










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Costarricense
Print version ISSN 0377-9424
Agron. Costarricense vol.35 n.1 San Pedro de Montes de Oca Jun. 2011
Nota técnica
Población de nematodos en forrajes tropicales en dos rangos de altura en el cantón de San Carlos, Alajuela
Plant-parasitic nematode population in tropical forages in 2 altitude range of San Carlos County, Alajuela
Plant-parasitic nematode population in tropical forages in 2 altitude range of San Carlos County, Alajuela
Rodolfo WingChing-Jones1, 2/*, Luís Salazar Figueroa**
1 Proyecto Nº. VI-739-A3-097.
2 Correo electrónico: rodolfo.wingching@ucr.ac.cr
* Escuela de Zootecnia, Centro de Investigación en Nutrición Animal, Universidad de Costa Rica. San José, Costa Rica.
** Escuela de Agronomía, Centro de Investigación en Protección de Cultivos, Universidad de Costa Rica. San José, Costa Rica.
Dirección para correspondencia.
1 Proyecto Nº. VI-739-A3-097.
2 Correo electrónico: rodolfo.wingching@ucr.ac.cr
* Escuela de Zootecnia, Centro de Investigación en Nutrición Animal, Universidad de Costa Rica. San José, Costa Rica.
** Escuela de Agronomía, Centro de Investigación en Protección de Cultivos, Universidad de Costa Rica. San José, Costa Rica.
Dirección para correspondencia.
Resumen
Se caracterizó la población de nematodos fitoparásitos presente en los pastos Sorgo negro (Sorghum almus), Camerún (Pennisetum purpureum var Camerun), King grass (Pennisetum purpureum var King grass), Tanzania (Panicum maximum var Tanzania), Brizantha (Brachiaria brizantha), Toledo (Brachiaria brizantha var Toledo), Estrella africana (Cynodon nlemfuensis), Mombaza (Panicum maximum var Mombaza), Ratana (Ischaemum ciliare) y la leguminosa maní forrajero (Arachis pintoi) en la localidad de San Carlos, Alajuela, de junio a noviembre del 2003. Se tomaron 5 muestras compuestas de suelo y de las raíces contenidas en el volumen de suelo para los forrajes evaluados, excepto para el pasto King grass, el Ratana y el Maní forrajero, en donde el número de muestras compuestas fue de 10 para cada material. Los nematodos Pratylenchus sp., Helicotylenchus sp., Tylenchus sp., Criconemella sp., se identificaron en 8 de los pastos y en la leguminosa evaluada. El género Psilenchus sp., solo se presentó en el pasto Tanzania, mientras que el género Xiphinema sp., en los forrajes Tanzania, Brizantha, Ratana y en el maní forrajero. El género Paratylenchus sp., se contabilizó con los forrajes Tanzania, Brizantha y Ratana. En el pasto Toledo, Camerún y en el maní forrajero se observó Hemicycliophora sp.
En cambio, Meloidogyne sp., se encontró en los pastos Brizantha y Ratana. Los nematodos no fitoparásitos se identificaron en los 9 pastos y la leguminosa evaluada.
Palabras clave: Pastos tropicales, Pratylenchus sp., Sistemas de producción láctea, Pasto de piso, Pasto de corta.
Abstract
Plant-parasitic nematode populations in Black sorghum grass (Sorghum almus), Cameroon grass (Pennisetum purpureum var Camerun), King grass (Pennisetum purpureum var King grass), Tanzania grass (Panicum maximum var Tanzania), Brizantha grass (Brachiaria brizantha), Toledo grass (Brachiaria brizantha var Toledo), African star grass (Cynodon nlemfuensis), Mombaza grass (Panicum maximum var Mombaza), Smutgrass (Ischaemum ciliare) and perennial peanut legume(Arachis pintoi) were characterized during June to November 2003 at San Carlos, Alajuela. Five compound samples of roots and associated soil of with grazing forages were taken, except for King grass, Smutgrass and peanut legume, where the number of samples was 10 for each forage. The nematodes Pratylenchus sp., Helicotylenchus sp., Tylenchus sp., and Criconemella sp., were identified in 8 forages and the perennial peanut legume. Psilenchus sp., was present only in Tanzania grass, while Xiphinema sp., was found in Tanzania grass, Brizantha grass, Smutgrass and perennial peanut legume. Paratylenchus sp., was recorded in Tanzania grass, Brizantha grass and Smutgrass. Hemicycliophora sp., was observed in Toledo grass, Cameroon grass and perennial peanut legume; Meloidogyne sp,. Was found in Brizantha grass and Smurtgrass. Nonplant parasitic nematodes were identified in the perennial peanut legume and all 9 forages evaluated.
