orquídeas epífitas son organismos que requieren de altos valores de humedad relativa (Maldonado & Mondragón-Chapano 2007). Benavides et al. (2005) encontraron que la mayor concentración de epífitas de la amazonía estaba en la base de los troncos.
Tabla 3 Valores de las correlaciones simuladas de Mantel entre cada una de las variables ambientales y las variables morfológicas (**correlación estadísticamente significativa).
| Variable | R Mantel | Percentil inferior (2.50 %) | Percentil superior (97.50 %) |
|---|---|---|---|
| Diámetro del tronco (cm) | 0.140** | -0.066 | 0.076 |
| Cobertura vegetal (%) | 0.011 | -0.077 | 0.109 |
| Distancia a la costa (m) | 0.0559** | -0.047 | 0.0553 |
| Altura sobre el suelo (m) | 0.004 | -0.068 | 0.083 |
| Altura del forófito (m) | -0.028 | -0.070 | 0.087 |
| Todas las variables | 0.047 | -0.057 | 0.064 |
La mayoría de los individuos de E. pyriformis presentaron valores de cobertura superiores al 50%, esto apoya lo planteado por Arévalo & Betancour (2006) y por Guo et al. (2011) de que la intensidad lumínica es uno de los factores que más influye en la distribución de las orquídeas epífitas. También puede ser explicado por el hecho de que las especies epífitas pueden experimentar daños en el fotosistema ante altos valores de intensidad lumínica (Callaway et al. 2002). Según Flores-Palacios y Ortiz-Pulido (2005) uno de los principales factores de mortalidad de las orquídeas es la desecación; esta puede ser la causa del hecho de que casi todos los individuos de E. pyriformis se encontraran en las porciones inferiores de los troncos de A. wrightii y en valores de cobertura vegetal superiores al 50%. La baja variabilidad que presentó el diámetro del tronco del forófito sugiere que esta variable podría ser de gran importancia en la selección de micrositios para E. pyriformis. Esto concuerda con Arévalo & Betancour (2006), quienes plantean que el diámetro del tronco, su inclinación y posición, presencia de ramificaciones y su rugosidad son factores que influyen en la distribución de las plantas epífitas.
El diámetro del tronco, la cobertura vegetal y la altura del forófito son variables que caracterizan a la especie hospedera. Según Hirata et al. (2009) una combinación de rasgos del hospedero determina la presencia de epífitas. La especie que actúa como forófito provee el sustrato para las espífitas, por lo que el establecimiento de estas se ve afectado por rasgos del hospedero que incluye el área del tronco disponible para el establecimiento de la epífita, las características físicas y químicas de la corteza, la arquitectura del forófito y la cobertura vegetal (Hirata et al. 2009). En el caso de las orquídeas con una elevada afinidad por uno o pocos forófitos, la preferencia de microhábitat depende en gran medida de la preferencia de microhábitat de su(s) forófito(s) (Otero et al. 2007). En el caso de E. pyriformis, que es altamente específica para A. wrightii (Ackerman 2014, Vale et al. 2014), cabría esperar que su distribución se vea altamente influenciada por los requerimientos de hábitat de su forófito.
La ausencia de correlación entre las variables ambientales y morfológicas parece indicar que la combinación de variables ambientales analizadas no tienen un claro efecto sobre la morfología de E. pyriformis. Sin embargo, la prueba de Mantel analiza solamente correlación lineal entre las variables, puede ser el caso de que la correlación que exista entre estas variables no siga esta premisa. A pesar de no existir una correlación evidente, el valor de la R de Mantel (valor de correlación obtenido) cae cerca de los extremos de la distribución de frecuencias. Lo cual puede ser tomado como una evidencia de que puede existir algún tipo de influencia no detectada por la prueba estadística. El análisis de cada variable por separado mostró un resultado similar, pues tres de las variables analizadas no presentaron correlación significativa con las características morfológicas. Una limitación del presente estudio es el “conflicto de estados de vida” o de “cambio ontogenético del nicho”, que plantea que la calidad del micrositio puede cambiar entre la semilla, la plántula y el adulto. Un micrositio puede ser favorable para que el estado de semilla se establezca, pero se vuelve menos favorable para otros estados (Scott & Morgan 2012). Aún así, entender los factores ambientales que limitan y afectan la distribución de las especies es crítico para el mantenimiento de la diversidad (Gowland et al. 2007). Esto cobra gran relevancia cuando se trabaja con orquídeas epífitas, que son especialmente sensibles al cambio climático (Seaton et al. 2013). Desde el punto de vista conservacionista, investigar los factores que limitan el potencial de colonización de esta orquídea permitiría predicciones certeras ante cambios futuros en la comunidad y el ecosistema lo cual puede influir en las estrategias de manejo de la especie.

