










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Geológica de América Central
On-line version ISSN 0256-7024Print version ISSN 0256-7024
Rev. Geol. Amér. Central n.45 San Pedro de Montes de Oca Dec. 2011
Tafonomía, cronoestratigrafía e implicaciones paleoceanográficas en la turbidita del Paleógeno Temprano (formación vertientes), Cuba
Leidy Menéndez1*, Reinaldo Rojas-Consuegra1, Jorge Villegas-Martín2* & Rafael A. López3*
Abstract
This study focuses on the taphonomy, paleontology, and invertebrate diversity of the Upper Paleocene to Lower Eocene, turbidite deposits of Vertientes Formation, northwest of Ciego de Ávila, Central Cuba. The section exposed is stratified with detritic rocks, heterogeneous litoclasts and bioclasts. The fossil assemblage includes bivalve mollusks, gastropods, equinoderms, corals, crustaceans, icnofossils, orbitoidal foraminifera, ostracods and radiolarians. Age of the deposit was determined by the accumulated planktonic foraminifera assemblage. The taphonomic characterization of the conserved entities suggests processes such as mineralization, recrystallization, sedimentary infilling, disarticulation, fragmentation, encrustation and others, indicating that these conserved entities are alocthonous and have suffered intense processes of transport, taphonomic reelaboration and resedimentation. The depositional sequence was accumulated in association with a slope, in a bathyal environment, influenced by strong precipitations, typical of tropical to subtropical latitudes.
Keywords: Fossil, Early Paleogene, taphonomic, turbidite, Vertientes Formation.
Resumen
Este trabajo se enfoca en el estudio tafonómico y paleontológico de invertebrados en la turbidita de edad Paleoceno Superior - Eoceno Inferior, perteneciente a la Fm Vertientes, en Ciego de Ávila, Cuba Central. La sección estratigráfica está constituida por rocas interestratificadas detríticas, con litoclastos y bioclastos heterogéneos. Se identificaron entidades conservadas de moluscos bivalvos, gastrópodos, equinodermos, corales, briozoos, fragmentos de crustáceos e icnofósiles, y abundantes invertebrados pequeños: macroforaminíferos, ostrácodos y radiolarios. Además, la edad del depósito se determinó con la asociación acumulada de foraminíferos planctónicos. La caracterización tafonómica de la asociación conservada permitió identificar procesos de mineralización, recristalización, rellenamiento, desarticulación, fragmentación, encostramiento, y otros, sugiriendo que la mayoría de los elementos registrados son alóctonos y que han sufrido intensos procesos de transporte, reelaboración y resedimentación tafonómica. La secuencia deposicional fue acumulada en un ambiente batial, con influencia de precipitaciones fuertes, en un ambiente cálido, clima tropical a subtropical.
Palabras claves: Fósil, Paleógeno Temprano, tafonómico, turbidita, Formación Vertientes.
Geologic Setting
This is the first stratigraphic view of an outcrop in the locality of San Vicente, near Jicotea. Vertientes Formation was first described by Lewis (1957). The outcrop provides an excellent exposure across more than 15 m. It extends in a W-E direction between NE of Ciego de Ávila and Florida. Actually, this formation is poorly studied and has a great diversity of invertebrate entities visible at instance. Stratigraphically it is almost seen over Presa Jimaguayú Formation of Maastrichtian Upper Cretaceous, in vertical and lateral transition to Florida Formation of Early Middle Eocene.
According to the evolution of the tectonic plates, the collision between the Cuban Arc and the Bahamas Bank occurred in Paleocene-Middle Eocene time (Pindell et al., 1988). At the latest Paleocene and early Eocene, the remnant ocean to the north of central Cuba remained closed and continued rollback of the oceanic crust flanking the Bahamas, as attested to by increased subsidence rates in the Bahamas at this time, and also by the subduction of the Proto-Caribbean plate (Pindell, et al., 1988). Moreover, Cretacean and older fauna may appear resedimented in Paleogene turbidites, in alocthonous sequences. Several taphonomic evidences denote they suffered an intense transportation and other diagenetic processes.
Methods
The outcrop is located about 5 km to the NE of Ciego de Ávila. The wall and floor of the superficial part of an excavation 1 or 2 m deep were studied.
The characterization of the section took in consideration the composition, texture, geometry of beds, and fossil composition; also, the taphonomic state of conserved entities.
The registered entities collected were diverse and abundant. Many macroinvertebrates were found, and it was possible to identify a large number of invertebrates of Maastrichtian age.