Keywords: Tropical grass, Pratylenchus sp., Production systems, Pasture, Fodder grass.
Introducción
La distribución y fluctuación de las poblaciones de nematodos en el sistema productivo de la planta puede depender de la morfología y distribución de las raíces, las características químicas, físicas y biológicas del suelo, prácticas de manejo del suelo y del cultivo, capacidad de asociación con hongos, algas, líquenes y musgos; cambios climáticos que afectan el patrón de lluvias y de temperatura de los microclimas, la capacidad reproductiva y migratoria del mismo, la presencia de parásitos y depredadores según la posición en la cadena trófica (Norton 1978).
Con base en los resultados obtenidos por WingChing-Jones et al. (2008) donde se confirma la presencia de nematodos en 7 forrajes de uso común en sistemas de producción de leche de altura, se hace necesario profundizar en este tema para generar información que describa la población de nematodos presentes en especies forrajeras por debajo de los 1100 msnm, en la comunidad de San Carlos, Alajuela. Otro aspecto importante es que el daño físico que provoca el nematodo durante el proceso de alimentación, permite el ingreso de bacterias, hongos y virus, lo que potencia el daño a la planta, medida en reducción de la productividad del mismo (Dropkin 1980). Por tal motivo, el objetivo de este trabajo fue la de identificar los nematodos presentes en áreas utilizadas para pastoreo y de corta-acarreo de materiales forrajeros que se utilizan entre los 501 a 1100 msnm y por debajo de los 500 msnm.
Materiales y métodos
a. Pastos evaluados y número de muestras tomadas. Se evaluaron 9 especies forrajeras y una leguminosa distribuida entre los 0 a los 1100 msnm, en las localidades de cuidad Quesada, La Tabla, Jabillos y Santa Clara; durante junio a noviembre del 2003. En el rango de altura de 501 a 1100 msnm se trabajo con el pasto de corta Sorgo negro (Sorghum almus), Camerún (Pennisetum purpureum var Camerun) y King grass (Pennisetum purpureum var King grass). Como pasto de piso se empleó el Tanzania (Panicum maximum var Tanzania), Brizantha (Brachiaria brizantha CIAT 6780), Toledo (Brachiaria brizantha var Toledo CIAT 26110) y Estrella africana (Cynodon nlemfuensis). Por debajo de los 500 msnm se utilizó el pasto Mombaza (Panicum maximum var Mombaza), Ratana (Ischaemum ciliare) y la leguminosa maní forrajero (Arachis pintoi). Además en este rango de altura, se repite el muestreo en los forrajes Sorgo negro, Tanzania, Brizantha y Toledo. En promedio se tomaron 5 muestras compuestas de suelo y de las raíces contenidas en el volumen de suelo para los forrajes evaluados, excepto para el pasto King grass, el Ratana y el maní forrajero, en donde el número de muestras compuestas fue de 10 para cada especie.
b. Manejo y análisis de las muestras. En este trabajo se siguió el mismo patrón metodológico descrito por WingChing-Jones et al. (2008) para la escogencia de los sistemas de producción, la toma y análisis de la muestra, en donde cada muestra compuesta procede de 5 submuestras, y cada submuestra se tomó a una profundidad de 0-20 cm con ayuda de un palín, en una área total de muestreo de 5 ha en forrajes de piso y de 2 ha para los pastos de corta. Para maní forrajero se muestrearon en 1,9 ha en total. Las muestras de suelo y de raíces fueron procesadas por el método de cernido y centrifugado en solución azucarada (Alvarado y López 1985), y su lectura se realizó con la ayuda de un microscopio estereoscopio a 45X, iy se identificaron los nematodos a nivel de género, por medio del sistema de clasificación descrito por Maggenti (1991). Los datos de las poblaciones de nematodos determinadas en cada forraje se expresó en términos de número de nematodos.100 ml-1 de suelo o de número de nematodos.100 g-1 de raíces.
Los datos de población de nematodos presentes en las muestras de suelo y de raíces fue organizada por tipo de forraje y rango de altura, y los parámetros de densidad máxima (DM), densidad promedio (DP) así como frecuencia absoluta (FA) descritos por Quintana (1996).
Resultados y discusión
La información obtenida en este trabajo esta organizada en 4 cuadros según el rango de altura y el tipo de forraje.
