










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.4 San José Dec. 2012
Fluctuación poblacional de Xyleborus ferrugineus y X. affinis (Coleoptera: Curculionidae) en ecosistemas de Tabasco, México
Population fluctuation of Xyleborus ferrugineus and X. affinis (Coleoptera: Curculionidae) in ecosystems of Tabasco, Mexico
Population fluctuation of Xyleborus ferrugineus and X. affinis (Coleoptera: Curculionidae) in ecosystems of Tabasco, Mexico
*Dirección para correspondecia
Abstract
The Scolytinae insects are commonly known as bark or ambrosial beetles because of their feeding habits. Among them, some species restrict their reproductive activity to a host plant or a small number of them, whereas others are highly polyphagous. In order to evaluate the population dynamics of Scolytinae species, populations of Xyleborus ferrugineus and X. affinis in ecosystems from Tabasco, Mexico were studied. The study was carried out from November 2010-July 2011, February 2010-January 2011 and 2007. The trapping methods used were alcohol traps, light traps and direct capture on their host plants. A total of 688 specimens of X. ferrugineus and 3 911 specimens of X. affinis were collected. The population dynamics of X. ferrugineus showed low size population in the studied ecosystems, without any marked seasonality. The highest population sizes were recorded both in dry season (March-May) and rainy season (September-December). Unlike the populations of X. ferrugineus, those of X. affinis were more abundant and showed the highest peaks during the rainy seasons (except in May at the Botanical Garden “José Narciso Rovisora”). Alcohol and light traps can be complementary methods for monitoring the populations of these types of insects. They occur most of the year and theirs population dynamics depends on food resources availability as well as environmental factors such as temperature and humidity. Judging by the observed abundance it is suggested that X. affinis has been adapted and exploited resources in a better way than X. ferrugineus in those ecosystems. Correlations between abundance and climatic factors had both positive and negative values. The results reflect the behavior of populations. However, a detail assessment of the biological and abiotic factors that influence the fluctuations of these insects is required.
Key words: insects, ambrosial, borers, host, dynamic.
Resumen
Los Scolytinae son insectos comúnmente conocidos como coleópteros descortezadores o ambrosiales debido a su forma de alimentarse. Existen especies que restringen su actividad reproductiva a una planta huésped o a un número reducido de ellas, otras son altamente polífagas. Se estudiaron las poblaciones de Scolytinae con el objetivo de conocer la fluctuación poblacional de X. ferrugineus y X. affinis en ecosistemas de Tabasco, México durante noviembre 2010-julio 2011, febrero 2010-enero 2011 y 2007. Los métodos de captura utilizados fueron trampa de alcohol, trampa de luz y captura directa sobre sus plantas huésped. Se recolectaron 688 especímenes de X. ferrugineus y 3 911 de X. affinis. La fluctuación poblacional de X. ferrugineus mostró de manera general poblaciones bajas en los ecosistemas estudiados sin alguna estacionalidad marcada, registrándose los máximos picos poblacionales tanto en época seca (marzo-mayo) como lluviosa (septiembre-diciembre). A diferencia de las poblaciones de X. affinis que fueron más abundantes y mostraron en la mayoría de los sitios (excepto en el Jardín Botánico José Narciso Rovirosa que se presentó en mayo) picos poblacionales en época lluviosa. Las trampas de alcohol y de luz son métodos de recolecta que pueden ser complementarios para el monitoreo de las poblaciones de estos insectos, los cuales se encuentran presentes durante la mayor parte del año y su fluctuación poblacional es dependiente de los recursos alimenticios y sobre todo de factores ambientales como la temperatura y humedad. Se sugiere que X. affinis se ha adaptado y explotado los recursos de mejor manera que X. ferrugineus en estos ecosistemas, a juzgar por la abundancia observada. Las correlaciones entre la abundancia y los factores climáticos presentaron valores positivos y negativos. Los resultados reflejan el comportamiento de las poblaciones, sin embargo, hace falta evaluar de manera detallada algunos factores biológicos y abióticos que influyen en las fluctuaciones de éstos insectos.
Palabras clave: insectos, ambrosiales, barrenadores, huésped, fluctuación.
Los Scolytinae son insectos comúnmente conocidos como coleópteros descortezadores o ambrosiales debido a su forma de alimentarse. Existen especies que restringen su actividad reproductiva a una planta huésped o a un número reducido de ellas, otras son altamente polífagas con preferencias para invadir árboles muertos, recién cortados o moribundos y algunos pueden atacar árboles vivos bajo ciertas condiciones (Wood 1982, Rudinsky 1962).
