











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los escarabajos coprófagos de la subfamilia Scarabaeinae (Coleoptera: Scarabaeidae), han sido ampliamente estudiados, debido a su gran diversidad y abundancia. Además, este grupo ha sido empleado como modelo para entender la respuesta de las comunidades biológicas a la modificación del hábitat, debido a que estas especies muestran una alta sensibilidad ante cambios en la estructura de la vegetación, o en la disponibilidad de recursos y las variaciones ambientales (Nichols et al., 2007; Pulido-Herrera et al., 2003; Spector, 2006).
Para el estudio de este grupo de insectos se utilizan distintos métodos de captura, por ejemplo; trampas de caída, trampas de intercepción de vuelo, necrotrampas, captura manual, entre otros. Sin embargo, la trampa de caída es la más utilizada en los estudios de diversidad de Scarabaeinae por su alta efectividad, bajo costo de operación y fácil manejo en campo; además, esta permite realizar muestreos estandarizados que permiten la comparación entre sitios o temporadas distintas (Barreto de Andrade et al., 2011; Knapp, et al., 2016). En los bosques colombianos se ha empleado esta técnica como la principal herramienta de muestreo; no obstante, se utilizan diferentes tamaños de cebo y tiempos de operación. Por ejemplo, estudios en el bosque seco tropical (BST) y en bosques de galería han empleado cebos de 25 g durante 48 h, capturando entre 22-61 especies de escarabajos (Escobar, 1997; Escobar, 2000; Noriega et al., 2007; Pulido-Herrera et al., 2003). Por otro lado, Amézquita et al. (1999) emplearon 1 g de excremento durante 24 h en un bosque de vega, y capturaron 32 especies; mientras en bosques pluviales se utilizaron trampas con 2 g de excremento durante 48 h en donde se capturó nueve especies (Castillo et al., 2011). En el BST del Caribe colombiano, Solís et al. (2011) y Noriega et al. (2012), emplearon cebos de 30 g durante 48 h, y capturaron entre 13-35 especies; mientras otros autores emplearon atrayentes de 25 g durante 24 h, registrando una riqueza similar de especies (Barraza et al., 2010; Martínez et al., 2009; Martínez et al., 2010; Rangel-Acosta & Martínez-Hernández, 2017; Rangel-Acosta et al., 2016a; Rangel-Acosta et al., 2016b).
Lo anterior evidencia una falta de estandarización de la técnica de muestreo de escarabajos (Finn & Giller, 2000); lo cual podría causar errores o discordancias en las estimaciones de la composición y estructura de los ensamblajes de este grupo de insectos. De esta manera, se hace necesario desarrollar investigaciones para evaluar el impacto de utilizar distintas metodologías en relación a la cantidad de excremento necesario para cebar las trampas de caída, así como el tiempo de permanencia de las trampas en ecosistemas distintos, como el BST. Además, la necesidad de aportar esta información surge del hecho de que tradicionalmente en el BST se han aplicado metodologías diseñadas para bosques de las zonas húmedas y bosque de la región andina del país (Cultid, 2007; Escobar & Chacón, 2000; Martínez et al., 2009), lo cual podría estar incidiendo en el rendimiento de las trampas y en la eficiencia de captura de los escarabajos coprófagos. Lo anterior es importante si se tiene en cuenta que las condiciones ambientales son totalmente contrastantes entre los ecosistemas mencionados anteriormente y el BST en el Caribe colombiano. Por ejemplo, en este último durante el periodo seco se presentan altos valores de temperaturas ambiente, del suelo e intensidad lumínica y una baja humedad ambiente y del suelo (Barraza et al., 2010; Martínez et al., 2010; Rangel-Acosta et al., 2016a, Rangel-Acosta et al., 2016b; Rangel-Acosta & Martínez-Hernández, 2017; Solís et al., 2011).
Los resultados de esta investigación permitirían obtener datos sobre el tiempo de operación y el tamaño del cebo, que se requieren al momento de la utilización de la trampa de caída, con el fin de realizar estimaciones confiables de la riqueza, abundancia, diversidad y biomasa de especies para este grupo de insectos (Buzas & Hayek, 1996; Gotelli & Colwell, 2001). De acuerdo con lo descrito, plateamos que una mayor cantidad de atrayente utilizado y un mayor tiempo de operación de las trampas de caída garantizaría una mejor estimación de la riqueza, abundancia, biomasa y diversidad de escarabajos coprófagos capturados en un fragmento de BST en el Caribe colombiano. Teniendo en cuenta lo anterior, la presente investigación analizó el efecto del tamaño del cebo y el tiempo de operación de la trampa de caída en la efectividad de la captura de escarabajos coprófagos en un fragmento de BST en La Reserva Campesina la Flecha (RLF), San Jacinto, Colombia.
Materiales y métodos
Área de estudio: El estudio se realizó en un fragmento de bosque seco tropical (BST) en la Reserva Campesina la Flecha (RLF), la cual se encuentra localizada en el municipio de San Jacinto, Bolívar (9°50’59.48” N & 75°10’00.83” W). Esta reserva presenta una altitud que oscila entre 300-500 m y una extensión de 149 ha. Con respecto a las condiciones climatológicas, la reserva hace parte de la subunidad climática D2, de acuerdo a la clasificación propuesta por Rangel-Ch y Carvajal-Cogollo (2012), con una precipitación promedio de 1 972 mm. El régimen de precipitaciones es bimodal tetraestacional, caracterizado por dos periodos secos comprendido entre diciembre-marzo y junio-julio con rangos de pluviometría entre 7.6-98 mm/mes. Los dos periodos de lluvias comprenden los meses de abril-mayo y septiembre-noviembre con rangos de precipitación entre 173-369 mm/mes (Márquez & Martínez, 2020; Villareal et al., 2019). El promedio de la temperatura es de 35.92 ± 1.10 °C en época seca y 24.18 ± 0.80 °C en lluvias, mientras que la humedad relativa oscila entre 42.18 ± 1.47 % en seca y 93.06 ± 2.24 % en lluvias (Márquez & Martínez, 2020). Durante la época seca se presenta un déficit del agua y muchas fuentes de aguas como arroyos y quebradas se secan, permaneciendo pequeños charcos de agua únicamente en las zonas profundas de las quebradas que sirven como reservorios (Villareal et al., 2019).
La vegetación predominante es la característica del BST, con condiciones higrotropofíticas (Pizano & García, 2014). El estrato arbóreo alcanza entre 20-25 m, con elementos emergentes que pueden llegar hasta los 35 m en las zonas adyacentes a la quebrada La Flecha. Los géneros representativos son: Ceiba, Pseudobombax, Tabebuia, Philodendron, Anthurium, Acrocomia y Aristolochia, además de las especies Spondias mombin, Crescentia cujete, Pseudobombax septenatum, Anacardium excelsum, Parinari pachyphylla, Justia bracteosa, Malvaviscus aboreus, Myrmecodendron costarriscense, Petiveria alliiaceae, Cordia alba, Mangifera indica, Guazuma ulmifolia, Pereskia quisqueyana, Bursera simaruba y Hura crepitans.