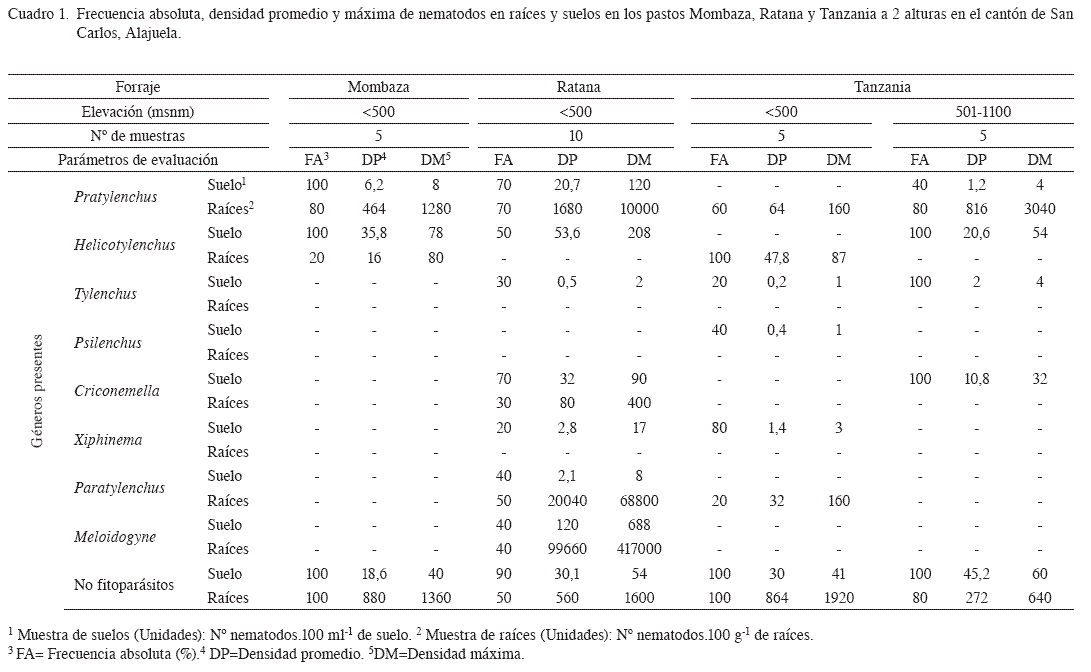
Pasto Ratana (I. ciliare). Se determinó la presencia de Pratylenchus sp., Criconemella sp., Meloidogyne sp., y Paratylenchus sp., en las muestras de raíces, mientras que, en las muestras de suelo, se identificó Pratylenchus sp., Helicotylenchus sp., Tylenchus sp., Criconemella sp., Xiphinema sp., Meloidogyne sp., y Paratylenchus sp. (Cuadro 1). En el caso de los nematodos presentes en las raíces, llama la atención, las altas poblaciones de Pratylenchus sp., Meloidogyne sp., y Paratylenchus sp. Estos géneros se caracterizan por ser endoparásitos en el caso de los 2 primeros y ectoparásito en el caso del Paratylenchus sp., los cuales se alimentan de las raíces del cultivo, lo que reduce la capacidad de absorción de agua y nutrimentos. Es importante describir, que durante los muestreos, el pasto Ratana, no presentó signos de clorosis o sitios en el potrero donde la producción de biomasa se mermara. Sin embargo los problemas del pasto Ratana en los sistemas productivos, es su baja producción de biomasa por hectárea ya que su perfil nutricional es similar al de otros forrajes (Villareal 1992).
Pasto Tanzania y Mombaza (P maximum). El pasto Tanzania presentó 3 géneros de nematodos fitoparásitos en el rango de menor altura (<500 msnm), tanto en las muestras de suelo (Tylenchus sp., Psilenchus sp., y Xiphinema sp.) como en las raíces (Pratylenchus sp., Helicotylenchus sp., y Paratylenchus sp.) (Cuadro 1). En cambio, en el rango de mayor altura (501- 1100 msnm), solo Pratylenchus sp. se identificó en las muestras de raíces. Mientras que en las muestras de suelo se encontró la presencia de Pratylenchus sp., Helicotylenchus sp., Tylenchus sp., y Criconemella sp. En todas las muestras evaluadas de raíces y de suelo, de ambos cultivares de P. maximum, se determinó la presencia de nematodos no fitoparásitos (Cuadro 1). En el caso de las poblaciones recuperadas en raíces, podría estar relacionada a porciones de suelo adheridas a las raíces y proceso de alimentación en raíces en estado de descomposición. En cambio, en el cultivar Mombaza (Cuadro 1), solo se observó la presencia de Pratylenchus sp., y Helicotylenchus sp., en las muestras de raíces y de suelo.