Las especies del género Xyleborus se alimentan de hongos ambrosiales que cultivan en las paredes de los túneles que barrenan. Los machos son raros y han perdido la capacidad de vuelo; solamente las hembras buscan un nuevo huésped y establecen una nueva colonia y construyen túneles que puede ser simples o complejos según el sistema de ramificaciones que produzcan (Wood 1982).
En Tabasco se han registrado especies de escolítinos que son consideradas plagas de importancia en zonas tropicales tal es el caso de X. ferrugineus (Fabricius 1801) que es un insecto que puede atacar árboles sanos, enfermos o recién muertos, infesta trocería apilada en campo o en aserraderos, eventualmente infesta pilas de madera recién aserradas y aún húmeda. Provoca la muerte de árboles aparentemente sanos al introducir hongos causantes de marchitamientos vasculares como el Ceratocystis fimbriata Ell. & Halst que causa la muerte de árboles de cacao (Cibrián et al. 1995). En madera de uso comercial, su daño consiste en las galerías que construye y en el manchado de la madera. X. affinis Eichhoff 1868, es otra especie de importancia que también infesta maderas tropicales con características similares a X. ferrugineus. Ambas especies pueden provocar grandes infestaciones en sus huéspedes (Cibrián et al. 1995). Sin embargo, el impacto que estos insectos provocan aun no ha sido evaluado y su conocimiento se ha restringido a registros esporádicos.
Por tal motivo, el conocimiento sobre el comportamiento de las poblaciones de insectos, y en especial, sus fluctuaciones a través del tiempo aunado a factores ambientales, tienen importancia desde el punto de vista ecológico, y sobre todo, si se desean implementar estrategias de manejo (Morales et al. 2000). Adicionalmente, el implementar diferentes métodos de recolecta, puede proporcionar información que coadyuve en el manejo o monitoreo de las poblaciones. Por tal motivo, el objetivo de este estudio es dar a conocer la fluctuación poblacional de X. ferrugineus y X. affinis.
Materiales y Métodos
El estudio se realizó en el Jardín Botánico José Narciso Rovirosa, durante los meses de noviembre 2010-Julio 2011; y en la Reserva Ecológica de la Chontalpa durante febrero 2010-enero 2011; se incluye también información de cuatro plantaciones de cacao durante el 2007 en Tabasco, México.
El Jardín Botánico José Narciso Rovirosa (JBU) se encuentra establecido dentro de las instalaciones de la División Académica de Ciencias Biológicas, ubicado al oeste de la ciudad de Villahermosa en el kilómetro 0.5 de la carretera 180 (costera del golfo, tramo Villahermosa-Cárdenas) en el municipio del centro, Tabasco. Se localiza entre las coordenadas 17°59’26’’17” N-92°58’16”37” W, con una superficie de 10ha con vegetación secundaria y porción de selva baja espinosa perennifolia con abundancia de árboles de Haematoxylum campechianum. La precipitación es de 2 029mm con una temperatura promedio de 27ºC (Ascencio & Maldonado 2002).
La Reserva Ecológica de la Chontalpa (REC) se localiza en el municipio de Cárdenas, en el kilómetro 21 de la carretera Cárdenas-Coatzacoalcos, Veracruz presenta una extensión de 277ha y se ubica entre los 17º59´05”51” N-93º34´13”33” W. La precipitación anual promedio es de 2 256.3mm. El clima es denominado Am (f), cálido húmedo con lluvias en verano, y la temperatura promedio anual es de 26ºC. Presenta una vegetación de bosque tropical perennifolio, que corresponde a la selva mediana perennifolia. La especie dominante es Bravaisia integerrima (Palma-López et al. 1999).