Diseño y método de captura: El estudio se realizó entre marzo y septiembre de 2015, con el fin de abarcar la época seca (marzo y abril) y de lluvias (agosto-septiembre). Por época se realizaron dos muestreos con un intervalo de 15 días entre uno y otro, para un total de cuatro muestreos. Para cada muestreo a lo largo de un transecto lineal de 800 m se establecieron cinco estaciones separadas aproximadamente 200 m, en cada una se delimitó un cuadrado (50 x 50 m) y en cada uno de sus ángulos se instaló una trampa de caída; para un total de cuatro trampas por estación y 20 trampas por muestreo.
Cada trampa consistió de un vaso plástico de 947 ml enterrado a ras del suelo, y se llenó un tercio de su volumen con solución preservante (para un litro de preservante: 250 g de sal, 600 ml de agua, 300 ml alcohol al 70 %, 10 ml detergente líquido). Al lado de cada trampa se fijó una varilla de 5 mm de diámetro con uno de sus extremos modificados en forma de gancho, en donde se sujetó el cebo. Con el fin de proteger las trampas de las lluvias, se instaló un plato desechable invertido (30 cm de diámetro), suspendido con tres estacas en la parte superior de las trampas, a una altura aproximada de 30 cm sobre el suelo.
Por estación se instalaban cuatro trampas, tres de estas con diferentes cantidades de excremento y una trampa control (sin cebo). Los criterios de selección de los tamaños de los cebos fueron los siguientes: La selección del tamaño pequeño (P = 34.6 ± 5.4 g), obedece a que la mayoría de trabajos que cuantifican la diversidad de escarabajos usan como referencia un tamaño entre 25-35 g, por lo que este rango fue asumido como un tamaño pequeño. Por otro lado, la elección del tamaño grande (G = 114.9 ± 12.3 g) es debido a que uno de los mayores aportadores de recurso de mayor frecuencia y mayor tamaño en el BST son los Alouatta seniculus por lo que antes de la ejecución de este trabajo fueron localizadas varias de estas deposiciones y pesadas obteniendo un peso promedio de ± 120 g, peso similar al que se usó en este trabajo. Por último, la selección del tamaño mediano (M = 53.8 ± 5.9 g) atiende a un peso intermedio entre los dos anteriores tamaños. El cebo utilizado en este trabajo consistió en una mezcla en proporciones iguales de excremento humano y carroña (vísceras de pollo), la cual se encontraba envuelta en gaza. El uso de esta combinación de atrayentes garantiza la captura de especies de escarabajos tanto coprófagas como necrófagas (Rangel-Acosta & Martínez-Hernández, 2017); además asegura la construcción de los cebos, debido a una mayor disposición de recurso para su elaboración (Marsh et al., 2013). Los pesos de los cebos se estandarizaron con una balanza analítica portátil (Pesola 200 ± 0.002 g).
Las trampas se instalaron a las 6:00 am, para su posterior revisión a las 24 h y 48 h. Las muestras recolectadas se lavaron, preservaron y rotularon con las indicaciones de Villareal et al. (2004). Adicionalmente, se monitoreó la pérdida de peso de los diferentes cebos con intervalos de 6 horas, la pérdida de peso fue calculada al tomar las medidas del peso inicial y el peso final de cada uno de los diferentes tamaños de cebo. Por otra parte, también fueron medidas las variables ambientales: temperatura y humedad ambiente (Datalogger Extech RHT50) e intensidad lumínica (Luxómetro Extech LT 300). La frecuencia de toma de datos fue de 30 minutos y la selección de estas variables fue debido a que estas pueden tener un efecto directo sobre el deterioro de la calidad del cebo.
Trabajo de laboratorio e identificación: Los coleópteros capturados se identificaron en el laboratorio de Zoología de la Universidad del Atlántico con un estereoscopio Leica S8 APO y se utilizó las claves taxonómicas propuestas por Medina y Lopera (2000), Vaz-de-Mello et al. (2011) hasta el nivel de género, y hasta nivel de especies con las claves propuestas por Delgado y Kohlmann (2007), Edmonds y Zidek (2010), Génier (2009), González et al. (2009), Kohlmann y Solís-Blanco (1997), Kohlmann y Solís-Blanco (2001), y Solís y Kohlmann (2002), Solís y Kohlmann (2004). Una vez confirmadas las especies se agruparon según disposición final del alimento (relocalización), en tres grupos funcionales teniendo en cuenta lo propuesto por Doube (1990): rodadores (R), cavadores (C) y residentes (Re). Además, se clasificaron según su tamaño en: pequeños (P) (< 10 mm) y grandes (G) (> 10 mm) según lo propuesto por Peck y Howden (1984) y Escobar (2004).
Para estimar la biomasa de los escarabajos coprófagos se seleccionó un máximo de 10 individuos por cada especie recolectada. Para la selección de cada individuo, el único criterio de selección aplicado fue que estuviera con sus partes completas. Posteriormente, cada individuo fue secado en un horno a una temperatura constante de 60 °C por 72 h y se registró el peso seco de cada uno por especie con una balanza analítica electrónica (Modelo Ohaus Adventurer de 220 ± 0.00001 g). Luego, se promedió el peso seco por especie, y se multiplicó por la abundancia total de cada especie (Lobo, 1993). Para aquellas especies con abundancias inferiores a 10 individuos, la biomasa fue calculada con los ejemplares disponibles.
Análisis de datos: Se determinó la riqueza, abundancia y biomasa de escarabajos coprófagos por tamaño del cebo, tiempo de operación y épocas climáticas (seca y lluvias). Para cada uno de estos atributos del ensamblaje se calculó el promedio y los respectivos intervalos de confianza al 95 %. Posteriormente se realizó comparaciones visuales basadas en la superposición de los intervalos de confianza (Cumming et al., 2007). Cabe resaltar, que los datos de abundancia a las 48 h fueron la sumatoria de los individuos recolectados en los dos tiempos de operación (24 y 48 h). Las trampas control fueron excluidas de los análisis debido a que ninguna de estas trampas presentó captura.
Por otro lado, fueron realizados modelos lineales generalizados (MLG) acompañados de análisis de varianza (ANOVA), con el fin de determinar el efecto de las variables predictoras (época, tamaño del cebo y tiempo de operación) sobre los atributos de la comunidad de escarabajos copronecrófagos (riqueza, abundancia y biomasa). La riqueza, abundancia y biomasa se ajustaron a distribuciones Poisson, Binomial Negativa y Gamma respectivamente, con una función de enlace logarítmica. Se eligieron los modelos con menor valor de AIC (Akaike, 1974). Estos análisis se realizaron con los paquetes STATS (Bolar, 2019), MASS (Ripley et al., 2020) y CAR (Fox et al., 2020).