En condiciones de laboratorio, Carneiro et al. (2006), demostraron que el pasto Mombaza y Tanzania presentaron resistencia a M. incognita, M. paranaenses y M. javanica, en donde se observó una reducción en el número de huevos, comparado a un cultivo susceptible, en este caso el tomate. Dichas experiencias, podrían explicar la ausencia de este género en las muestras evaluadas.
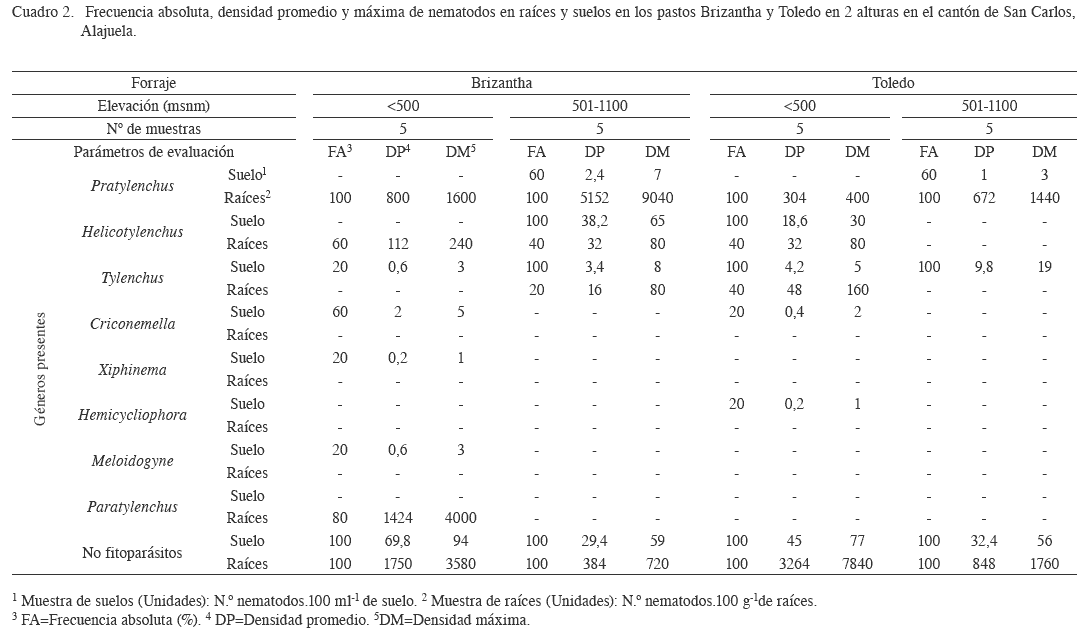
Pasto Brizantha y Toledo (B. brizantha). Para los casos del pasto Brizantha y Toledo se presenta un resumen en el Cuadro 2. Se muestra la presencia de 3 géneros de nematodos fitoparásitos en las muestras de raíces en ambos rangos de altura, en donde Pratylenchus sp. y Helicotylenchus sp., están presentes en ambas alturas, pero Paratylenchus sp., solo esta por debajo de los 500 msnm, mientras que Tylenchus sp. en el rango mayor. En las muestras de suelo, se identificaron 4 géneros de nematodos fitoparásitos, por debajo de los 500 msnm (Tylenchus sp., Criconemella sp., Xiphinema sp., y Meloidogyne sp.) y 3 géneros en el ámbito de altura que comprende los 501 msnm hasta los 1100 msnm (Pratylenchus sp., Helicotylenchus sp., y Tylenchus sp.). Por su parte en el pasto Toledo, se hayó un único género de nematodos fitoparásitos en el rango de altura entre 501 y 1100 msnm, que fue Pratylenchus sp. (Cuadro 2). Para las muestras de suelo y raíces por debajo de los 500 msnm, se encontraron 3: Pratylenchus sp., Helicotylenchus sp., y Tylenchus sp. En ambas alturas con cultivares de pasto Brizanta, se determinó la presencia de nematodos no fitoparásitos en el suelo y las raíces, los cuales podrían participar en el reciclaje de nutrimentos, depredación y descomposición de la materia orgánica (Bardgett et al. 1999).
