











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Annona es un género con 160 especies, 156 se conocen para el Neotrópico (Maas et al., 2019), de ellas, 18 se han registrado paraMéxico (De la Cruz, Castro-Montero, González-Esquinca, 2016), algunas descritas recientemente (Jiménez-Ramírez & Soto-Núñez, 2015). La importancia de las anonas por sus frutos comestibles se tiene ampliamente documentada (Standley, 1920; Luna-Cázares & González-Esquinca, 2015), por lo que algunas especies se cultivan para atender la demanda de frutos en los mercados nacionales e internacionales y otras se recolectan de poblaciones silvestres, entre ellas A. purpurea (Andres-Agustín, 2015).
Annona purpurea se conoce con más de 60 nombre locales (Luna-Cázares & González-Esquinca, 2015), estando entre los más generalizados “cabeza de negro”, “chinguya” y “sincuya”, haciendo alusión al fruto, el cual es cosechado y vendido en los mercados locales (Standley, 1920, Luna-Cázares, & González-Esquinca, 2015; Topete-Corona, 2018), por lo que ha sido considerada una especie de interés frutícola (Standley, 1920; Andres-Agustín, 2015). Se trata de una especie Mesoamérica, con una distribución desde Nayarit y Veracruz, en México, hasta Venezuela, Colombia y Ecuador (Topete-Corona, 2018), en bosques tropicales perennifolios a caducifolios y áreas abiertas dentro de ellos (Luna-Cázares & González-Esquinca, 2015) y en bosques de encino (Topete-Corona, 2018).
Los pobladores mesoamericanos han utilizado las hojas, tallos, raíces, frutos y semillas de la especie, con fines medicinales, como insecticida y antibacterial (Standley, 1920; Luna-Cázares & González-Esquinca, 2008). Los estudios de las propiedades químicas de estas estructuras, revelan la presencia de sustancias con potencial como anticancerígenas, hipoglucémicas, antibacteriales y para la producción de insecticidas y biocombustibles (Luna-Cázares & González-Esquinca, 2008; Vidal-Lezama, Villegas-Monter, Vaquera-Huerta, Robledo-Paz, & Martínez-Palacios, 2019).
Se considera que en algunas regiones de México las poblaciones de la especie están disminuyendo, debido principalmente a la alteración de su hábitat (Vidal-Lezama et al., 2019). A pesar de lo anterior no se conocen investigaciones sobre la estructura de las poblaciones y el hábitat de A. purpurea, por lo que el objetivo de la investigación es describir la estructura poblacional, el hábitat y los factores ambientales que influyen sobre la distribución y abundancia de la especie en el occidente de México.
Materiales y métodos
Área de estudio: La investigación se realizó en la zona tropical de los estados de Jalisco y Colima en el occidente de México (104°03’-104°54’ N & 19°20’-19°46’ W), a una elevación de 300-800 m, con una temperatura media anual de 22.8-25.1 °Cy una precipitación anual de 1 346-1 480 mm; con un periodo de sequía de 6-8 meses y una concentración del 90 % o más de la precipitación de junio a septiembre (Fernández, Zavala, & Romero-Fernández, 2010). La vegetación que se registra en el área corresponde con bosque tropical caducifolio y subcaducifolio y, encinares caducifolios (Rzedowski & McVaugh, 1966).
Registro de datos en campo: De abril a noviembre 2015, se establecieron 24 unidades circulares de 500 m2, a través de un muestreo selectivo de áreas con presencia de A. purpurea. Dentro de cada unidad de muestreo se identificaron, midieron y registraron las especies leñosas con diámetro a la altura de pecho (dap) ≥ 2.5 cm. Dentro de cada parcela (500 m2) se establecieron cinco subparcelas circulares de 5 m2 cada una, una al centro y las otras cuatro hacia los extremos de la parcela, en orientación norte-sur y este-oeste, con la finalidad de evaluar la repoblación de A. purpurea, registrando y midiendo los individuos con un dap < 2.5 cm. Se recolectaron ejemplares de las especies para corroborar las identificaciones y se depositaron en el Herbario ZEA de la Universidad de Guadalajara, como material de referencia de la investigación. Para cada parcela se registró coordenadas, elevación, tipo de vegetación, pendiente, exposición, factores de perturbación, entre otros (Tabla 1). Para obtener por localidad la temperatura y la precipitación, se utilizaron datos disponibles en el Atlas Climático Digital de México (Fernández et al., 2010).