Con el fin de determinar si los diferentes tamaños de los cebos presentan diferencia en la captura de la riqueza, abundancia y biomasa de escarabajos de cuerpo pequeño y grande en las diferentes épocas de los muestreos se utilizó una prueba de Kruskal-Wallis acompañado de una prueba pareada de U Mann-Whitney. Este análisis fue realizado con el paquete STATS (Bolar, 2019). La cobertura del muestreo se calculó utilizando el estimador de cobertura propuesto por Chao y Jost (2012). La diversidad se calculó mediante un análisis de interpolación y extrapolación propuesto por Chao et al. (2014), utilizando los números de Hill (Hill, 1973) en unidades del número efectivo de especies (Jost, 2006). Se calcularon los valores de diversidad verdadera, 0 D (riqueza especies), 1 D (especies comunes [el exponencial del índice de Shannon]) y 2 D (especies muy abundantes [el inverso del índice de Simpson]). Estos índices de diversidad fueron comparados usando los intervalos de confianza al 95 % (Cumming et al., 2007). El análisis fue realizado con el paquete iNEXT (Hsieh et al., 2020).
Los cambios en composición y estructura del ensamblaje entre tamaños de cebo para las épocas climáticas fueron analizados empleando un análisis de escalamiento multidimensional no métrico (nMDS) utilizando como medida de distancia el índice de similitud de Bray-Curtis binario, acompañado a posteriori por un análisis de similitud (ANOSIM). Este análisis fue realizado con los paquetes MASS (Ripley et al., 2020) y vegan (Oksanen et al., 2019). Para los análisis multivariados (nMDS y ANOSIM) se excluyeron las especies con pocos representantes (singletons y doubletons) para evitar sesgos por submuestreo. Por último, para determinar si la pérdida de peso de los diferentes tamaños de cebo es afectada por las variables humedad ambiente, temperatura ambiente e intensidad lumínica, se utilizó un análisis de correlación de Spearman, empleando el paquete psych (Revelle, 2020). Todos los análisis fueron realizados con el programa R versión 4.0.3 (R Development Core Team, 2017).
Resultados
Se capturaron 4 563 individuos agrupados en diez géneros y 27 especies. Las especies más abundantes fueron Onthophagus lebasi (Boucomont, 1932) con 866 individuos, seguida por Uroxys boneti (Pereira & Halffter, 1961) con 813 y U. deavilai (Delgado & Kohlmann, 2007) con 599. Estas tres especies aportaron el 50 % de la abundancia total capturada. La cobertura del muestreo por tamaño del cebo y época climática fue alta (99-100 %) (Tabla 1).
Tabla 1 Abundancia, biomasa (entre paréntesis, mg), y grupos funcionales de los escarabajos coprófagos por época climática y tamaño del cebo. / Table 1. Abundance, biomass (in parentheses, mg), and functional groups of dung beetles per climatic season and bait size.
| Especies-Tamaño | Época seca | Época de lluvia | Total | ||||
| P | M | G | P | M | G | ||
| Escarabajos de cuerpo pequeño (< 10 mm) | |||||||
| Cavadores | |||||||
| Canthidium sp. 1 | 1 (8) | 5 (39) | 4 (31) | 3 (23) | 8 (62) | 4 (31) | 25 (194) |
| Onthophagus acuminatus Harold, 1880 | 0 (0) | 0 (0) | 0 (0) | 2 (8) | 3 (12) | 24 (96) | 29 (116) |
| Onthophagus crinitus Harold, 1869 | 8 (43) | 18 (96) | 40 (213) | 64 (341) | 64 (341) | 157 (837) | 351 (1 871) |
| Onthophagus landolti Harold, 1880 | 4 (68) | 26 (442) | 23 (391) | 50 (850) | 55 (935) | 135 (2 295) | 293 (4 981) |
| Onthophagus lebasi Boucomont, 1932 | 0 (0) | 3 (16) | 3 (16) | 170 (923) | 187 (1 015) | 503 (2 731) | 866 (4 701) |
| Onthophagus marginicollis Harold, 1880 | 4 (160) | 42 (1 680) | 86 (3 440) | 50 (2 000) | 61 (2 440) | 205 (8 200) | 448 (17 920) |
| Onthophagus sp 1 | 2 (15) | 17 (128) | 6 (45) | 22 (166) | 9 (68) | 32 (242) | 88 (664) |
| Trichillum pilosum Robinson, 1948 | 0 (0) | 0 (0) | 0 (0) | 12 (180) | 22 (330) | 29 (435) | 63 (945) |
| Uroxys boneti Pereira & Halffter, 1961 | 33 (154) | 97 (453) | 106 (495) | 154 (719) | 206 (961) | 217 (1 013) | 813 (3 795) |
| Uroxys deavilaiDelgado y Kolhmann, 2007 | 47 (78) | 114 (190) | 140 (233) | 93 (155) | 82 (137) | 123 (205) | 599 (998) |
| Rodadores | |||||||
| Agamopus lampros Bates, 1887 | 0 (0) | 0 (0) | 0 (0) | 14 (87) | 9 (56) | 62 (384) | 85 (527) |
| Canthon acutus Harold, 1868 | 17 (157) | 45 (416) | 24 (222) | 5 (46) | 4 (37) | 17 (157) | 112 (1 035) |
| Canthon cyanellus Leconte, 1859 | 0 (0) | 1 (12) | 0 (0) | 4 (48) | 3 (36) | 3 (36) | 11 (132) |
| Canthon juvencus Harold, 1868 | 0 (0) | 5 (13) | 8 (20) | 3 (8) | 4 (10) | 6 (15) | 26 (66) |
| Canthon lituratus Germar, 1813 | 0 (0) | 0 (0) | 1 (7) | 1 (7) | 0 (0) | 0 (0) | 2 (14) |
| Canthon mutabilis Lucas, 1857 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 2 (24) | 2 (24) |
| Canthon septemmaculatus Latreille, 1811 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 () | 1 (16) | 1 (16) |
| Canthon sp. 1 | 0 (0) | 0 (0) | 0 (0) | 1 (5) | 1 (5) | 2 (10) | 4 (20) |
| Canthon subhyalinus Harold, 1867 | 6 (41) | 63 (428) | 50 (340) | 1 (7) | 2 (14) | 1 (7) | 123 (837) |
| Residentes | |||||||
| Eurysternus impressicollis Castelnau, 1840 | 15 (182) | 21 (254) | 17 (206) | 16 (194) | 37 (448) | 57 (690) | 163 (1 974) |
| Escarabajos de cuerpo grande (> 10 mm) | |||||||
| Cavadores | |||||||