En un estudio realizado en Brasil por Sharma et al. (2001) sobre la población de nematodos fitoparásitos en pasto Brizantha cv Marandu, se determinó en orden decreciente la presencia de Aphelenchoides subtenuis, Ditylenchus terricolus, Tylenchus sp., Pratylenchus zeae, Aphelenchus avenae, Helicotylenchus dihystera, Meloidogyne sp., y Criconemella sp., resultados que coinciden en esta investigación, con los géneros Pratylenchus sp., Tylenchus sp., Helicotylenchus sp., Meloidogyne sp., y Criconemella sp. Otro estudio realizado por Stanton et al. (1989) en Colombia, hace referencia a Longidorus laevicapitatus, Helicotylenchus sp., Pratylenchus sp., y Xiphinema sp., como nematodos presentes en áreas cultivadas con pasto Brizantha. Al comparar los resultados de Brasil, Colombia y los de esta investigación, se podría considerar al pasto Brizantha hospedero para Pratylenchus sp. y Helicotylenchus sp.
Pasto Camerún y King grass (P. purpureum). En el pasto Camerún se encontró Pratylenchus sp., y Helicotylenchus sp., en las muestras de raíces y de suelo (Cuadro 3). Solo en las muestras de suelo de este forraje, se cuantificaron especímenes de Tylenchus sp., Criconemella sp., y Hemicycliophora sp. Similar comportamiento en géneros de nematodos se obtuvo para el pasto King grass en las muestras de raíces y de suelo excepto Hemicycliophora sp. (Cuadro 3). En ambos cultivares de Pennisetum, sobresalen las poblaciones de Pratylenchus sp., obtenidas. Se desconoce el impacto que este nematodo pueda provocar sobre la productividad y el número de cosechas de estos forrajes. Los resultados obtenidos en esta investigación, contradicen lo informado por WingChing-Jones et al. (2008), en donde indican la no presencia de Pratylenchus sp., en áreas de corta de pasto Camerún y de potreros de pasto kikuyo sobre los 1100 msnm, lo cual se asocia a un factor en las raíces que reduce la colonización del nematodo. Con respecto a la ausencia de Meloidogyne sp., en ambos pastos, se podría asociar al efecto supresivo de este forraje descrito por Coyne et al. (2009).
Pasto Estrella Africana (C. nlemfuensis). En este forraje se identificó a Pratylenchus sp., elicotylenchus sp., y Tylenchus sp., en las muestras de raíces y a Helicotylenchus sp., Tylenchus sp., y Criconemella sp., en las muestras de suelo (Cuadro 3). En comparación con los resultados informados por WingChing-Jones et al. (2008) en potreros de pasto Estrella Africana sobre los 1100 msnm, se contabilizan en ambos trabajos los géneros de nematodos fitoparásitos Pratylenchus sp., Helicotylenchus sp., Tylenchus sp., y Criconemella sp. Igualmente, Sharma (1978) informa de la presencia de Pratylenchus sp. y Tylenchus sp., en el pasto Estrella africana en los cerrados brasileños, con poblaciones de 960 nematodos.100 g-1 raíces y de 160 nematodos. 100 g-1 raíces, lo que podría indicar que este forraje cumple función de cultivo hospedero para estos géneros de nematodos.
En relación con el pasto Cynodon sp., McGroary et al. (2009) informan que Belonolaimus longicaudatus, es el nematodo más importante para el pasto Bermuda (Cynodon dactylon), pues provoca daños severos a las raíces laterales, por reducción en la capacidad de toma de agua, nutrimentos y reduce la tasa de evapotranspiración.