TABLA 1 TABLE 1 Información geográfica y ambiental de las unidades de muestreo de A. purpurea en el occidente de México Geographic and environmental information of the sampling units of A. purpurea in Western Mexico
| Par | Lat | Lon | Ele | Pen | Exp | Ped | Phs | Inc | Pas | Amu | Toc | Tma | Pan |
| P01 | 19°36ˈ28” | 104°18ˈ33” | 650 | 14 | 20° NO | 4 | 20 | 0 | 4 | 0 | 2 | 24.0 | 1 308 |
| P02 | 19°36ˈ26” | 104°18ˈ31” | 650 | 11 | 30° NO | 4 | 22 | 0 | 4 | 5 | 4 | 24.0 | 1 308 |
| P03 | 19°36ˈ41” | 104°18’38” | 550 | 17 | 40° NO | 1 | 35 | 0 | 3 | 1 | 2 | 24.0 | 1 304 |
| P04 | 19°40’18” | 104°25’46.4” | 662 | 44 | 20° NE | 4 | 30 | 4 | 3 | 2 | 4 | 24.1 | 1 281 |
| P05 | 19°40’17.7” | 104°25’57.7 | 630 | 31 | 35° NO | 1 | 20 | 1 | 2 | 0 | 7 | 24.1 | 1 281 |
| P06 | 19°27’15.9” | 104°29’04” | 603 | 57 | 88° NO | 2 | 25 | 4 | 2 | 1 | 0 | 24.2 | 1 540 |
| P07 | 19°27’15.2” | 104°29’8.5” | 565 | 10 | 40° SE | 3 | 30 | 1 | 3 | 1 | 2 | 24.2 | 1 540 |
| P08 | 19°31’12.9” | 104°26’7.7” | 485 | 5 | 83° NE | 1 | 10 | 4 | 4 | 0 | 0 | 25.0 | 1 578 |
| P09 | 19°23’39.9” | 104°27’37.6” | 639 | 50 | 03° SE | 1 | 30 | 4 | 4 | 0 | 1 | 23.9 | 1 509 |
| P10 | 19°23’40” | 104°27’34.7” | 672 | 28 | 32° SE | 2 | 10 | 4 | 4 | 0 | 0 | 23.9 | 1 509 |
| P11 | 19°38’29.7” | 104°54’11.8” | 354 | 12 | 65° NE | 1 | 10 | 4 | 4 | 1 | 2 | 25.1 | 1 244 |
| P12 | 19°35’25.8” | 104°38’31” | 439 | 7 | 14° SE | 2 | 10 | 4 | 4 | 0 | 0 | 24.7 | 1 475 |
| P13 | 19°34’34.7” | 104°92’10.3” | 561 | 32 | 58° SE | 4 | 7 | 4 | 4 | 0 | 0 | 24.2 | 1 470 |
| P14 | 19°34’51.1” | 104°42’11.1” | 565 | 30 | 86° SE | 2 | 10 | 4 | 3 | 2 | 0 | 25.0 | 1 278 |
| P15 | 19°34’07.3” | 104°42’04.5” | 579 | 18 | 62° SE | 1 | 10 | 4 | 4 | 0 | 4 | 25.1 | 1 238 |
| P16 | 19°20’22” | 104°03’58.4” | 697 | 52 | 15° SO | 2 | 15 | 1 | 3 | 1 | 0 | 22.8 | 1 331 |
| P17 | 19°20’0.94” | 104°04’08.8” | 715 | 22 | 45° SO | 2 | 5 | 1 | 3 | 10 | 3 | 23.3 | 1 465 |
| P18 | 19°21’25.1” | 104°05’46” | 632 | 48 | 52° SE | 2 | 10 | 1 | 3 | 4 | 2 | 23.5 | 1 291 |
| P19 | 19°21’33.3” | 104°03’48.9” | 726 | 40 | 80° SE | 2 | 7 | 4 | 4 | 0 | 0 | 23.5 | 1 478 |
| P20 | 19°44’47.4” | 104°31’25.9” | 540 | 40 | 55° NO | 2 | 7 | 4 | 4 | 0 | 18 | 24.8 | 1 544 |
| P21 | 19°45’14.1” | 104°31’14.3” | 546 | 5 | 65° NE | 1 | 40 | 1 | 4 | 0 | 2 | 24.7 | 1 503 |
| P22 | 19°45’19.3” | 104°31’13.6” | 568 | 8 | 18° SE | 1 | 15 | 1 | 4 | 0 | 11 | 24.7 | 1 503 |
| P23 | 19°45’1.7” | 104°31’4.9” | 579 | 30 | 65 SE | 4 | 10 | 1 | 4 | 0 | 2 | 24.7 | 1 503 |
| P24 | 19°42’34.4” | 104° 32’37” | 429 | 16 | 60° SE | 1 | 20 | 4 | 4 | 0 | 2 | 25.1 | 1 603 |
Par = parcela, Lat = latitud, Lon = longitud, Ele = elevación en m s.n.m., Pen = Pendiente (%), Exp = exposición, Ped = pedregosidad, Phs = profundidad del horizonte superficial (cm), Inc = incendios, Pas = pastoreo, Amu = árboles muertos, Toc = tocones, Tma = temperatura media anual (°C), Pan = precipitación anual (mm).
Par = plot, Lat = latitude, Lon = longitude, Ele = elevation in m.a.s.l., Pen = slope (%), Exp = exposition, Ped = stoniness, Phs = depth superficial horizon (cm), Inc = fires, Pas = livestock, Amu = dead trees, Toc = stumps, Tma = average annual temperature (°C), Pan = annual precipitation (mm).
Análisis de los datos: Estructura poblacional de A. purpurea: se definió en función de los tallos con dap ≥ 2.5 cm registrados en las 24 parcelas. Las clases diamétricas se realizaron en función de:M = 3 log10N, donde M = número de clases diamétricas y N = número de tallos de A. purpurea. El intervalo de clase fue igual a (diámetro mayor-diámetro menor) / M. Se determinó por parcela la densidad, área basal (AB) y el índice devalor de importancia (IVI) de A. purpurea, este último como la suma de los promedios relativos de densidad y área basal (Curtis & McIntosh, 1951; Vázquez & Givnish, 1998; Cuevas, García-Moya, Vázquez-García, &Núñez-López, 2008).