| Dichotomius agenor Harold, 1869 | 0 (0) | 0 (0) | 0 (0) | 9 (2 241) | 15 (3 735) | 24 (5 976) | 48 (11 952) |
| Dichotomius sp. 1 | 0 (0) | 0 (0) | 0 (0) | 41 (30 463) | 42 (31 206) | 53 (39 379) | 136 (10 1048) |
| Phanaeus hermes Harold, 1868 | 0 (0) | 0 (0) | 0 (0) | 6 (507) | 3 (254) | 9 (761) | 18 (1 522) |
| Rodadores | |||||||
| Canthon aequinoctialis Harold, 1868 | 19 (961) | 51 (2 581) | 57 (2 884) | 2 (101) | 0 (0) | 1 (51) | 130 (6 578) |
| Deltochilum guildingii Westwood, 1835 | 0 (0) | 1 (1 594) | 2 (3 188) | 1 (1 594) | 3 (4 782) | 5 (7 970) | 12 (19 128) |
| Residentes | |||||||
| Eurysternus caribaeus Herbst, 1789 | 0 (0) | 9 (928) | 6 (619) | 3 (309) | 9 (928) | 10 (1 031) | 37 (3 815) |
| Eurysternus mexicanus Harold, 1869 | 11 (293) | 17 (452) | 10 (266) | 8 (213) | 10 (266) | 22 (585) | 78 (2 075) |
| Número de especies | 12 | 17 | 17 | 25 | 23 | 26 | 27 |
| Número de individuos | 167 | 535 | 583 | 735 | 839 | 1 704 | 4 563 |
| Biomasa (mg) | 2 160 | 9 722 | 12 616 | 41 195 | 48 078 | 73 177 | 186 948 |
| Cobertura del muestreo | 99 % | 100 % | 100 % | 99 % | 100 % | 100 % | - |
| Número de especies pequeñas | 7 | 10 | 10 | 16 | 15 | 17 | 20 |
| Número de especies grandes | 5 | 7 | 7 | 9 | 8 | 9 | 7 |
| Número de individuos de especies pequeñas | 81 | 321 | 345 | 437 | 453 | 1 158 | 2 795 |
| Número de individuos de especies grandes | 86 | 214 | 238 | 298 | 386 | 546 | 1 768 |
| Biomasa (mg) de especies pequeñas | 906 | 4 167 | 5 659 | 5 767 | 6 907 | 17 424 | 40 830 |
| Biomasa (mg) de especies grandes | 1 254 | 5 555 | 6 957 | 35 428 | 41 171 | 55 753 | 146 118 |
Abreviaturas: Pequeño (P), mediano (M) y grande (G). / Abbreviations: Small (P), medium (M), and large (G).
Teniendo en cuenta el tamaño del cebo y la época, la mayor abundancia promedio se capturó con el cebo grande durante la época de lluvia, siendo las especies O. lebasi con 503 individuos y Onthophagus marginicollis (Harold, 1880) con 205 individuos las que realizaron el mayor aporte. Con respecto a la biomasa, se estimó un peso seco total de 186 950 mg, aportado principalmente por la especie Dichotomius sp. 1 (101 048 mg). La mayor biomasa se presentó en el cebo grande durante la época de lluvia, siendo las especies Dichotomius sp. 1 (3 940 mg) y O. marginicollis (8 000 mg) las que aportaron mayor biomasa.
Los mayores valores de riqueza, abundancia y biomasa promedio de escarabajos coprófagos se presentaron durante la época de lluvia y los menores en la época seca. La mayor riqueza promedio se observó durante la época de lluvia, en el cebo grande, a las 48 h (19.6 especies). Este valor es significativamente alto y difiere de todos los valores observados, exceptuando al cebo grande en la época de lluvia a las 24 h (Fig. 1A). En contraste, valores significativamente bajos de riqueza se presentaron en la época seca en el cebo pequeño a las 24 h, dichos valores difieren de los otros observados, exceptuando al cebo pequeño en la época de lluvia a las 24 h. Por otra parte, este análisis también muestra que la riqueza de especies no cambia entre horas de muestreo para los mismos tamaños de cebo independientemente de la época (Fig. 1A).
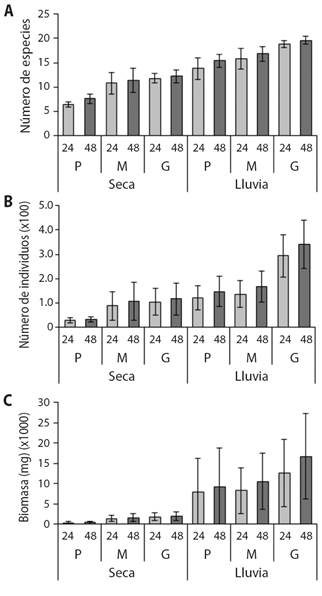
Fig. 1 Variación de los promedios de la A. riqueza, B. abundancia y C. biomasa para los diferentes tiempos de operación y tamaños del cebo por épocas climáticas (media ± I.C.). Abreviaturas: Pequeño (P), mediano (M) y grande (G). Fig. 1. Variation of the averages of A. richness, B. abundance and C. biomass for the different operating times and bait sizes per climatic season (mean ± I.C.). Abbreviations: Small (P), medium (M), and large (G).
Un patrón similar al descrito anteriormente fue observado para la abundancia, donde el mayor valor se registró en la época de lluvia en el cebo grande a las 48 h (340.8 individuos) y el menor en el pequeño durante la época seca a las 24 h (30.4 individuos) (Fig. 1B). En el caso de la biomasa, el mayor valor se registró en el cebo grande a las 48 h durante la época de lluvia (1 670 mg). Este valor no presentó diferencias con los otros tamaños de cebos y horas de exposición para la época de lluvia, pero sí difiere de los valores observados para la época seca (Fig. 1C). Por otra parte, los promedios de biomasa más bajos (400 mg) se presentaron en el cebo pequeño a las 24 h durante la época seca y fueron significativamente diferentes al resto de observaciones. Es importante resaltar que independientemente de la época, la abundancia y la biomasa no presentaron diferencias entre horas de muestreo para un mismo tamaño del cebo.
Con los Modelo Lineales Generalizados (MLG), se logró determinar que la época climática y el tamaño del cebo, así como su interacción presentaron un efecto positivo y estadísticamente significativo (P < 0.05) sobre la riqueza, abundancia y biomasa de escarabajos, presentándose mayores valores en la época de lluvia y en el cebo grande. Por el contrario, el tiempo de operación de las trampas y su interacción con los demás factores no presentaron un efecto estadísticamente significativo sobre la riqueza, abundancia y biomasa de escarabajo (P > 0.05) (Tabla 2). Teniendo en cuenta lo anterior, el tiempo de exposición de las trampas fue excluido de los análisis posteriores.
