










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkMedicina Legal de Costa Rica
On-line version ISSN 2215-5287Print version ISSN 1409-0015
Med. leg. Costa Rica vol.30 n.1 Heredia Mar. 2013
Original
Evaluación práctica de las alteraciones postmortem debidas a la actividad de los artrópodos
Alejandro González Medina+*, Lucas González Herrera*+, José de la Higuera Hidalgo+, Gilberto Jiménez Ríos+
Resumen
Palabras clave
Entomología Forense; tafonomía; diptera; coleoptera; lepidoptera; modificación postmortem.
Abstract
After death of a living organism, the activity of insects is the main mechanism by which decomposing organic matter returns to the nutritional chain. However, it is necessary to consider that this activity can imply modifications of great taphonomic and forensic interest. If its magnitude is not known well, it can induce to error in the later analysis of the forensic pathologist. In this article, we give an organized classification of the different taphonomic phenomena induced by insects (addition, elimination and modification of the corpse) and enunciate the differential features that allow to separate them from similar pathological processes, focusing our attention on the orders Coleoptera, Diptera and Lepidoptera as the main responsibles of these modifications.
Keywords
Forensic entomology; taphonomy; diptera; coleoptera; lepidoptera; postmortem modification.
Establecer este punto reviste una importancia capital ya que, obviando cuestiones legales y patrimoniales, permite que se daten otros eventos relacionados con la muerte, tales como la vitalidad de las lesiones, esclarecer una posible agonía...
Debido a la gran variabilidad observada en el tiempo de aparición de una determinada fase de descomposición cadavérica debida a factores externos e internos, otros métodos se hicieron rápidamente necesarios. Se han desarrollado pruebas a partir de muy diversas metodologías, pero las más fiables a partir de las 72 horas del fallecimiento son las aportadas por la Entomología Forense (2). El empleo de los artrópodos y de los conocimientos de su biología a la hora de resolver cuestiones médico-legales es una rama de la Entomología que cada vez presenta mayor aplicabilidad en los tribunales de justicia.
Estos fenómenos pueden distorsionar los resultados relativos a la data de la muerte (Entomología Forense) o bien los de la causa o circunstancias que rodearon la misma (Patología Forense).
Alteraciones por adición
Consideraremos en este grupo de alteraciones aquellas en las que, al material que podríamos encontrar en el lugar del levantamiento, se añaden estructuras materiales que, en ausencia de sus fuerzas productoras, no aparecerían.
Contaminación por larvas, pupas o adultos:
No es poco frecuente que, por motivos criminales o de vida social marginal, se encuentren cadáveres humanos asociados a los detritos orgánicos que periódicamente desechamos en la vida cotidiana.
Algunos, como los enfermos de Síndrome de Diógenes, los acumulan en el interior de sus viviendas. La materia orgánica contenida en estos subproductos constituye un sustrato alimenticio de gran valor para especies que también pueden encontrarse en escenarios forenses (Muscidae, Calliphoridae, Sarcophagidae...) o que son exclusivas de estos medios (larvas de Drosophilidae, de Scatophagidae...), aunque muchas veces la discriminación no es tan clara.
Si un cuerpo sin vida es abandonado en las cercanías de un lugar rico en este tipo de sustrato, es necesario tener en cuenta la posibilidad de una contaminación cruzada con transferencia deadultos o de larvas desde los desechos orgánicos hasta el cadáver(6). Conocer los antecedentes del caso y realizar pruebas de contenido digestivo en busca de DNA o antígenos humanos parecen ser las mejores soluciones para abordar esta "adición" de material foráneo(7)(8).