El análisis de la repoblación se realizó considerando los individuos de A. purpurea con dap < 2.5 cm registrados en las 24 parcelas, realizando las sumas de los individuos registrados en las subparcelas de cada parcela. Se realizaron correlaciones entre los atributos estructurales de A. purpurea y variables ambientales en busca de asociación entre los parámetros. Para las variables cuantitativas se aplicó la correlación de Pearson, para las cualitativas (Ped, Pas e Inc) la de Spearman (Balzarini et al., 2008). Las variables ambientales incluidas fueron la elevación (Ele), registrada en el centro de cada parcela con un GPS Garmin Etrex, la pendiente (Pen), registrada con una pistola Haga, la exposición (Exp), tomada con una brújula Brunton, la pedregosidad (Ped), incidencia de incendios (Inc) y pastoreo (Pas), con base en los criterios establecidos en Olvera, Moreno y Figueroa (1996), la profundidad del horizonte superficial (Phs), registrada en cada subparcela de 5 m2 y promediada para toda la parcela, se midió en cm con una regla graduada, tocones (Toc), se contabilizaron al interior de la parcela de 500 m2, considerando únicamente aquellos ≥ 5 cm de diámetro a los 30 cm de altura, con señales claras de haber sido cortados por el hombre, árboles muertos (Amu), se contabilizaron al interior de la parcela, incluyendo aquellos en pie y caídos que no habían iniciado su descomposición y con un dap ≥ 10 cm. Los análisis de los datos se realizaron utilizando el software InfoStat (Di Rienzo, Casanoves, Balzarini, Gonzalez, Tablada, & Robledo 2019), PC-ORD v7 (McCune & Mefford, 2011) y Primer v7 (Clarke & Gorley, 2015).
Especies asociadas con A. purpurea y estructura de la comunidad: Para el análisis de asociación de especies, y determinar con cuáles A. purpurea tiene sus mayores relaciones y con cuáles se agrupa, se utilizó la matriz de IVI, considerando sólo las especies que presentaron un índice ≥ 10 % en una o más parcelas. Las especies fueron estandarizadas dividiendo el valor de cada una de ellas por su total y multiplicándolo por 100, seguido por un cálculo de la semejanza Bray-Curtis entre pares de especies, lo que se conoce como índice de asociación de especies de Whittaker(Clarke, Gorley, Somerfield, & Warwick, 2014). Con la matriz de semejanza de las especies se realizó un análisis de agrupamiento, con grupo promedio como método de unión de grupos, acompañado de una prueba de perfiles de semejanza (SIMPROF) tipo 2 y 3. La prueba tipo 2 trabaja con la hipótesis nula de la no existencia de asociaciones entre las especies y la tipo 3 con la no existencia de diferencias entre los grupos de especies, teniendo como prueba estadística a π (pi), la cual es una prueba de permutaciones, para la cual se puede encontrar más información en Clarke et al. (2014). Los resultados se expresan a través de un dendrograma, en el cual los grupos estadísticamente disimiles se diferencian por una línea continua.
Se determinóla riqueza de especies, densidades, AB e IVI para las especies por parcela (Curtis & McIntosh, 1951;Vázquez-García & Givnish, 1998; Cuevas et al., 2008). Se integró una matriz de IVI con las 23 que contribuyeron con al menos el 10 % en una o más parcelas. La matriz de variables ambientales incluyó la Ele, Pen, Exp, Ped, Phs, Inc, Pas, Amu, Toc, Tma, Pan (Tabla 1). Las variables cualitativas como la Ped, Inc y Pas, fueron convertidas en variables auxiliares (Dummy) y se trataron como cuantitativas. La variable Exp fue transformada siguiendo el criterio de Beers, Dress y Wensel (1966). Quitando las variables con autocorrelaciónes significativas (McCune & Mefford, 2011) e incluyendo las variables cualitativas transformadas a dummy, la matriz se integró por 11 variables ambientales. Utilizando las dos matrices se hizo un Análisis de Correspondencia Canónica (ACC) seleccionando las opciones: los resultados de los ejes centrados y estandarizados a una unidad de varianza, los ejes escalados para representar la optimización de las especies, los resultados finales de la ordenación en el gráfico representando combinaciones lineales de las variables ambientales, y la hipótesis nula de la no existencia de relación entre la matriz de las especies y la de variables ambientales, con 999 iteraciones (McCune & Mefford, 2011). La prueba estadística para aceptar o rechazar la hipótesis nula se realiza a través de una prueba de permutaciones (prueba de Monte Carlo), la cual muestra como los valores propios observados (eigenvalores) y, las correlaciones entre las especies y el ambiente comparan con los de las corridas aleatorias y determinando un valor de “p” (McCune & Grace, 2002).