For the micropaleontologic study 22 samples were collected along the outcrop. The samples were washed at the laboratory to facilitate disaggregation of microfossils. Later, the specimens were placed in plates for their identification and to determine the age of the accumulated association of planktonic foraminifera of each sample, and for the deposit.
Lithostratigraphy
The section exposed is composed of detritic rocks, heterogeneous litoclasts and bioclasts in composition and size, where the beds slightly tend to the SE, in monoclinal form.
Four lithologic units are described from bottom to top. The 1st and basal unit is about 3 m thick, with massive polimictic sand, some stratigraphic inclined beds, and planar cross stratification. Subredounded litho- and bioclast appear. The 2nd unit is about 2 m thick, separated from the first by an erosive surface of the basal part. Charks of clastic and interstratified rocks with breccias and sand in rhythmic form are present. Perforations like bioerosion structures proper of intertidal littoral facies can be seen. The 3rd unit is about 1 m thick, mostly horizontal, discordant with the lower unit. A section shows parallel stratification, and it is compound with breccias and conglomerates, with a sand-clay matrix. Rounded carbonate bioclasts appear. The 4th and upper part is about 3 to 5 m thick, with marls, lutites (siltstones), and sand-clay stratified charks.
Taphonomic Characterization
The production and accumulation of fossil remains aided in interpreting the fossil register of the sedimentary section. Most important is not only the taxonomic identification, but how the taxobiothemes appear in the chronologic context.
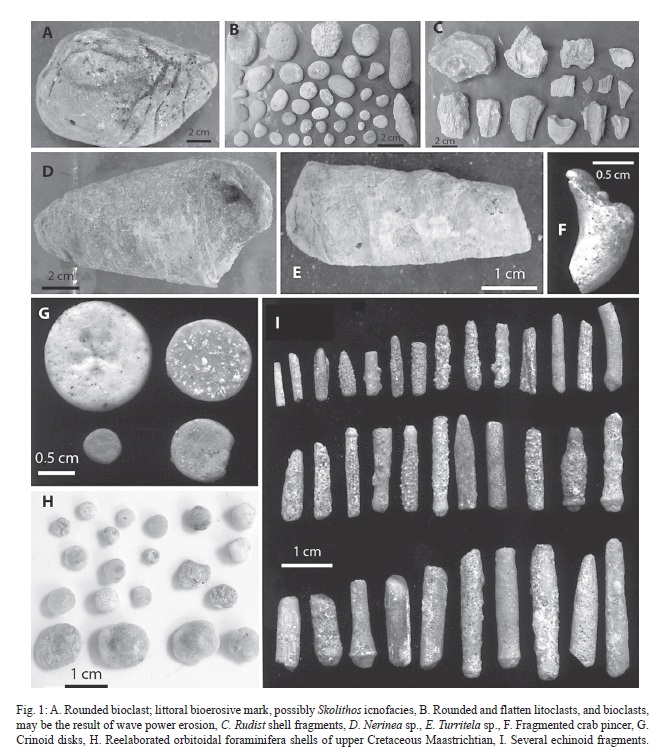
Several reelaborated entities show regular to bad conservation. Bioclasts were common at the stratigraphic section, mainly rounded, variable in size, and irregular (Fig. 1 - A y B). The genus Entobia isp. , Fig. 1 - A found inside the section has poor preservation. It forms several external openings, circular to subcircular, occasionally irregular, connected inside by galleries. This icnogenus results from the erosive action of clinoid sponges and drilling poliquets (Bromley & D’Alessandro, 1984), indicating shallow sea. The taphonomic alteration, polish and rounded features surely resulted from a long and intense process of transport, and finally alocthony.
Abundant rudists were found too. Some detritus fragments and internal molds of radiolitic rudists permited the identification of Titanosarcolites giganteus, an index species of Late Maastrichtian (Rojas-Consuegra, 1998; 2005). Other bivalve mollusks were represented by rare fragments and shells.
Also included well recrystallized and run down nerideids, turritelid shells, and fragments of a crustacean pincer, which clearly indicate lithification previous to reelaboration (Fig. 1 C, D y E). This is the first fossil crab so far known from Upper Cretaceous Maastrichtian of Cuba, (Rojas- Consuegra, 2009); nevertheless, gallery systems attributed to crab excavation activity in the icnogenus Thalassinoides have been studied (Pszczólkowski, 2002). Run down and polished crinoid disks of diverse species were present too (Fig. 1 - G e I), as well as rare corals and bryozoan fragments.