Tabla 2 Anova del Modelo lineal Generalizado (MLG) sobre la riqueza, abundancia y biomasa de escarabajos coprófagos. / Table 2. Anova of the Generalized Linear Model (GLM) on the richness, abundance, and biomass of dung beetles.
| Riqueza | |||
| lr x 2 | gl | Valor-P | |
| Época | 50.4 | 1 | < 0.0001* |
| Tamaño | 17.8 | 2 | 0.0001* |
| Tiempo | 0.97 | 1 | 0.32 |
| Época*Tamaño | 3.61 | 2 | 0.16 |
| Época*Tiempo | 0.004 | 1 | 0.94 |
| Tamaño*Tiempo | 0.28 | 2 | 0.86 |
| Época* Tamaño*Tiempo | 0.04 | 2 | 0.97 |
| Abundancia | |||
| Época | 63.3 | 1 | < 0.0001* |
| Tamaño | 48.0 | 2 | < 0.0001* |
| Tiempo | 1.68 | 1 | 0.19 |
| Época*Tamaño | 11.9 | 2 | 0.002* |
| Época*Tiempo | 0.05 | 1 | 0.82 |
| Tamaño*Tiempo | 0.06 | 2 | 0.97 |
| Época* Tamaño*Tiempo | 0.01 | 2 | 0.99 |
| Biomasa | |||
| Época | 98.0 | 1 | < 0.0001* |
| Tamaño | 14.5 | 2 | 0.007* |
| Tiempo | 0.68 | 1 | 0.40 |
| Época*Tamaño | 7.08 | 2 | 0.02* |
| Época*Tiempo | 0.06 | 1 | 0.80 |
| Tamaño*Tiempo | 0.01 | 2 | 0.99 |
| Época*Tamaño*Tiempo | 0.04 | 2 | 0.97 |
Abreviaturas: Razón de verosimilitud x 2 (lr x 2) y Grados de libertad (g.l.). Valores p-significativos (P < 0.05) con asterisco. / Abbreviations: Likelihood ratio x2 (lr x2) and Degrees of freedom (g.l.). Significant p-values (P < 0.05) with asterisk.
El análisis de diversidad (iNEXT), muestra un patrón temporal en la riqueza de especies (0 D), observándose valores bajos en la época seca, los cuales aumentaron en la época de lluvia (Fig. 2A), un patrón que no se conserva para la diversidad de especies comunes (1 D) y especies muy abundantes (2 D) (Fig. 2B, Fig. 2C). Para la época de lluvias, los tamaños de cebos presentaron valores de riqueza, de diversidad de especies comunes y dominantes muy similares (Fig. 2); mientras que, para la época seca, el cebo pequeño presentó valores bajos para los tres órdenes de la diversidad, con respecto a los cebos medianos y grandes los cuales presentaron los valores más altos (Fig. 2).
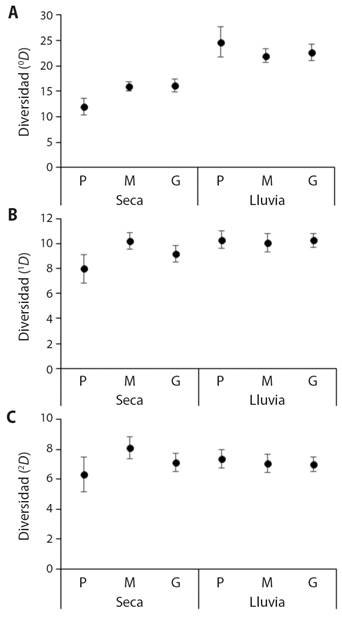
Fig. 2 Análisis de la diversidad A. 0 D, B. 1 D y C. 2 D de escarabajos coprófagos, para los diferentes tamaños del cebo y épocas climáticas. Abreviaturas: Pequeño (P), mediano (M) y grande (G). / Fig. 2. Analysis of the diversity A. 0 D, B. 1 D and C. 2 D of dung beetles for different bait sizes and climatic seasons. Abbreviations: Small (P), medium (M), and large (G).
Con el análisis de escalamiento multidimensional no métrico (nMDS), se observó la formación de dos grupos que difieren en la estructura del ensamblaje de escarabajos. El primero formado por la época de lluvia y el segundo por la época seca (ANOSIM, R = 0.89, P = 0.001) (Fig. 3). Durante la época de lluvia, los tamaños del atrayente no difieren en la estructura (R = 0.19, P = 0.06) y forman un solo grupo. Sin embargo, en la época seca estos presentaron diferencias (R = 0.38, P = 0.03). Se aprecia que el cebo pequeño forma un grupo independiente que guarda cierto grado de similitud con el cebo mediano (R = 0.04, P = 0.31), pero difiere del cebo grande (R = 0.35, P = 0.04). Por su parte el cebo mediano y grande forman un grupo con estructura del ensamblaje similar (R = -0.06, P = 0.61).
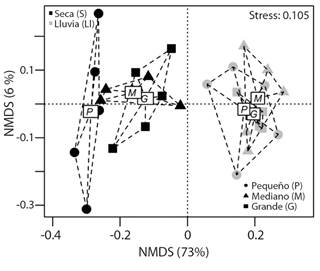
Fig. 3 Análisis de escalamiento multidimensional no métrico (nMDS), basado en una matriz de similitud de Bray-Curtis, teniendo en cuenta la abundancia de distintas especies de escarabajos coprófagos en los diferentes tamaños del cebo y las épocas climáticas. / Fig. 3. Non-metric multidimensional scaling analysis (nMDS), based on a Bray-Curtis similarity matrix, taking into account the abundance of different species of dung beetles in different bait sizes and the climatic seasons.
Al analizar la riqueza, abundancia y biomasa de escarabajos teniendo en cuenta el tamaño del cuerpo de los escarabajos (pequeño y grande), encontramos patrones diferentes a los observados cuando se analizan sin utilizar esta categorización. Por ejemplo, la riqueza de escarabajos de cuerpo pequeño difiere entre tamaños de cebos para la época seca (H = 5.02, P = 0.02), siendo los cebos grandes y mediano los que capturaron más especies de escarabajos pequeños. Durante la época de lluvia todos los cebos capturan similar número de escarabajos de cuerpo pequeño (H = 2.09, P = 0.19) (Tabla 1, Tabla 3). En cuanto a la riqueza de escarabajos de cuerpo grande, los diferentes tamaños de cebo capturaron similar número de especies tanto en época seca (H = 2.16, P = 0.233) como en época de lluvias (H = 2.69, P = 0.09) (Tabla 1, Tabla 3).