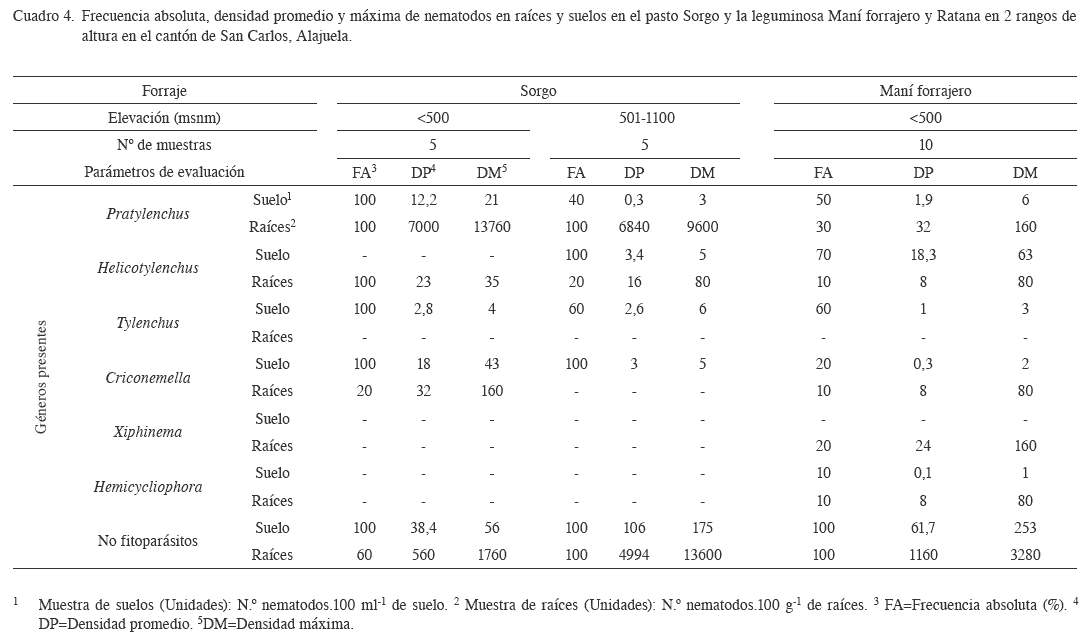
Pasto Sorgo negro (S. almus). En ambas alturas, se determino la presencia de Pratylenchus sp., como nematodo endoparásito, Helicotylenchus sp., Tylenchus sp., Criconemella sp., (Nematodo ectoparásito) y el grupo de los nematodos no fitoparásitos (Cuadro 4). En este sentido, McSorley y Gallaher (1993) informan de poblaciones de Criconemella spp., Meloidogyne incognita, Paratrichodorus minor, Pratylenchus spp., y Xiphinemma spp., en muestras de suelo cultivado con Sorghum bicolor. Estas poblaciones fluctuaron entre 26-378, 11-71, 40-56, 15-140 y 0-46 individuos.100 cc-1 de suelo, respectivamente. Resultados que concuerdan en esta investigación con Pratylenchus sp., y Criconemella sp.
En el caso de Pratylenchus sp., las poblaciones contabilizadas en las muestras de raíces, en los 2 pisos altitudinales evaluados, son frecuentes y mayores a 6800 individuos en 100 g de raíces. Lo que podría indicar que el sorgo es un forraje hospedero sin importar la altura y el tipo de sistema. A la vez, estas poblaciones podrían afectar el rendimiento del cultivo (kg.ha-1.año-1 MS), como la vida útil del mismo, medida en número de cortes (Boschini y Elizondo 2005).
La ausencia de Meloidogyne sp., en las muestras analizadas, podría ser explicada por la resistencia que presenta este cultivo a este género (Carneiro et al. 2007, Silva y Silva 2009), debido a que se emplea en sistemas de rotación con gramíneas de la familia de las brachiarias y con leguminosas como el maní, como medida de supresión (Kokalis-Burelle et al. 2002). Resultados que concuerdan con lo descrito por Coyne et al. (2009), donde cita al sorgo (Sorghum bicolor y Sorghum sudanense) como un material que presenta un efecto supresivo sobre las poblaciones de M. incognita, M. arenaria, M. hapla y M. javanica.
El caso de los recuentos de nematodos no fitoparásitos obtenidos en las muestras de raíces del sorgo (Cuadro 4), podrían estar relacionadas con el proceso de degradación, producto de la senescencia de las raíces en respuesta a la extracción de la biomasa del forraje, por ciclos de pastoreo o por actividad de corta y acarreo para la suplementación en canoa (Georgieva et al. 2005).
Maní forrajero (A. pintoi). Los géneros de nematodos fitoparásitos determinados en este trabajo fueron 5, tanto en las muestras de suelo (Pratylenchus sp., Helicotylenchus sp., Criconemella sp., Hemicycliophora sp., y Tylenchus sp.) como en las de raíces (Pratylenchus sp., Helicotylenchus sp., Criconemella sp., Hemicycliophora sp., y Xiphinema sp.) (Cuadro 4). Aunque, se presentaron valores de poblaciones de nematodos menores a 32 individos en 100.g de raíces en las muestras evaluadas, presentan frecuencias menores al 50%, lo que podría indicar que el maní forrajero, no es un buen cultivo hospedero de nematodos fitoparásitos. Al respecto, Santiago et al. (2001) indican que las leguminosas presentan un antagonismo sobre la población de nematodos fitoparásitos, producto de la liberación de exudados o de un mecanismo de resistencia.
En el caso de los nematodos no fitoparásitos, se encontraron poblaciones promedio mayores a 1000 individuos en 100 g de raíces, las cuales podrían estar favorecidas por la producción y acumulación de biomasa producto del crecimiento del cultivo, como también a la capacidad de fijar nitrógeno atmosférico del maní (Argel y Villareal 1998).