Resultados
Estructura poblacional de A. purpurea y su relación con variables ambientales: Se registraron 402 individuos de A. purpurea con 1 108 tallos con un diámetro ≥ 2.5 cm en una superficie de 1.2 ha. La altura promedio de los individuos fue de 9.34 ± 3.64 m, registrándose 34 individuos con alturas entre 15-18 m. El 76 % de los individuos presentó entre uno y tres tallos y sólo el 24 % tuvomás de tres, encontrándose 24 individuos con 10-15 tallos. El diámetro promedio de los tallos fue de 9.11 ± 5.96 cm. La densidad de tallos de A. purpurea por parcela osciló entre 13-105 con una media de 46.17 ± 7.27, mientras que el AB fue de 0.077-0.72 m2 con una media de 0.0093 ± 0.014 y los IVI estuvieron entre 15.67-83.43 % con una media de 48.31 ± 21.54 por parcela (Tabla 2). La densidad de A. purpurea representó el 49 % (1 108 tallos 1.2 ha-1) y el AB el 38.31 % (10.30 m2 1.2 ha-1) de la comunidad.
TABLA 2 TABLE 2 Atributos estructurales de A. purpurea por unidad de muestreo y atributos estructurales de la comunidad Structural attributes of A. purpurea by sampling unit and structural attributes of the community
| Par | Den AP | Tal AP | Aba AP | IVI AP | Reg AP | Fam | Gén | Spp | Aba Com | Tal Com |
| P01 | 16 | 18 | 0.1546 | 22.53 | 0 | 13 | 15 | 16 | 1.58 | 51 |
| P02 | 16 | 16 | 0.0769 | 13.38 | 0 | 14 | 15 | 17 | 1.82 | 71 |
| P03 | 22 | 27 | 0.2585 | 23.66 | 0 | 16 | 19 | 22 | 1.17 | 107 |
| P04 | 40 | 105 | 0.7100 | 77.49 | 6 | 7 | 7 | 7 | 1.08 | 118 |
| P05 | 14 | 50 | 0.1771 | 52.05 | 1 | 6 | 6 | 6 | 0.80 | 61 |
| P06 | 8 | 39 | 0.3883 | 50.2 | 1 | 10 | 13 | 13 | 0.63 | 101 |
| P07 | 15 | 77 | 0.5595 | 63.24 | 1 | 12 | 14 | 15 | 0.80 | 136 |
| P08 | 16 | 53 | 0.4020 | 55.87 | 0 | 7 | 7 | 7 | 0.76 | 90 |
| P09 | 20 | 47 | 0.6924 | 56.31 | 1 | 12 | 16 | 17 | 0.99 | 110 |
| P10 | 10 | 28 | 0.3817 | 43.01 | 2 | 4 | 5 | 5 | 1.23 | 51 |
| P11 | 8 | 27 | 0.4220 | 65.7 | 1 | 6 | 7 | 7 | 0.60 | 44 |
| P12 | 11 | 37 | 0.6032 | 39.3 | 5 | 11 | 14 | 15 | 3.19 | 62 |
| P13 | 11 | 24 | 0.3250 | 29.31 | 1 | 15 | 22 | 24 | 0.97 | 95 |
| P14 | 3 | 13 | 0.2903 | 26.09 | 3 | 14 | 19 | 21 | 0.77 | 89 |
| P15 | 15 | 66 | 0.7179 | 72.56 | 2 | 7 | 8 | 8 | 1.05 | 86 |
| P16 | 14 | 42 | 0.1108 | 24.48 | 0 | 10 | 12 | 17 | 0.57 | 142 |
| P17 | 11 | 80 | 0.3460 | 33.78 | 0 | 16 | 20 | 23 | 1.97 | 160 |
| P18 | 9 | 33 | 0.3108 | 22.74 | 0 | 22 | 23 | 26 | 1.31 | 151 |
| P19 | 13 | 38 | 0.7084 | 56.85 | 3 | 10 | 12 | 12 | 0.97 | 93 |
| P20 | 53 | 79 | 0.3764 | 61.82 | 2 | 8 | 10 | 11 | 0.77 | 106 |
| P21 | 8 | 18 | 0.3504 | 52.45 | 0 | 6 | 7 | 8 | 0.86 | 28 |
| P22 | 24 | 103 | 0.7411 | 92.16 | 0 | 5 | 5 | 6 | 0.77 | 117 |
| P23 | 37 | 68 | 0.4720 | 38.89 | 0 | 10 | 14 | 15 | 1.27 | 168 |
| P24 | 8 | 20 | 0.7294 | 85.67 | 0 | 2 | 2 | 2 | 0.96 | 21 |
Par = parcela, Den AP = densidad de individuos de A. purpurea, Tal AP = densidad de tallos de A. purpurea, Aba AP = área basal de A. purpurea (m2), IVI AP = índice de valor de importancia de A. purpurea (%), Reg AP = regeneración de A. purpurea, Fam = familias, Gén = géneros, Spp = especies, Aba Com = área basal de la comunidad (m2), Tal Com = densidad de tallos en la comunidad.
Par = plot, Den AP = density of A. purpurea individuals, Tal AP = stem density of A. purpurea, Aba AP = basal area of A. purpurea (m2), IVI AP =importance value index of A. purpurea (%), Reg AP = regeneration of A. purpurea, Fam = families, Gen = genera, Spp = species, Aba Com = basal area of the community (m2), Tal Com = stem density in the community.