Tabla 3 Resultados de la prueba pareada U Mann-Whitney para detectar diferencia entre los tres tamaños de cebo con respecto a la riqueza, abundancia y biomasa, teniendo en cuenta el tamaño de los escarabajos coprófagos. / Table 3. Results of the U Mann-Whitney paired test to detect difference between the three sizes bait with respect to richness, abundance and biomass, taking into account the size of the dung beetles.
| Ítems | Época de seca | Época de lluvia | ||||
| P-M | P-G | G-M | P-M | P-G | G-M | |
| Número de especies pequeñas | U = 2 363, P = 0.03 | U = 2 250, P = 0.01 | U = 2 700, P = 0.62 | U = 2 700, P = 0.61 | U = 2 438, P = 0.08 | U = 2 505, P = 0.20 |
| Número de especies grandes | U = 855, P = 0.14 | U = 855, P = 0.15 | U = 1 013, P = 0.99 | U = 900, P = 0.76 | U = 910, P = 0.09 | U = 922, P = 0.89 |
| Número de individuos especies pequeñas | U = 2 295, P = 0.01 | U = 2 182, P = 0.004 | U = 2 704, P = 0.65 | U = 2 703, P = 0.67 | U = 2 149, P = 0.001 | U = 2 240, P = 0.002 |
| Número de individuos especies grandes | U = 768, P = 0.03 | U = 782, P = 0.04 | U = 1 009, P = 0.97 | U = 898, P = 0.34 | U = 715, P = 0.01 | U = 814, P = 0.10 |
| Biomasa (mg) especies pequeñas | U = 3 846, P = 0.01 | U = 3 655, P = 0.003 | U = 4 431, P = 0.60 | U = 3 702, P = 0.02 | U = 3 717, P = 0.03 | U = 4 494, P = 0.96 |
| Biomasa (mg) especies grandes | U = 574, P = 0.01 | U = 558, P = 0.01 | U = 787, P = 0.90 | U = 613.5, P = 0.04 | U = 459, P = 0.0009 | U = 641, P = 0.12 |
Abreviaturas: pequeño (P), mediano (M) y grande (G). / Abbreviations: small (P), medium (M), and large (G).
En cuanto a la abundancia de escarabajos de cuerpo pequeño esta difiere entre los diferentes tamaños de cebo tanto para la época seca (H = 6.39, P = 0.01) como para la época de lluvia (H = 7.37, P = 0.02). Para ambos periodos la abundancia capturada con el cebo grande supera ampliamente a la abundancia del cebo pequeño, mientras que solo supera la abundancia del cebo mediano durante la época de lluvia. Por su parte el cebo mediano solo supera al cebo pequeño en número de individuos durante el periodo seco (Tabla 1, Tabla 3). La abundancia de especies de escarabajos de cuerpo grande, entre tamaños de cebo difirió tanto para la época seca (H = 4.90, P = 0.04) como en lluvias (H = 6.12, P = 0.04), con un patrón similar al observado anteriormente, durante la época seca los cebos grandes y medianos superan en número de individuos al cebo pequeño; sin embargo, en la época de lluvias solo el cebo grande supera al pequeño en abundancia (Tabla 1, Tabla 3).
Respecto a la biomasa aportada por los escarabajos de cuerpo pequeño esta presentó diferencias para los tres tamaños de cebos, tanto en la época seca (H = 6.85, P = 0.008), como en la época de lluvias (H = 6.388, P = 0.04). Los cebos medianos y grandes superan en biomasa al cebo pequeño, mientras que estos no difieren en biomasa. Patrón similar fue observado para la biomasa aportada por los escarabajos de cuerpo grande, para la época seca (H = 6.79, P = 0.01) y la época lluviosa (H = 10.93, P = 0.003) (Tabla 1, Tabla 3).
En general especies como: Agamopus lampros, Eurysternus impresicollis, Onthopagus acuminatus, O. crininitus, O. landolti, O. lebasi, O. marginicollis y U. deavilai fueron abundantes en el cebo grande. Por otro lado, especies como Canthon acutus, C. aequinoctialis, C. subhyalinus, C. juvencus, Deltochilum guildingui, Dichotomius agenor, E. caribaeus, E. mexicanus, Trichillum pilosum y Uroxys bonetti fueron abundantes en los cebos medianos y grandes; mientras que en el cebo pequeño no hay especies que lo tipifiquen teniendo en cuenta su abundancia. Es de resaltar que con el cebo grande durante la época seca y de lluvias se capturo una abundancia que corresponde al 50.1 % de total de la abundancia capturada en este estudio, entre tanto con el cebo mediano se registró el 30.1 % y en el pequeño el 19.8 %.
En general, los grupos funciónales de escarabajos se ajustan a un patrón temporal, presentándose mayores valores de riqueza, abundancia y biomasa durante los periodos de lluvias y los menores durante la época seca; con excepción de los grandes rodadores (G.R), los cuales presentaron mayor abundancia durante la época seca (Fig. 4). Para la época de lluvias, en todos los tamaños del cebo se capturaron valores similares de riqueza, abundancia y biomasa de grupos funcionales. Sin embargo, para la época seca esta panorámica cambia totalmente y son los cebos mediano y grande donde se registraron los mayores valores de riqueza, abundancia y biomasa de la mayoría de los grupos funcionales de escarabajos (Fig. 4).
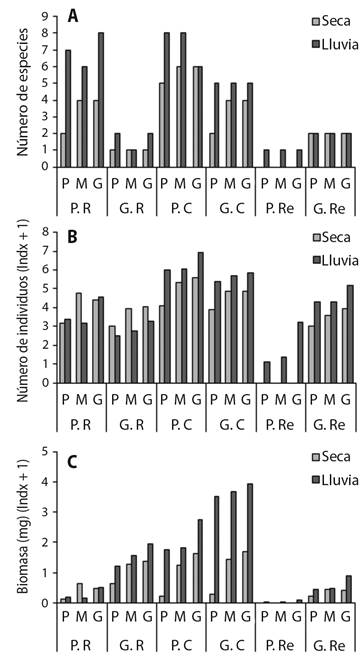
Fig. 4 Variación de la A. riqueza, B. abundancia y C. biomasa de los gremios funcionales de Scarabaeinae, según los hábitos de relocalización del alimento. Abreviaturas: Pequeño rodador (P.R), pequeño cavador (P.C), pequeño residente (P.Re), grande rodador (G.R), grande cavador (G.C) y grande residente (G.Re). / Fig. 4. Variation of A. richness, B. abundance and C. biomass of the functional guilds of Scarabaeinae, according to the relocation habits of the food. Abbreviations: Little rollers (P.R), little tunnelers (P.C), small resident (P.Re), big rollers (G.R), big tunnelers (G.C), and big resident (G.Re).