Según Carneiro et al. (2003), el A. pintoi presenta un efecto antagónico sobre las poblaciones de M. incognita, M. arenaria, M. javanica y M. arabicida, cuando fue sembrada en asocio. Iguales resultados encuentran Santiago et al. (2001) sobre la presencia de agallas y masa de huevos de M. incognita en muestras de raíces de A. pintoi y en muestras de raíces de tomate sembrado en suelos con incorporación de A. pintoi como abono verde. Por otro lado, Church et al. (2005) informan de la presencia de un gen de resistencia a M. arenaria en Arachis sp., híbrido. Ambas investigaciones, podrían explicar la no presencia de Meloidogyne sp., en las muestras de raíces y del suelo evaluadas en esta investigación. Por otro lado, en Costa Rica, el uso del maní forrajero como planta de cobertura para proteger la estructura y fertilidad del suelo (Sancho y Cervantes 1997), genera inconvenientes en relación con la productividad como es el caso del banano y el nematodo barrenador Rhadopholus similis, por su efecto en el sistema radical (Araya y Cheves 1997).
Conclusiones
Se confirma la presencia de nematodos fitoparásitos en pastos de uso común en sistemas de producción láctea en la zona de San Carlos, Alajuela por debajo de los 1100 msnm.
Los pastos Sorgo, Tanzania, Mombaza, Brizantha, Toledo, Camerún, King grass, Estrella y Ratana, podrían considerarse como cultivos hospederos del nematodo Pratylenchus sp., pero a la vez se desconoce en las condiciones estudiadas el impacto de este nematodo sobre la productividad y vida útil del cultivo.
Los pastos tropicales evaluados en este trabajo, como el maní forrajero presenta resistencia a ser colonizado por el nematodo Meloidogyne sp., esto debido a que en las muestras evaluadas se determinaron poblaciones promedio menores a 1 nematodo en 100 g de raíces. Caso contrario, las poblaciones obtenidas en el pasto Ratana, permiten considerarlo como un cultivo hospedero.
Las poblaciones de nematodos de los géneros Pratylenchus sp., Meloidogyne sp. y Paratylenchus sp. en el pasto Ratana podrían estar relacionadas con los problemas de persistencia del mismo, debido a la carga residual de estos nematodos en el área a renovar.
Agradecimientos
Al Comité de Educación y Bienestar Social y al Ing. Franklin Ramírez de la Cooperativa de Productores de Leche Dos Pinos R.L., por el apoyo brindado en el desarrollo de este trabajo.
Literatura citada
Alvarado M., López R. 1985. Extracción de algunos nematodos fitoparásitos mediante modificaciones de las técnicas de centrifugación-flotación y embudo de Bareman modificado. Agronomía Costarricense 9(2):175-180. [ Links ]
Araya M., Cheves A. 1997. Poblaciones de los nematodos parásitos del banano (Musa AAA), en plantaciones asociadas con cobertura de Arachis pintoi y Geophilla macropoda. Agronomía Costarricense 21(2):217-220. [ Links ]
Argel P., Villareal M. 1998. Nuevo maní forrajero perenne (Arachis pintoi Krapovickas y Gregory). Cultivar Porvenir (CIAT 18744): Leguminosa herbácea para alimentación animal, el mejoramiento y conservación del suelo y el embellecimiento del paisaje. Ministerio de Agricultura y Ganadería de Costa Rica (MAG), Centro Internacional de Agricultura (CIAT). Boletín técnico. 32 p. [ Links ]
Bardgett R., Cook R., Yeates G., Denton C. 1999. The influence of nematodes on below-ground processes in grassland ecosystems. Plant and Soil 212:23-33. [ Links ]
Boschini C., Elizondo J. 2005. Determinación de la calidad y la producción de sorgo negro forrajero (Sorghum almun) en edades para ensilar. Agronomía mesoamericana 16(101):29-36p. [ Links ]
Carneiro R., Carneiro R., Das Neves D., Almeida M. 2003. Nova raça de Meloidogyne javanica detectada em Arachis pintoi no estado do Paraná. Nematologia Brassileira 27(2):219-221. [ Links ]