La estructura diamétrica de A. purpurea presentó una forma de “J” invertida, con una mayor concentración de tallos en las categorías diamétricas inferiores y una disminución hacia las superiores, con el 85 % de los tallos en las tres primeras categorías (Fig. 1). La densidad de tallos mostró una correlación positiva y significativa con la presencia de Toc (r = 0.53, P = 0.01), el AB y el IVI con la incidencia de Inc (r = 0.55 y r = 0.50, P = 0.005 y P = 0.01, respectivamente) y el IVI con la Tma (r = 0.5 y P = 0.01) y negativa con la Ped (r = -0.48, P = 0.029). La regeneración fue escasa, en 11 parcelas no se registraron individuos y solo en una se encontraron seis. Una correlación positiva y significativa se obtuvo entre la incidencia de Inc y la regeneración (r = 0.70, P = 0.001).
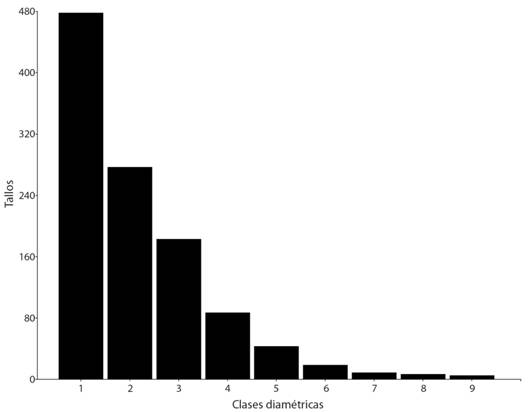
Fig. 1. Fig. 1 Estructura diamétrica de A. purpurea. La clase 1 = 2.50-6.52, 2 = 6.52-10.54, 3 = 10.54-14.57, 4 = 14.57-18.59, 5 = 18.59-22.61, 6 = 22.61-26.63, 7 = 26.63-30.66, 8 = 30.66-34.68, 9 = 34.68-38.70. Diametric structure of A. purpurea. Class 1 = 2.50-6.52, 2 = 6.52-10.54, 3 = 10.54-14.57, 4 = 14.57-18.59, 5 = 18.59-22.61, 6 = 22.61-26.63, 7 = 26.63-30.66, 8 = 30.66-34.68, 9 = 34.68-38.70.
Estructura de la comunidad y hábitat de A. purpurea: Se registraron 47 familias, 87 géneros y 109 especies de plantas leñosas con un dap ≥ 2.5 cm en 1.2 ha. La riqueza de familias osciló de 2-22 por parcela, la de géneros de 2-23, la de especies de 2-26 (Tabla 2). Las familias con mayor riqueza de especies fueron: Fabaceae (17), Euphorbiaceae (8) y Moraceae, Rubiaceae y Salicaceae (6 cada una). Los géneros con más especies fueron Ficus (4), Casearia, Randia y Vachellia (3 cada uno). El AB por parcela osciló de 0.57-3.19 m2 y la densidad de 21-168 tallos (Tabla 2). Los mayores IVI promedio los tuvieron A. purpurea (48.31 %), Tabebuia rosea (3.96 %), Quercus magnoliifolia (3.79 %), Enterolobium cyclocarpum (3.42 %), Lonchocarpus salvadorensis (2.97 %), Casearia arguta (2.72 %) y Guazuma ulmifolia (2.67 %). La prueba SIMPROF, mostró seis grupos de especies estadísticamente diferentes (Fig. 2). Annona purpurea tuvo sus mayores índices de asociación con G. ulmifolia (48.58 %), T. rosea (34.63 %), C. arguta (34.41 %), E. cyclocarpum (29.17 %) y Q. magnoliifolia (29.13 %), formando con G. ulmifolia un grupo estadísticamente diferente a los demás (Fig. 2). Ambas especies compartieron el 58 % de las unidades de muestreo con IVI altos.
El ACC obtuvo una inercia total de 7.57, con una varianza extraída en los tres primeros ejes de 28.1 %. El eje 1 extrajo el 10.7 %, el 2 el 9.4 % y 3 el 8.1 %. Los coeficientes canónicos estandarizados con mayor contribución al eje 1 fueron los Inc_1 e Inc_4 y el Pas_3; para el eje 2 la Ele, Inc_4 y la Ped_2 y para el 3, Ped_2, Ped_4 y el Pas_4 (Tabla 3). La prueba de Monte Carlo para los valores propios de los ejes obtuvo una p = 0.002 y para la correlación entre la matriz de las especies y de las variables ambientales una p = 0.005. De acuerdo con el diagrama de ordenación, A.purpurea y las especies con las que tiene mayor relación, presentaron sus mayores IVI en los sitios con incendios severos (Inc_4), más precipitación anual (Pan), menor elevación y con menos pedregosidad (Fig. 3). En general fueron también estas variables las que presentaron sus mayores correlaciones con los ejes (en el sentido de ter Braak, 1986) (Tabla 3).