Several and diverse macroscopic benthic foraminifera shells were found (Fig. 1 - H) showing a taphonomic reelaboration, polished surfaces, ventral wearing signs, mineral crusts, and sedimentary refill. Coloration and texture show that recrystallization occurred before reelaboration, preserving the entity during unearth, transport, and accumulation. Their primary features disappeared.
The taxobiotheme assemblage shows that the fossil material collected belongs to tidal – subtidal shallow lithofacies of a consolidate Maastrichtian carbonate platform in this region, all of them really transformed by the intense transport.
Microfossils suffer several modifications in the fossilization process not as far as in the process suffered by macroscopic fossils, but often difficult to distinguish. The resedimentation is common in micropaleontology, mostly because of the small size and weight, permitting horizontal and lateral dispersion, causing necroplanktonic drift, and also the alocthony of the conserved entities. The reelaborated or accumulated state of the taxa depend on the taphonomic interpretation, and this discussion emerged generally because there is no easy visual criterium in micropaleontology to distinguish one from the other. Isotopic studies could be the principal tool to distinguish accumulated from reelaborated entities, although dealing with problems like diagenesis and ontogenesis (D´Hondt & Zachos, 1993; Smith & Nederbragt, 1997). Other powerful tool to distinguish indirectly the probabilities of reelaboration may be the quantitative study based on the relative abundance of the several genera; through a comparative study with other near similar outcrops, or simply looking for different colors or conservation states between the species in the sample (Arz et al., 1999).
In the micropaleontologic analysis disarticulated ostracod shells, rare and small recrystallized radiolarians with several foraminifers, were found together. Some radiolarians and the genus Chiloguembelina that often appear in cool deep sea at upper latitudes (Arenillas et al. 2000) may have suffered a drift.
Mostly foraminifers appeared well preserved; although it is evident they suffered taphonomic processes like shell fragmentation, recrystallization, crusts, and mineralization.
The authoctony has been asserted by the accumulated assemblage of planktonic foraminifera. The icnogenus Planolites isp., which appeared over sand - mud substrate as long structures, straight to incline form, with a similar stuffing of the hosting rock was present, but poorly preserved. Unfortunately, this genus does not permit to date the sediments. It has been documented in several sedimentary contexts and ages, attributed to the action of worms of wide environmental range between shallow to deep water.
Biostratigraphy and Paleoceanographic Implications
Microfossils can be accumulated, resedimentated or reelaborated, but could be mixed in the same stratigraphic bed, not representing the same cronostratigraphic zone.
The section shows a diversity of microfossils: planktonic foraminifera, macroforaminifers, radiolarians, and ostracods. In some samples clearly identified, but not in others, the taphonomic disturbance suffered difficults the identification of the registered species. Moreover, the fossil record is partial, incomplete, and slant stratigraphically.
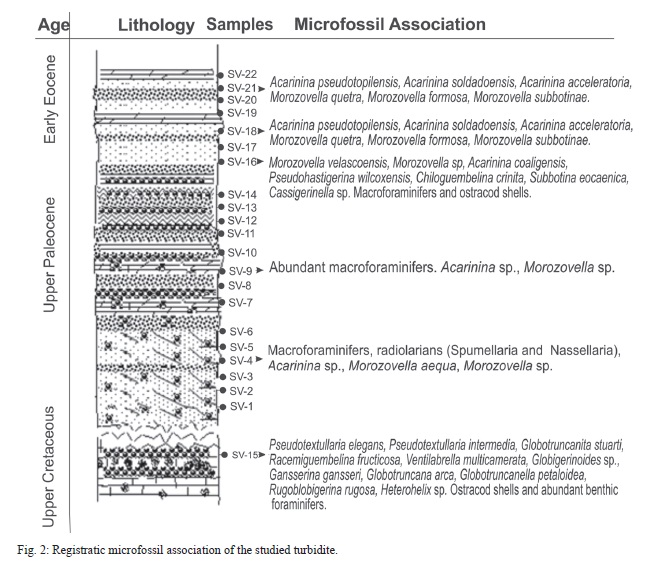
The accumulated assemblage proves that, contrary to the first view, the turbidite set during Upper Paleocene to Lower Eocene (Fig. 2).
At the lower part of the section, except in sample SV-15, the abundance of species was really poor, and it has been difficult to find well preserved planktonic foraminifers to date the samples, fragmented simple shells only permitting the identification of genera. The section groups biozones P4c Acarinina soldadoensis Subzone (Late Paleocene, Late Thanetian) to P7 Morozovella formosa formosa Zone (Middle - Early Eocene, Ypresian).