En general, las variables como intensidad lumínica y temperatura ambiental presentaron valores altos en la época seca; los cuales disminuyeron en la época de lluvias, caso contrario a lo registrado con la humedad ambiente (Tabla 4). Por otro lado, independientemente de la época la pérdida de peso de los atrayentes es mayor en el cebo de mayor tamaño; ya que a las 48 h de instaladas en campo pierden entre el 46 y 47.5 %, mientras el mediano pierde entre 42 y 43.5 %. El cebo pequeño es el que presenta la menor perdida transcurridas 48 h, solo entre 31 y 34.5 % (Fig. 5). Con el análisis de correlación de Spearman, se determinó que solo la intensidad lumínica tiene un efecto positivo y significativo sobre pérdida de peso de los cebos (Ro = 0.86, P = 0.03); mientras la humedad del ambiente (Ro = 0.64, P = 0.08) y la temperatura del ambiente (Ro = 0.74, P = 0.06) muestran una correlación positiva no significativa.
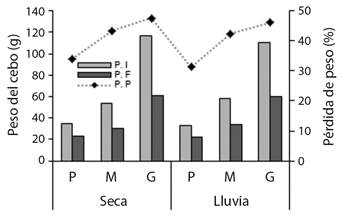
Fig. 5 Pérdida de peso promedio de los diferentes tamaños de cebo para cada época de muestreo. Abreviatura: pequeño (P), mediano (M), grande (G), peso inicial (P. I), peso final (P. F) y pérdida de peso (P. P). / Fig. 5. Average weight loss of the different bait sizes for each sampling time. Abbreviation: small (P), medium (M), large (G), initial weight (P. I), final weight (P. F) and weight loss (P. P).
Tabla 4 Valores promedio (± desviación estándar) de las variables abióticas medidas. / Table 4. Average values (± standard deviation) of the abiotic variables measured.
| Variables abióticas | Seca | Lluvias | ||||
| P | M | G | P | M | G | |
| I. L (Klux) | 20.49 (± 10.40) | 22.04 (± 9.61) | 20.39 (± 19.52) | 0.50 (± 0.29) | 1.80 (± 3.12) | 0.46 (± 0.39) |
| H. A (%) | 55.34 (± 3.98) | 54.92 (± 6.31) | 56.16 (± 8.04) | 66.70 (± 11.43) | 75.21 (± 1.69) | 69.42 (± 1.18) |
| T. A (°C) | 31.11 (± 1.80) | 31.56 (± 1.33) | 30.78 (± 1.85) | 28.98 (± 4.11) | 29.52 (± 2.92) | 31.59 (± 0.70) |
Abreviaturas: intensidad lumínica (I. L), humedad ambiente (H. A) y temperatura ambiente (T.A). / Abbreviations: light intensity (I. L), ambient humidity (H. A) and ambient temperature (T.A).
Discusión
El número de especies de escarabajos (27 especies) que se capturó en la Reserva La Flecha se encuentra en el rango reportado en otros trabajos (17-35 especies) realizados en la región del Caribe colombiano (Barraza et al., 2010; Delgado-Gómez et al., 2012; Martínez et al., 2010; Navarro et al., 2011; Noriega et al., 2013; Noriega et al., 2016; Rangel-Acosta et al., 2012; Rangel-Acosta et al., 2016a; Rangel-Acosta et al., 2016b; Solís et al., 2011) y en el resto del país (Amézquita et al., 1999; García & Pardo, 2004; Noriega et al., 2012; Noriega et al., 2015). De esta manera, este parche de bosques seco en el departamento de Bolívar conserva un porcentaje importante de la fauna de Scarabaeinae presente para el Caribe colombiano y para el país en general. Esto demuestra la importancia de este fragmento de bosque para la conservación de la diversidad biológica, razón por la cual se hace necesario incluirlo en un sistema de protección, conservación y monitoreo permanente; debido a que actualmente afronta la problemática de la expansión agrícola y ganadera, al igual que otros fragmentos de BST en Colombia (Pizano & García, 2014). Lo anterior posiblemente cause a futuro problemas más severos como la pérdida del ecosistema; incidiendo directamente en la disminución y extinción local especies de escarabajos, así como la pérdida de los servicios ecosistémicos aportados por este grupo. Entre los resultados más relevantes de este trabajo, se destacan la temporalidad mostrada por el ensamblaje de escarabajo, la nula diferencia en riqueza, abundancia y biomasa entre horas de operación para un mismo tamaño del cebo, los altos valores de riqueza, abundancia, diversidad y biomasa en los cebos de mayor tamaño sobre todo en la época seca y los resultados diferenciales en cuanto al tamaño de los escarabajos capturados cuando se usan cebos de diferentes tamaños. Con relación al primer resultado, varios trabajos han demostrado la temporalidad de los ensamblajes de escarabajos coprófagos de los bosques secos, mostrando claramente altos valores de riqueza, abundancia y diversidad de especies durante la época de lluvias, las cuales decrecen drásticamente en la época seca (Barraza et al., 2010; Navarro et al., 2011; Solís et al., 2011; Rangel-Acosta et al., 2016a; Rangel-Acosta et al., 2016b). Estos cambios son atribuidos básicamente a la sincronización de los escarabajos con la dinámica del BST (Rangel-Acosta & Martínez-Hernández, 2017). Por ejemplo, durante la época de lluvias, los valores altos de variables como cobertura vegetal, la humedad ambiental y la humedad del suelo favorecen el establecimiento y permanencia del ensamblaje de escarabajos. Además, en este periodo aumenta la disponibilidad del recurso (excremento), así como su tiempo de permanencia sin degradarse, lo cual favorece los altos valores de diversidad reportados para este periodo (Rangel-Acosta & Martínez-Hernández, 2017; Rangel-Acosta et al., 2018). En el caso de la época seca, los altos valores de la temperatura ambiente, la temperatura del suelo y la intensidad lumínica combinados con una baja cobertura vegetal imposibilita la permanencia de los escarabajos, reduciendo drásticamente su diversidad. Adicionalmente, se presenta una disminución en la disponibilidad del recurso (excremento) así como su permanencia, debido a que las variables ambientales mencionadas favorecen la desecación de este en poco tiempo, convirtiéndolo en un recurso efímero (Rangel-Acosta et al., 2016b; Rangel-Acosta & Martínez-Hernández, 2017).