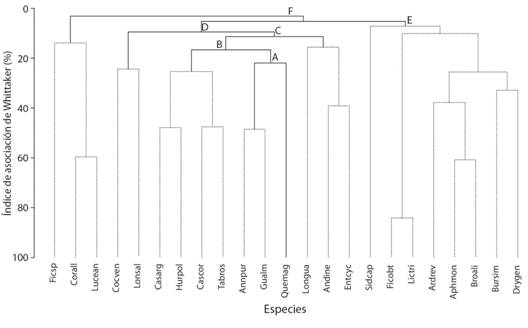
Fig. 2 Fig. 2 Dendrograma de las 21 especies más importantes por su IVI. Ficsp = Ficus sp., Corall = Cordia alliodora, Luecan = Luehea candida, Cocven = Coccoloba venosa, Lonsal = Lonchocarpus salvadorensis, Casarg = Casearia arguta, Hurpol = Hura polyandra, Cascor = Casearia corymbosa, Tabros = Tabebuia rosea, Annpur = Annona purpurea, Guaulm = Guazuma ulmifolia, Quemag = Quercus magnoliifolia, Longua = Lonchocarpus guatemalensis, Andine = Andira inermis, Entcyc = Enterolobium cyclocarpum, Sidcap = Sideroxylon capiri, Ficobt = Ficus obtusifolia, Lictri = Licaria triandra, Ardrev = Ardisia revoluta, Aphmon = Aphananthe monoica, Broali = Brosimum alicastrum, Bursim = Bursera simaruba, Drygen = Drypetes gentryi. Líneas continuas indican grupos de especies estadísticamente diferentes. Las letras indican las diferencias estadísticas entre los grupos: A π = 7.37 p = 0.02, B π = 5.36 p = 0.002, C π = 4.24 p = 0.001, D π = 3.27 p = 0.006, E π = 2.38 p = 0.003, F π = 2.2 p = 0.003. Dendrogram of the 21 most important species for their IVI. Ficsp = Ficus sp., Corall = Cordia alliodora, Luecan = Luehea candida, Cocven = Coccoloba venosa, Lonsal = Lonchocarpus salvadorensis, Casarg = Casearia arguta, Hurpol = Hura polyandra, Cascor = Casearia corymbosa, Tabros = Tabebuia rosea, Annpur = Annona purpurea, Guaulm = Guazuma ulmifolia, Quemag = Quercus magnoliifolia, Longua = Lonchocarpus guatemalensis, Andine = Andira inermis, Entcyc = Enterolobium cyclocarpum, Sidcap = Sideroxylon capiri, Ficobt = Ficus obtusifolia, Lictri = Licaria triandra, Ardrev = Ardisia revoluta, Aphmon = Aphananthe monoica, Broali = Brosimum alicastrum, Bursim = Bursera simaruba, Drygen = Drypetes gentryi. Solid lines indicate statistically different groups of species. Letters indicate statistical differences between groups: A π = 7.37 p = 0.02, B π = 5.36 p = 0.002, C π = 4.24 p = 0.001, D π = 3.27 p = 0.006, E π = 2.38 p = 0.003, F π = 2.2 p = 0.003.
TABLA 3 TABLE 3 Coeficientes canónicos obtenidos a través de la regresión múltiple con CCA, correlaciones de las variables ambientales con los ejes y correlaciones de A. purpurea y otras especies con los ejes Canonical coefficients getted through multiple regression with CCA, correlations of the environmental variables with the axes and correlations of A. purpurea and other species with the axes
| Variable | Coeficientes estandarizados | Correlaciones con los ejes | ||||
| Eje 1 | Eje 2 | Eje 3 | Eje 1 | Eje 2 | Eje 3 | |
| Elevación | 0.332 | 1.145 | 0.029 | 0.419 | 0.763 | -0.311 |
| Profundidad de horizonte superficial | -0.188 | -0.016 | -0.241 | 0.537 | -0.164 | 0.119 |
| Tocones | 0.048 | 0.080 | 0.017 | 0.113 | 0.043 | -0.085 |
| Precipitación anual | 0.178 | 0.321 | 0.382 | -0.414 | 0.543 | 0.056 |
| Pedregosidad (Ped_2). | -0.307 | -0.595 | -0.738 | -0.278 | 0.288 | 0.173 |
| Pedregosidad (Ped_3). | -0.115 | 0.061 | -0.110 | -0.073 | 0.136 | 0.083 |
| Pedregosidad (Ped_4). | -0.130 | -0.281 | -0.704 | 0.241 | -0.038 | -0.773 |
| Pastoreo fuerte (Pas_3). | 0.427 | -0.228 | 0.250 | 0.493 | -0.063 | 0.461 |
| Pastoreo severo (Pas_4). | -0.137 | -0.257 | -0.627 | -0.424 | -0.040 | -0.520 |
| Incendios ligeros (Inc_1). | -0.948 | 0.211 | -0.368 | -0.098 | 0.242 | -0.163 |
| Incendios severos (Inc_4). | -0.928 | 0.638 | 0.300 | -0.653 | 0.035 | 0.366 |
| Annona purpurea | -0.451 | 0.449 | 0.363 | |||
| Casearia arguta | -0.330 | -0.274 | 0.298 | |||
| Enterolobium cyclocarpum | -0.703 | 0.676 | -0.262 | |||
| Guazuma ulmifolia | -0.418 | 0.444 | 1.00 | |||
| Quercus magnoliifolia | -0.596 | 0.637 | 0.470 | |||
| Tabebuia rosea | -0.239 | 0.233 | -0.104 | |||
Ped_2 = 20-40 % superficie de la parcela cubierta de piedras, Ped_3 = 40-60 % superficie con piedras, Ped_4 = 60-80 % superficie con piedras.