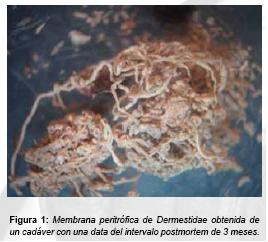
Membrana peritrófica de Coleópteros
La aparición y persistencia de estos elementos depende en gran medida de las condiciones ambientales en las que se encuentren los restos humanos. Es necesario que transcurra un cierto tiempo (que puede ir desde 1 mes hasta una década) (5), que los restos se conserven en un medio en el que no haya demasiado movimiento ni tracciones que puedan dañar las membranas y que la zona corporal donde se generó haya sufrido una desecación rápida en ausencia de actividad larvaria de dípteros ya que éstos podrían añadir humedad al medio retrasando la aparición de las membranas. Frecuentemente, las regiones corporales cubiertas con ropa de cierta consistencia suelen generar algunas de estas condiciones ambientales. Los Coleoptera generadores de membrana peritrófica más frecuentes en casos forenses son los pertenecientes a la familia Dermestidae(10).
Manchas de sangre debidas a la actividad de moscas adultas
Un criterio que no debe usarse es la reacción positiva frente a los kits comerciales de identificación presuntiva de sangre, ya que dan positivo también en las regurgitaciones y defecaciones de los Diptera(14)(15).
Alteraciones por eliminación
Las alteraciones por eliminación son aquellas en las que parte del material procedente de los restos humanos es eliminado o deteriorado con pérdida de materia.
Cámaras de pupación de Dermestidae y Tineidae (Lepidoptera)
Pueden dar lugar a confusión con patologías reumáticas, pero presentan una serie de características únicas: las de mayor tamaño presentan una localización casi exclusiva en los márgenes interarticulares, las de menor tamaño se encuentran en zonas anti-declive y en los tubérculos óseos cercanos a la articulación pero no integrantes de ella, nunca se encuentran aisladas ni presentan gran variación de diámetro dentro del mismo hueso, afectan únicamente a la sustancia compacta de las epífisis y presentan lados claramente verticales con sección transversal en forma de U(18)(19).
Dentro de las cámaras hay tres grandes tipos: los "hoyos hemisféricos", que sólo se producen cuando el derméstido horada directamente el hueso; los "hoyos planos", producidos cuando la excavación de la cámara comienza sobre un sustrato de carne seca; y las "rosetas", que constituyen la fase inicial de la formación de un "hoyo plano"(20)(21)(figura 3)
Los Tineidae, concretamente Tinea deperdella y el género Ceratophaga también forman cámaras de pupación (22), pero tienen una morfología más alargada y canalicular, además de estar siempre asociada a las superficies ricas en queratina (las uñas de manos y pies y el cuero cabelludo) ya que son especies queratinofílicas.
Daño de textiles por Lepidoptera
Las familias Tineidae y Pyralidae son las familias de Lepidoptera más frecuentemente asociadas a cadáveres (23)(24), pero además se caracterizan por ser plagas de gran importancia socioeconómica.
Mientras las larvas de Tineidae muestran preferencia por la lana y otros materiales textiles(25), las de Pyralidae la muestran por alimentos en grano almacenados(26). Nuestro interés se centrará, por tanto, en la familia Tineidae.
Además, las larvas no suelen alejarse demasiado tras alimentarse y se pueden encontrar en las proximidades tanto ejemplares larvarios como pupas o puparios de estos insectos. Esas pupas o puparios tienen aspecto alargado, granuloso y sedoso (27)(28).
Cabe destacar la actividad del género Ceratophaga, que se alimenta casi con exclusividad de la queratina procedente de las uñas, el pelo o las pezuñas de animales muertos(22), pudiendo confundirse con una micosis antemortem, aunque la ausencia de tejido inflamatorio y los cultivos negativos permitirían descartarla.
Lesiones en tejidos blandos por larvas de Diptera y Coleoptera
La piel intacta suele ser una barrera muy efectiva para mantener en el medio externo a las larvas de Diptera y Coleoptera, que suelen preferir los orificios naturales del cuerpo o las heridas para entrar a un plano visceral (29)(30). Ese carácter terso se va perdiendo a medida que la descomposición avanza. Cuando las larvas de Diptera y Coleoptera han finalizado su periodo de alimentación, se deben alejar de la fuente de alimento para pupar.