Por otro lado, se evidenció que no existe diferencia en la riqueza, abundancia y biomasa de escarabajos coprófagos entre horas de operación de las trampas de caída. Esto podría ser atribuido a los cambios en las propiedades del cebo durante el tiempo de operación de la trampa. Por ejemplo, algunos autores como Barraza (2015) y Rangel-Acosta y Martínez-Hernández (2017), afirman que durante la época seca en los fragmentos de BST en el Caribe colombiano, posterior a las 24 h de operación de las trampas los cebos pueden perder entre 50-70 % de humedad, provocando una disminución en la emisión de olor y consecuentemente en la capacidad de atracción de escarabajos. Un resultado similar fue reportado por Flechtmann et al. (2009) en un bosque tropical semideciduo latifoliado en Selvíria, Mato Grosso del Sur, en Brasil. Estos autores manifiestan que la mayor efectividad del cebo se presenta durante las primeras 24 h, y posteriormente su capacidad de atracción disminuye debido a la pérdida de humedad del mismo. Nuestros resultados concuerdan con las afirmaciones de estos autores, ya que se demostró una pérdida de peso significativa de los cebos y un efecto de la intensidad lumínica sobre este; lo cual influye en la calidad del atrayente y la disminución de su efectividad en la captura de escarabajos en este fragmento de BST.
La cobertura del muestreo tanto a las 24 h como a las 48 h de operación de las trampas de este estudio fue cercana al 100 %. Sin embargo, este valor no difiere a lo reportado en estudios previos realizados en fragmentos de bosque seco en el Caribe colombiano, donde la cobertura del muestreo se encuentra entre el 90-100 %, con solo 24 h de operación de la trampa de caída (Barraza et al., 2010; Navarro et al., 2011; Rangel-Acosta et al., 2016a; Rangel-Acosta et al., 2016b; Rangel-Acosta et al., 2018; Rangel-Acosta et al., 2020; Solís et al., 2011). Lo anterior deja en evidencia que 24 h de operación de las trampas de caída es suficiente para obtener una buena estimación de la riqueza, abundancia y biomasa de escarabajos coprófagos en BST del Caribe colombiano. Sin embargo, prolongar ese tiempo de operación hasta las 48 h eventualmente permitiría capturar algunas especies de escarabajos que son difíciles de detectar durante las primeras horas de operación de las trampas y además de proporcionar un leve aumento de la abundancia y biomasa (Ferrer-Paris et al., 2013; Price & Feer, 2012). No obstante, para la adecuada elección del tiempo de operación de las trampas en campo se debe tener en cuenta el objetivo del trabajo, así como el tipo de hábitat a muestrear. Esto último atendiendo que hábitats simplificados, como los de uso antrópico, requieren menor tiempo de operación de las trampas, mientras que hábitats complejos, como los fragmentos de bosque, requieren de un mayor tiempo (Ferrer-Paris et al., 2013).
Los resultados de este trabajo evidencian que el uso de cebos de mayor tamaño garantiza una mayor efectividad en la estimación de la riqueza, abundancia, diversidad y biomasa de escarabajos. Estos hallazgos apoyan la idea propuesta por Peck y Howden (1984) y Davis, (2000), la cual indica que al usar un cebo de mayor tamaño se garantiza una mejor representatividad del ensamblaje y estimación de las abundancias de ciertas especies. Esta última parte quedó evidenciada en este trabajo, donde se observó que especies como C. juvencus, C. subhyalinus, D. guildingui, E. caribaeus y O. acuminatus, presentaron bajas abundancias en el cebo pequeño, a tal punto que podrían considerarse como especies raras, turistas o de poblaciones bajas. Sin embargo, en el cebo de mayor tamaño estas especies presentaron abundancias altas, de manera que pueden ser catalogadas como especies comunes. A la luz de estos resultados un uso adecuado del tamaño del cebo puede conducir a mejores conclusiones sobre el ensamblaje de escarabajo coprófago y a decisiones más acertada con relación a planes de conservación y manejo de este grupo de insectos.
Por otra parte, se reafirma la idea de que un cebo de mayor tamaño garantiza una mejor representación de todos los grupos de escarabajos (grandes y pequeños, rodadores, cavadores y residentes), sobre todo de los escarabajos grandes (> 10 mm); los cuales están poco representados en las muestras capturadas con cebos pequeños (Errouissi et al., 2004; Finn & Giller, 2000; Finn & Giller, 2002; Peck & Howden, 1984). Lo anterior, se debe a que los cebos de mayor tamaño son más fáciles de detectar por los escarabajos, ya que: 1) emiten una mayor cantidad de olor (Peck & Howden, 1984), 2) mantienen una mayor humedad (Flechtmann et al., 2009) y 3) su deshidratación y la formación de costras ocurren de manera más lenta, garantizando la emisión de olores durante un mayor tiempo (Fincher et al., 1970; Finn & Giller, 2000; Ueda et al., 2015). Estas tres características convierten al cebo grande en una herramienta útil para la realización de muestreos que permitan obtener una buena estimación de la riqueza, abundancia y biomasa de escarabajos coprófagos.
Por otro lado, en ecosistemas naturales la presencia de posturas de estiércol de gran tamaño garantiza una fuente de recurso para el ensamblaje de los escarabajos coprófagos de la subfamilia Scarabaeinae; beneficiando principalmente a los escarabajos de cuerpo grande, ya que estos requieren de mayor cantidad de recurso para su nidificación y alimentación (Peck & Howden, 1984). De igual forma, una mayor porción de estiércol provee un lugar estable para el desarrollo y permanencia de larvas de escarabajos, así como para los adultos que viven dentro de estos o en la interfaz suelo-estiércol (Finn & Giller, 2000; Finn & Giller, 2002). Esto explicaría la alta abundancia y biomasa del grupo funcional de los residentes en los cebos grandes en este trabajo.
La presente investigación resalta la importancia de estandarizar los métodos de capturas de escarabajos coprófagos para investigaciones comparativas, principalmente en ecosistemas con ambientes fuertemente estacionales como el BST en el Caribe colombiano. En este ecosistema las condiciones ambientales durante la época seca (altos valores de temperatura, intensidad lumínica, y baja humedad) disminuyen la efectividad de captura de las trampas; debido a que aceleran la desecación y degradación del atrayente (Rangel-Acosta & Martínez-Hernández, 2017). Teniendo en cuenta lo anterior, el presente trabajo evidencia que la utilización de cebos de mayor tamaño (mediano y grande) durante los periodos secos asegura una mejor estimación de la riqueza, abundancia y biomasa, tanto de escarabajos grandes como pequeños, lo cual permitirá, a posteriori, realizar una buena estimación del ensamblaje (Davis, 2000).
Sin embargo, uno de los principales problemas de emplear cebos de gran tamaño es la dificultad de obtener la cantidad de estiércol necesario para cebar las trampas, ya que conseguir excremento fresco de mamíferos nativos como primates en el BST no es una tarea fácil, debido a los bajos tamaños poblaciones de estos animales y a la degradación acelerada posterior a su postura (observación personal). Además, una alternativa sería el uso de estiércol de mamíferos introducidos, pero éste solo atrae una parte de la fauna de escarabajos (Rangel-Acosta et al., 2012).
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