Ped_2 = 20-40 % area of the plot covered with stones, Ped_3 = 40-60 % area with stones, Ped_4 = 60-80 % area with stones.
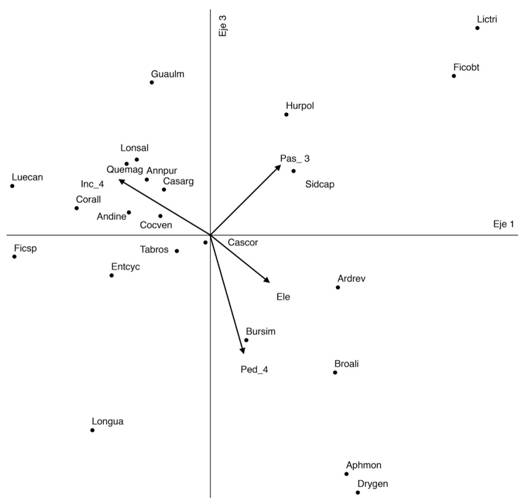
Fig. 3 Fig. 3 Gráfico de CCA para los ejes 1 y 3. Los puntos indican las especies, para sus nombres ver pie de la Fig. 2. Los vectores representan variables ambientales, Inc_4 = incendios severos, Pas_3 = pastoreo fuerte, Ele = elevación, Ped_4 = parcelas con el 60-80 % de su superficie cubierta por piedras. CCA graph for axes 1 and 3. The dots indicate the species, for their names see figure 2. Vectors represent environmental variables, Inc_4 = severe fires, Pas_3 = strong grazing, Ele = elevation, Ped-4 = plots with 60-80 % of their area covered by stones.
Discusión
Estructura poblacional de A. purpurea y su relación con variables ambientales: La altura promedio de A. purpurea en el occidente de México, se encuentra dentro de las registradas para la especie; pero aquellas hasta de 18 m, son superiores a las que se mencionan en las descripciones de la especie (Standley, 1920; Standley & Steyermark, 1946; Schatz, 2009). Esto podría interpretarse de que en la región de estudio, existen condiciones ambientales que permiten un mejor desarrollo en la altura de la especie o bien que en otras localidades no se han hecho mediciones más precisas de su altura y se han empleado las que se registran en la literatura. No se encontró información para comparar la ramificación de la especie por debajo de 1.30 m de altura. El que se registren individuos con más de 10 tallos, pudiera atribuirse a la capacidad de generar brotes que tiene la especie, lo cual ya ha sido registrado en otras investigaciones (Griscom, Griscom, & Ashton, 2009). Los porcentajes altos de densidad de tallos, AB e IVI de A. purpurea, es coincidente con lo registrado en otros estudios cercanos al área de estudio (Cuevas, 2002).
La estructura diamétrica de la población en forma de “J” invertida, indica una buena reincorporación de individuos (Bongers, Pompa, Meave del Castillo, & Carabias, 1988), aunque ha sido registrado más para especies umbrófilas (Cuevas, López-Mata, & García-Moya, 2002). Es de considerar que los histogramas pueden diferir en función del criterio que se siga para generar el número y la amplitud de clases diamétricas en la población, (Fox, 2016). Si aplicáramos el criterio de Hubbell y Foster (1987), clases de tamaño que consecuentemente duplican a la que le precede, la población de A. purpurea (gráfico no presentado), se ajusta más a las especiesheliófilas o pioneras consignadas en ese trabajo (Hubbell & Foster, 1987). Las densidades grandes de tallos en las primeras categorías diamétricas, se atribuye a la capacidad que tiene la especie para regenerar en claros grandes, los cuales son comunes y continuos en la región de estudio, lo que podría promover que la especie colonice y se establezca en forma continua en área abiertas (obs. de los autores en campo), a pesar de su escasa o nula regeneración bajo su propio dosel. La relación positiva de la regeneración con la incidencia de Inc, es un elemento más de apoyo de que la especie requiere de espacios abiertos para su establecimiento, donde la semilla pueda llegar por dispersores, principalmente mamíferos medianos, entre los que se han observado en campo al mapache, zorra, coyote, tejón y jabalí, algunos de los cuales ya han sido registrados como dispersores de semillas de Annona (Natural Heritage Trust, 2003; Medrano-Nájera, Ramírez-Pinero, Guevara-Sada, 2014). Otros elementos que dan soporte para considerar a A. purpurea como una especie con preferencia por áreas abiertas, en bosque tropical subcaducifolio, son caracteres que tiene la especie y que han sido considerados de taxones pioneros (Swaine & Whitmore, 1988). Como ejemplo, el tener hojas caducifolias, membranáceas y grandes (Standley, 1920; Standley & Steyermark, 1946; Schatz, 2009; Topete-Corona, 2018), multiplicación de tallos por brotes, escasa repoblación de plántulas y juveniles bajo su dosel (en este trabajo).
Las correlaciones positivas del IVI con la Tma y el AB e IVI con la incidencia de incendios, son elementos que apoyan de que A. purpurea es una especie de zonas tropicales cálidas, donde hay perturbaciones continuas que generan claros grandes (Denslow, 1980; Hubbell et al., 1999). Esto último también soportado por el que la mayor densidad de tallos se encuentra en donde hay más tocones y por consiguiente mayor apertura del dosel. Las aseveraciones de que, en algunas regiones de México, las poblaciones de A. purpurea están disminuyendo debido principalmente a la alteración de su hábitat (Vidal-Lezama et al., 2019), parecen no aplicar para el occidente de México, donde actividades como la apertura de áreas para la ganadería y la incidencia de incendios favorecen a la especie.