Carneiro R., Mônaco A., Lima A., Nakamura K., Moritz M., Scherer A., Santiago D. 2006. Reação de gramíneas a Meloidogyne incognita, a M. paranaensis e a M. javanica. Nematologia Brasileira, Brasilia 30(3):287-291. [ Links ]
Carneiro R., Moritz M., Mônaco A., Nakamura K., Scheser A. 2007. Reação de Milho, Sorgo e Milheto a Meloidogyne incognita, M. javanica e M. paranaensis. Nematologia Brasileira 31(2):9-13. [ Links ]
Church G., Starr J., Simpson C. 2005. A recessive gene for resistance to Meloidogyne arenaria in interspecific Arachis ssp., hybrids. Journal of Nematology 37(2):178-184. [ Links ]
Coyne D., Fourie H., Moens M. 2009. Current and future management strategies in resource poor farming, pp: 444-475. In: R. Perry, M. Moens, J. Starr. Root knot nematodes. CAB International. London, UK. 488 p. [ Links ]
Dropkin V. 1980. Introduction to plant nematology. John Wiley &Sons, Inc. EE.UU. 293 p. [ Links ]
Geogieva S., Chritensen., Stevnbak K. 2005. Nematode succession and microfaunamicroorganism interactions during root residue decomposition. Soil Biology & Biochemistry 37:1763-1774. [ Links ]
Maggenti A.R. 1991. Nemata: higher classification, pp. 147-187. In: W.R. Nickle (ed.) Manual of Agricultural Nematology. Marcel Dekker, Inc., New York, USA. [ Links ]
Mcgroary P., Crow W., Mcsorley R., Giblindavis R., Cisar J. 2009. Seasonal fluctuations of Belonolaimus longicaudatus in Bermudagrass. Nematropica 39:99-110. [ Links ]
Mcsorley R., Gallaher R. 1993. Population dynamics of plant-parasitic nematodes on cover crops of corn and sorghum. Journal of Nematology 25(3):446-453. [ Links ]
Norton D. 1978. Ecology of plant-parasitic nematodes. A Willey interscience. John Wiley & Sons, New York, USA 268 p. [ Links ]
Quintana C. 1996. Elementos de inferencia estadística. Editorial de la Universidad de Costa Rica. San José. Costa Rica. 219 p. [ Links ]
Sancho F., Cervantes C. 1997. El uso de plantas de cobertura en sistemas de producción de cultivos perennes y anuales en Costa Rica. Agronomía Costarricense 21(1):111-120. [ Links ]
Santiago D., Homechin M., Krzyzanowski A., De Carvalho S. 2001. Efeito antagônico de Arachis pintoi sobre Meloidogyne incognita Raça 2 em solo de mata e solo esterilizado. Nematologia Brasileira 25(1):45-51. [ Links ]
Sharma R. 1978. Nematodes associated with gramineous forage crops in Cerrado soil. Soc. Brasileira Nematologia 3:53-54. [ Links ]
Sharma R., Cavalcante M., Valentim J. 2001. Nematóides associados ao capim Brachiaria brizantha cv Marandu no estado do Acre, Brasil. Nematología Brasileira 25(2):217-222. [ Links ]
Silva K., Silva G. 2009. Reação de gramíneas e leguminosas a Meloidogyne mayaguensis. Nematologia Brasileira 33(2):198-200. [ Links ]
Skerman J., Riveros F. 1992. Gramíneas tropicales. Colección FAO. Producción y protección vegetal N° 23. FAO, Roma. 849 p. [ Links ]
Stanton J., Siddiqui M., Lenné J. 1989. Plant parasitic nematodes associated with tropical pasture in Colombia. Nematropica 19(2):169-175. [ Links ]
Vargas G. 2006. Geografía de Costa Rica. EUNED, San José, Costa Rica. 265 p. [ Links ]
Villareal M. 1992. Evaluación comparativa de Ratana (Ischaemum ciliare) como especie forrajera. Agronomía Costarricense 16(1):37-44. [ Links ]
Wingching-Jones R., Salazar-Figueroa L., Flores-Chávez L., Rojas-Bourrillón A. 2008. Reconocimiento de nematodos en pastos tropicales en las comunidades de Sucre y San Vicente, Cantón de San Carlos. Agronomía Costarricense 32(2):129-136. [ Links ]
Correspondnecia a: Rodolfo WingChing-Jones. Escuela de Zootecnia, Centro de Investigación en Nutrición Animal, Universidad de Costa Rica. San José, Costa Rica. / Correo electrónico: rodolfo.wingching@ucr.ac.cr
Luís Salazar Figueroa. Escuela de Agronomía, Centro de Investigación en Protección de Cultivos, Universidad de Costa Rica. San José, Costa Rica.
Recibido: 09/11/10 Aceptado: 30/03/11

{kind=link}
{kind=link}
{kind=link}
{kind=link}