Para ello, muchas veces horadan la superficie de una piel ya putrefacta para abrirse camino (figura 4).
Estos orificios de salida son de escaso diámetro y de bordes muy bien delimitados, lo que los hace inconfundibles respecto a otras señales antemortem tales como pinchazos o heridas de metralla que, cuando la piel se descompone, tienden a irregularizar sus bordes.
Mordeduras de Formicidae y Dictyoptera
Los Formicidae tienen un carácter omnívoro (31), lo cual presenta un doble inconveniente. Por un lado, pueden depredar sobre estadios inmaduros de otros insectos de interés forense, disminuyendo el número y probablemente la calidad de las muestras recogidas por el entomólogo al darse la posibilidad de que elimine estadios más avanzados de desarrollo útiles para la determinación del intervalo postmortem (32).
Por otro, al alimentarse por regla general en gran número del cadáver, van a introducir mordeduras que pueden ser confundidas con quemaduras u otros traumatismos.
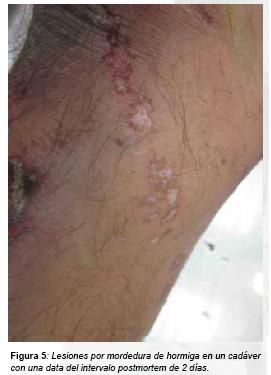
El diagnóstico diferencial de las mordeduras por Formicidae se basa en la presencia de gran número de lesiones próximas en el espacio, apergaminamiento de las zonas donde se produce coalescencia de varias mordeduras y aspecto serpiginoso en la localización de las mismas(33)(34)(figura 5).
Otro signo tipico de la actividad postmortal de las hormigas son la destrucción de los párpados (35). Asimismo por su carácter postmortem, no podremos observar marcas inflamatorias en torno a las soluciones de continuidad de la piel, pudiendo observar estas al microscopio tras una biopsia cutánea.
Los Dictyoptera tienen un patrón de mordedura muy similar (36), aunque con una gran diferencia.
Mordeduras de Crustacea (en cadáveres de medio acuático)
Por otro lado, Gammarus pulex presenta un comportamiento que merece mención aparte(38).
Alteraciones por modificación
Las definiremos como la introducción de cambios en los caracteres identificativos visuales de elementos pertenecientes al cadáver y su contexto, conservando su integridad sin adición de estructuras adicionales.
Manchas de sangre aberrantes por arrastre de larvas de Diptera
Cuando las larvas de Diptera entran en fase migratoria, es muy frecuente que se arrastren alejándose del cuerpo hasta que entran en pupación(39). Ya que están en contacto con fluidos corporales producidos por la descomposición o la sangre procedente de las heridas abiertas, es lógico que dejen tras de sí un reguero que marca el camino seguido en esta fase. Puede inducir al error creando una falsa sensación de marcas de arrastre, aunque son fáciles de distinguir debido a que si se miran con detenimiento se puede observar que la marca de fluido está "anillada" o "segmentada" como producto de la división corporal en segmentos de las larvas.
Desplazamiento de parte o la totalidad de los restos cadavéricos
Por regla general, los vertebrados carroñeros suelen ser los responsables de dispersar partes de los cadáveres. Sin embargo, cuando los insectos tienen un gran tamaño, forman grupos numerosos y presentan mecanismos morfológicos mediante los cuales pueden arrastrar durante un recorrido una cierta cantidad de masa, puede darse el caso de que transporten la totalidad o parte de los restos (en función de la desarticulación y del peso total de la carroña)(40).