The subzone P4c Acarinina soldadoensis was recognized with the appearance of: Acarinina soldadoensis; also recorded were Morozovella aequa, Acarinina coaligensis, and Subbotina eocaenica; others appear masked by the taphonomic attributes. In samples SV-16, SV-18 taxa recorded were Morozovella formosa formosa (P7 zone), A. wilcoxensis, A. pseudotopilensis, A. camerata, A. acceleratoria, M. quetra, M. formosa, M. lensiformis, M. subbotinae, and Pseudohastigerina wilcoxensis. The sampling method used does not permit to distinguish the stratigraphic horizons.
In this period the genera Morozovella and Acarinina were really abundant. The existence of a minimal expanded oxygenic zone has been confirmed by differences in δ13C of plancktonic foraminifers of superficial (Morozovella) and middle to deep (Subbotina) waters, with the increase of δ13C, at the increase of bathymetry (Shackleton et al., 1985). The highest levels of δ13C, probably of the entire Cenozoic, are at the Upper Paleocene (Shackleton et al., 1985; Stott et al., 1990), suggesting the enrichment of δ13C is caused by productivity increase of superficial sea waters. In this micropaleontological sampling the abundance of the genera Morozovella and Acarinina, and the juvenile stage, denotes a eutrophic environment.
From isotopic studies it has been concluded that the latitudinal and bathymetry distribution of planktonic species change with evolution; for example, Morozovella and Acarinina are typically from superficial waters, although they can move to deeper levels (Arenillas et al. 2000).
Sample SV-15 shows a typical association of Upper Cretaceous, probably corresponding to Gansserina gansseri zone. The accumulated association is well preserved; Pseudotextullaria elegans, P. intermedia, Globotruncanita stuarti, Racemiguembelina fructicosa, Ventilabrella multicamerata, Gansserina gansseri, Globotruncana arca, Globotruncanella petaloidea, Rugoblobigerina rugosa, and Heterohelix sp. were recorded. This sample may belong to subyacent Jimaguayú Formation; consequently, the Cretaceous-Paleocene boundary in this part of the section could be found.
According to the geologic map, the sedimentary units of Upper Cretaceous Campanian - Maastrichtian (Durán and Jimaguayú Formations) and from Paleogene (Vertientes and Florida Formations) occur transgressively: SW to NW from the positive relief of Cretaceous volcanites, Caobilla Formation (Iturralde-Vinent, 1981). The study section has siliciclastic and calcareous clastic - detritus rocks from a shallow platform source, probably from these Upper Cretaceous emerged terrains.
The Early Eocene climate is tropical to subtropical with the influence of intense precipitations bringing an intense meteorization (Uriarte, 2009). The area functioned like a drain of atmospheric CO2, with cool sea draft and abundant nutrients from the rivers to the sea.
Conclusions
The taphonomic and micropaleontologic data from de turbidite section of Vertientes Formation in western Cuba having a great diversity of clastic and bioclastic sediments make know an epivolcanic terrain of a carbonated Maastrichtian marine cover accumulated in a slump, in a bathyal environment of Upper Paleocene to Early Eocene. The section probably has formed under a cyclic regimen of precipitation, aided by alluvial contribution inside the turbidite, reflected in the repetitive erosive surfaces described, and ending in a general transgression to cover the emerging terrain.
Registered entities show an intense transport process with a variable state of conservation. The macroinvertebrates association found, reelaborated entities, corresponds to the Maastrichtia carbonated platform; meantime, the microfossils were accumulated in Upper Paleocene to Early Eocene. An Upper Maastrichtian association was found. It possibly belongs to the Jimaguayú Formation that underlies Vertientes Formation. In fact, it suggests that the Cretaceous–Paleogene boundary may be preserved in this place.
Acknowledgments
This paper is a contribution to the Proyect: “Biodiversidad paleontológica del archipiélago cubano: bases cartográficas y conservacionistas”, of Paleontology and Biogeography Department, Natural Museum of Natural History of Cuba. We are grateful to the Center of Oil Research of Cuba for the processing of the micropaleontologic samples and to the reviewer Dr. Gilberto Silva Taboada for many comments and suggestions.