Estructura de la comunidad y hábitat de A. purpurea: La riqueza de familias, géneros y especies registradas en 1.2 ha, está por debajo de lo encontrado en superficies iguales en bosques tropicales subcaducifolios cercanos (Vázquez-Magaña & Mendoza-Cortes, 2006), diferencias que podrían atribuirse a que son áreas con diferentes grados de conservación. La riqueza de taxones es superior a los registrado en áreas de la costa del estado de Jalisco (González-Cueva, Aguirre-Calderón, Alanís-Rodríguez, & Corral-Rivas, 2018), diferencias que podrían atribuirse al límite inferior de los diámetros de los tallos que fueron considerados en las investigaciones, dap ≥ 2.5 cm en el primero y dap > 7.5 cm en el segundo. Las familias y géneros registrados con mayor riqueza específica, también coinciden con lo encontrado por Cuevas (2002), en lo que llamó bosque tropical subcaducifolio de partes bajas. Aunque puede haber coincidencia, con otras investigaciones, con algunas de las familias con mayor riqueza en este tipo de vegetación, no ocurre lo mismo con los géneros y las especies con las mayores abundancias, AB e IVI, lo cual se atribuye a que cuando se cambia de una localidad a otra, sobre todo cuando las distancias entre los sitios que se comparan se incrementan, las condiciones ambientales varían y los estados de conservación o sucesión son diferentes (Gutiérrez-Báez, Zamora-Crescencio, & Hernández-Mundo, 2012; Dzib-Castillo, Chanatásig-Vaca, & González-Valdivia, 2014; González-Cuevas et al., 2018).
Las especies con las que A. purpurea comparte los mayores índices de asociación como G. ulmifolia,T. rosea, C. arguta, E. cyclocarpum, han sido registradas como sus compañeras en la región (Cuevas, 2002;Vázquez-Magaña& Mendoza-Cortés, 2006), pero también en Centro y Sudamérica (Oviedo-Pérez, 2015; Cabrera-Amaya & Rivera-Díaz, 2016), sobre todo la asociación de A. purpurea y G. ulmifolia, grupo que fue estadísticamente diferente a los restantes (Fig. 2). Esta asociación podría atribuirse a que las dos especies tienen requerimientos ambientales similares, sobre todo el hecho que se ven favorecidas en áreas de vegetación secundaria derivada de bosques tropicales húmedos, y tienen en común una fuerte capacidad de multiplicación por rebrote (Griscom et al., 2009).
Las variables ambientales recuperadas por el CCA como de más influencia sobre la distribución y abundancia de A. purpurea y de las especies con las que presenta mayores asociaciones (G. ulmifolia, Q. magnoliifolia,E. cyclocarpum), son la Pan, los incendios severos (Inc_4), menor Ele y menos Ped (Fig. 3, Tabla 3). La influencia de la Pan en este grupo de especies, podría ser interpretada como la preferencia de ellas por los lugares más húmedos. Annona purpurea tiene algunas características morfológicas y fisiológicas que parecen responder a esta variable, como el poseer hojas membranáceas y grandes, las cuales generan periodos de crecimiento estacional, aprovechando la humedad durante el periodo corto de lluvias, con abscisión del follaje en la estación seca y más larga del año (Reicht, 2014). Lo anterior permitiría explicar porque los mayores atributos estructurales de A. purpurea se registran en los bosques más húmedos (bosque tropical subcaducifolio), aunque algunas veces se pueden registrar individuos en el bosque tropical caducifolio o en la transición con el bosque de encino. La influencia de la precipitación sobre A. purpurea ya ha sido registrada y se ha encontrado un acoplamiento de la producción de hojas y flores con el inicio de las lluvias y la maduración de los frutos con el final de la temporada (González-Esquinca, De la Cruz, Castro-Moreno, Riley-Saldaña, 2016). El grupo de especies mencionado también presenta una relación positiva y estrecha con Inc_4, lo que podría atribuirse a que se trata de especies que requieren apertura de claros grandes para su establecimiento y que se ven favorecidas en lugares con perturbaciones, incluyendo el fuego (López-Moctezuma, Rodríguez-Trejo, Santiago-Cortes, & Granado-Sánchez 2015; Oviedo-Pérez, 2015; CONABIO, 2020; IFO-CATIE, 2020). Otra variable importante fue la Ele, registrando A. purpurea sus mayores valores estructurales en los sitios con menor elevación, si consideramos que esta variable estuvo correlacionada en forma negativa con la Tma, entonces existe una preferencia de la especie por los lugares más cálidos, lo cual coincide con lo registrado por Cuevas (2002) y Vázquez-Magaña y Mendoza-Cortés (2006) (Fig. 3, Tabla 3).
Concluimos que A. purpurea presenta una estructura poblacional con buena reincorporación de individuos y que su distribución y abundancia parecen responder principalmente a factores ambientales como precipitación anual, temperatura media anual, pedregosidad y a perturbaciones causadas por incendios y ganadería.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