Bibliografía
1. Aso, J., Corrons, J., Cobo, J. A. (1998). El intervalo postmortal. Interés médico, policial, legal y forense. España: Masson. [ Links ]
2. Tempelman, A. (1993). Insects and entomologists become police tools in murder investigations. Can Med Assoc J , 148, 601-604. [ Links ]
3. Arnaldos, M. I., García, M. D., Romera, E., Presa, J. J. y Luna, A. (2005). Estimation of postmortem interval in real cases based on experimentally obtained entomological evidence. Forensic Sci Int, 149(1), 57-65. [ Links ]
4. Fernández, S. R. (2000). Temas de Tafonomía. España: Universidad Complutense de Madrid. [ Links ]
5. Haskell, N. H., Hall, R. D., Cervenka, V. J., Clark, M. A. (1997). On the Body: Insect´s life stage presence and their postmortem artifacts. En: Haglund WD & Sorg MH: Forensic Taphonomy. The postmortem fate of human remains.
6. Archer, M. S., Elgar, M. A., Briggs, C. A. y Ranson, D. L. (2006). Fly pupae and puparia as potential contaminants of forensic
7. Linville, J. G., Hayes, J. y Wells, J. D. (2004). Mitochondrial DNA and STR analyses of maggot crop contents: effect of specimen preservation technique. J Forensic Sci, 49(2), 341-344. [ Links ]
8. Clery, J. M. (2001). Stability of prostate specific antigen (PSA) and subsequent Y-STR typing of Lucilia (Phaenicia) sericata
9. Lehane, M. J. (1997). Peritrophic matrix structure and function. Ann Rev Entomol, 42, 525-550. [ Links ]
10. Schroeder, H., Klotzbach, H., Oesterhelweg, L. y Püschel, K. (2002). Larder beetles (Coleoptera, Dermestidae) as an accelerating factor for decomposition of a human corpse. Forensic Sci Int, 127(3), 231-236. [ Links ]
11. Kulikova, N. A. (2004). The comparative morphology of the oral disc labella of the proboscis in flies of the superfamilies Muscoidea and Tachinoidea (Diptera). Entomologicheskoe Obozrenie, 83, 73–86. [ Links ]
12. Palomo, J. L. y Ramos, V. (2003). Papel del médico forense en la Inspección ocular y levantamiento del cadáver. Porpuesta de documento (recomendaciones, guías, normas o protocolos de actuación profesional). Cuad Med Forense, 36, 41-57. [ Links ]
13. Benecke, M. y Barksdale, L. (2003). Distinction of bloodstain patterns from fly artifacts. Forensic Sci Int, 137, 152-159. [ Links ]
14. Fujikawa, A., Barksdale, L., Higley, L.G. y Carter, D. O. (2011). Changes in the morphology and presumptive chemistry
15. Striman, B., Fujikawa, A., Barksdale, L. y Carter, D. O. (2011). Alteration of expirated bloodstain patterns by Calliphora vicina and Lucilia sericata (Diptera: Calliphoridae) through ingestion and deposition of artifacts. J Forensic Sci, 56(S1), 123-
16. Hefti, E., Trechsel, U., Rufenacht, H. y Fleisch, H. (1980). Use of dermestid beetles for cleaning bones. Calcif Tissue Int,
17. Laudet, F. y Antoine, P. O. (2004). Des chambres de pupation de Dermestidae (Insecta: Coleoptera) sur un os de mammifère tertiaire (phosphorites du Quercy): implications taphonomiques et paléoenvironnementales. Geobios, 37, 376-381. [ Links ]
18. Martin, L. D. y West, D. J. (1995). The recognition and use of dermestid (Insecta, Coleoptera)pupation chambers in
19. Kaiser, T. M. (2000). Proposed fossil insect modification to fossil mammalian bone from Plio-Pleistocene hominid-bearing deposits of Laetoli (
20. West, D. J. y Hasiotis, S. T. (2007). Trace Fossils in an Archaeological Context: Examples from Bison Skeletons, Lipscomb