References
ARENILLAS, I., ALEGRET, L., ARZ, J.A. & MOLINA, E., 2000: El uso didáctico de los foraminíferos en la enseñanza de ciencias de la Tierra: su distribución paleoceanográfica en el Tránsito Cretácico Terciario.- Enseñanza de Ciencias de la Tierra, Univ. de Zaragoza, 8(2): 108-118. [ Links ]
ARZ, J.A., ARENILLAS I., MOLINA, E. & DEPUIS C., 1999: Los efectos tafonómicos y ´Signor-Lipps´ sobre la extinción en masa de foraminíferos planctónicos en el límite Cretácico/Terciario de Elles (Tunicia).- Rev. Soc. Geol. España, 12(2): 251-268. [ Links ]
BROMLEY, R.G. & D’ALESSANDRO, A., 1984: The ichnogenus Entobia from the Miocene, Pliocene and Pleistocene of southern Italy.- Riv. Ital. Paleontol. Strat. 90(2): 227-296. [ Links ]
D´HONDT, S. & ZACHOS J.C., 1993: On stable isotopic variation and earliest Paleocene planktonic foraminifera.- Paleoceanography, 8(4): 527-547. [ Links ]
LEWIS Jr., P.D. 1957. Reconnaissance Geology of the Vertientes -Francisco area.- 235 págs. Arch. Nac. Fondo Geol. MINBAS, La Habana [Inf. Interno]. [ Links ]
PINDELL, J.L., CANDE, S.C., PITMAN, W.C., ROWLEY, D.B., DEWEY, J.F.,LABRECQUE, J. & HAXBY, W., 1988: A plate-kinematic framework for models of Caribbean evolution.- Tectonophysics, 155: 121-138. [ Links ]
PSZCZÓLKOWSKI, A., 2002: Crustacean burrows from Upper Maastrichtian Ddeposits of south-central Cuba.- Bull. Polish Acad. of Earth Sci. 50(2): 147-163. [ Links ]
ROJAS-CONSUEGRA, R., 1998: Facies marinas costeras en sedimentos carbonatados del Maastrichtiano en Ciego de Ávila, Cuba central.- III Congr. de Geología y Minería, La Habana, Cuba: 263-266. [ Links ]
ROJAS-CONSUEGRA, R., 2005: Los rudistas de Cuba: Estratigrafía, tafonomía, paleoecología y paleobiogeografía.- 264 págs. Inst. Sup. Politécnico “José Antonio Echevarría”, La Habana Tesis Ph.D. [ Links ]
ROJAS-CONSUEGRA, R., 2009: Sinopsis del registro fósil de Cuba.- href= http://www.redciencia.cu/webpaleo.html Consulta: 11 de noviembre de 2010. [ Links ]
SHACKLETON, N.J., CORFIELD, R.M. & HALL, M.A., 1985: Stable isotope data and ontogeny of Paleocene planktonic foraminifera.- J. Foram. Res. 15: 321-336. [ Links ]
SMITH, J. & NEDERBRAGT, A.J., 1997: Analysis of the El Kef blind test II.- Marine Micropaleontol. 29: 95-100. [ Links ]
STOTT, L.D. & KENETT, J.P., 1990: Antarctic Paleogene planktonic foraminifera biostratigraphy: ODP Leg 113 Sites 689 and 690.- Proc. ODP Sci. Res. 113: 549-569. [ Links ]
URIARTE, A., 2009. Historia del clima en la Tierra.- 403 págs. Eusko Jaurlaritza, Gobierno Vasco. [ Links ]
*Correspondencia: Leidy Menéndez: Museo Nacional de Historia Natural de Cuba. AMA-CITMA. Obispo 61, Plaza de Armas, Habana Vieja; CP10100, Cuba. Autora para contacto: maia@mnhnc.inf.cu
Reinaldo Rojas-Consuegra: Museo Nacional de Historia Natural de Cuba. AMA-CITMA. Obispo 61, Plaza de Armas, Habana Vieja; CP10100, Cuba
Jorge Villegas-Martín: Instituto de Ecología y Sistemática. AMA-CITMA, Carretera de Varona km. 31/2, Capdevilla, Boyeros, AP8029, CP10800, Ciudad La Habana, Cuba
Rafael A. López: Instituto de Geología, UNAM, Ciudad Universitaria, México, D. F, 04510, México
1Museo Nacional de Historia Natural de Cuba. AMA-CITMA. Obispo 61, Plaza de Armas, Habana Vieja; CP10100, Cuba
2Instituto de Ecología y Sistemática. AMA-CITMA, Carretera de Varona km. 31/2, Capdevila, Boyeros, AP8029, CP10800, Ciudad La Habana, Cuba
3Instituto de Geología, UNAM, Ciudad Universitaria, México, D. F, 04510, México
*Autora para contacto: maia@mnhnc.inf.cu
(Recibido: 27/05/2011; aceptado: 28/11/2011)


{kind=link}
{kind=link}