21. Hasiotis, S. T. (2003). Complex ichnofossils of solitary to social soil organisms: understanding theis evolution and roles in
22.Deyrup, M. Deyrup, N. E., Eisner, M. y Eisner, T. (2005). A caterpillar that eats tortoise shells. Am Entomol, 51(4), 245-
23. Introna, F., De Donno ,A., Santoro, V., Corrado, S., Romano, V. y Porcelli, F. et al. (2011). The bodies of two missing children in an enclosed underground environment. Forensic Sci Int, 207(1-3), 40-47. [ Links ]
24. Oliva, A. (2001). Insects of forensic significance in
25. Cox, P. D. y Pinniger, D. B. (2007). Biology, behaviour and environmentally sustainable control of Tineola bisselliella (Hummel) (Lepidoptera: Tineidae). J Stored Prod Res, 43, 2-32. [ Links ]
26. Grieshop, M. J., Flinn, P. W. y Nechols, J. R. (2006). Biological Control of Indianmeal Moth (Lepidoptera: Pyralidae) on Finished Stored Products Using Egg and Larval Parasitoids. J Econ Entomol, 99(4), 1080-1084. [ Links ]
27. Brokerhof, A. W., Morton, R. y Banks, H. J. (1993). Time-mortality relationships for different species and developmental stages of clothes moths (Lepidoptera: Tineidae) exposed to cold. J Stor Prod, 29, 277-282. [ Links ]
28. Pereira, M. (1960). Contribuiçao para o estudo da Tineola bisselliella e seu combate. Estud Ensaios Doc, 78, 1-134. [ Links ]
29. González, A., González, L., Martínez, I., Archilla,F. y Jiménez, G. (2011). Análisis patológico y entomológico de unos restos
30. Pohjoismäki, J.L.O. Karhunen, P. J., Goebeler, S., Saukko, P. y Sääksjärvi,
31. Martínez, M. D., Arnaldos, M. I., Romera, E. y García, M. D. (2002). Los Formicidae (Hymenoptera) de una comunidad sarcosaprófaga en un ecosistema mediterráneo. An Biol, 24, 33-44. [ Links ]
32. Moura, M. O., Carvalho, C. y Monteiro, E. A. (1997). A preliminary analysis of Insects of Medico-legal importance in
33. Campobasso, C. P., Marchetti, D., Introna, F. y Colonna, M. F. (2009). Postmortem artifacts made by ants and the effect of ant activity on decompositional rates. Am J Forensic Med Pathol, 30, 84-87. [ Links ]
34. Byard, R. W. (2005). Autopsy problems associated with postmortem ant activity. Forensic Sci Med Pathol, 1, 37-40. [ Links ]
35. Byard, R. W., James, R. A. y Gilbert, J. D. (2002). Diagnostic problems associated with cadaveric trauma from animal activity. Am J Forensic Med Pathol, 23(3); 238-244. [ Links ]
36. Denic, N., Huyer, D. W., Sinal, S. H., Lantz, P. E., Smith, C. R. y Silver, M. M. (1997). Cockroach: the omnivorous scavenger. Potential misinterpretation of postmortem injuries. Am J Forensic Med Pathol, 18(2), 177-80. [ Links ]
37. Vanin, S. y Zancaner, S. (2011). Post-mortal lesions in freshwater environment. Forensic Sci Int, 212, 18-20. [ Links ]
38. Duband, S.,
39. Kočarek, P. (2001). Diurnal patterns of postfeeding larval dispersal in carrion blowflies (Diptera, Calliphoridae). Eur J Entomol, 98, 117-119. [ Links ]
40. Ururahy, A., Rafael, J. A., Wanderley, R. F., Marques, H. y Pujol, J. R. Coprophanaeus lancifer (Linnaeus, 1767) (Coleoptera, Scarabaeidae) activity moves a man-size pig carcass: Relevant. [ Links ]
+ Instituto de Medicina Legal de Granada Avenida de Madrid s/n, 18012
* Departamento de Medicina Legal y Toxicología. Facultad de Medicina, Universidad de Granada.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}