Se trabajó con cuatro plantaciones de cacao. La primera consiste de un predio de seis hectáreas y 37 años de edad, sembrada en marco real de 4x4m, la cual se localiza entre las coordenadas 17º59’23” N -93º37’09” W, a una altura de 20msnm en el Ejido Las Delicias de la Ranchería José María Morelos y Pavón, municipio de Teapa, Tabasco, México (Teapa). La fisiografía de la zona corresponde a un valle abierto, con un clima cálido húmedo con lluvias todo el año (Af), la temperatura promedio es de 26ºC. La precipitación alcanza una media anual de 3 424mm (INEGI 2006). La segunda plantación corresponde a una hectárea de aproximadamente 38 años de edad, localizada entre las coordenadas 17º37’19” N-92º27’56” W, en el kilómetro 21 de la carretera Cárdenas, Tabasco-Coatzacoalcos, Veracruz, México (km 21). La tercera plantación consta de dos hectáreas de 10 años de edad, localizadas entre las coordenadas 17º58’09” N-93º20’55” W en el Ejido El Bajío 2a sección, municipio de Cárdenas, Tabasco (El Bajío). La cuarta plantación tuvo una extensión de 3.3ha con 45 años de edad, localizada entre las coordenadas 18º02’34” N-93º22’44” W en el Ejido Río Seco 2da sección, municipio de Cárdenas, Tabasco (Río Seco). Las tres últimas plantaciones se encuentran a una altura de 10msnm, establecidas en marco real de 4x4m. Su fisiografía corresponde a la llanura costera del Golfo Sur, con clima cálido húmedo con abundantes lluvias en verano (Am), la temperatura promedio es de 26.4°C. La precipitación media anual es de 1 993.7mm (iNEGi 2005).
Captura de Xyleborus spp. con trampas utilizando como atrayente alcohol etílico: Se instalaron cinco trampas en una hectárea ubicada en el centro de cada localidad (excepto en JBU que se instalaron sólo dos trampas en las áreas de mayor vegetación), a una altura de 1.50m y 50m de distancia entre ellas formando un cuadrado con una trampa en el centro, utilizando como atrayente alcohol etílico desnaturalizado con octa-acetato de sacarosa al 70%. Las trampas permanecieron en campo durante todo el periodo de estudio (Bustamante & Atkinson 1984, iturre & Darchuck 1996, Pérez-De la Cruz et al. 2009). La recolección de los insectos atraídos en cada una de las trampas se realizó quincenalmente, y los especímenes se conservaron en alcohol al 70% para su posterior determinación. En cada recolección se procedió a reponer el atrayente.
Captura de Xyleborus spp. con trampa de luz fluorescente: Se utilizó una trampa similar a la de alcohol para recolectar a los insectos sustituyendo el material atrayente por una fuente de luz fluorescente proporcionada por una lámpara recargable marca Lloyd´s de 20watts con una duración aproximada de cuatro horas, colocada ésta a un costado de la trampa. La trampa se instaló en el centro de la plantación en cada una de las localidades de estudio (este método sólo fue usado en el agro-ecosistema cacao) a una altura de 1.50m. La trampa fue colocada a las 18:00 horas y fue levantada al día siguiente; los muestreos se realizaron mensualmente. Los especímenes se conservaron en alcohol al 70% para su posterior determinación (Pérez-De la Cruz
et al. 2009).
Captura directa de Xyleborus spp. en sus plantas huésped: La recolecta consistió en hacer revisiones directamente sobre las partes de las plantas (tallo, ramas y frutos) en donde se observó daños de estos insectos. La presencia de aserrín, huecos de entrada y grumos de madera fueron de gran utilidad en el campo para detectar el ataque de estos barrenadores (Bustamante & Atkinson 1984). Los muestreos se realizaron quincenalmente durante el año de estudio y sólo en el agro-ecosistema cacao. El material vegetal que no pudo ser revisado en campo debido al grosor de las ramas fue llevado al laboratorio y colocado en cámaras de emergencia para la recolección de los insectos. Los organismos recolectados se conservaron en alcohol al 70% para su posterior determinación (Pérez-De la Cruz et al. 2009).
Además se realizó una correlación simple entre la precipitación y temperatura con los datos de abundancia mensual de las especies para conocer el grado de asociación entre las variables (infante & Zárate 1990).
Identificación del material biológico: La determinación taxonómica de los insectos se realizó mediante el uso de claves taxonómicas (Wood 1982, 1986) y comparaciones con material depositado en la colección del Colegio de Postgraduados Campus Montecillo, México (CEAM). Para obtener los datos de plantas huésped y contribuciones sobre biología se consultaron las siguientes publicaciones: Schedl (1940), Atkinson & Equihua (1985a, b, 1988), Equihua & Atkinson (1986), Estrada & Atkinson 1988, Wood & Bright (1992a, b), Cibrián et al. (1995), iturre & Darchuck (1996), Romero et al. (1997), Morales et al. (2000), Bright & Skidmore (1997, 2002), Equihua & Burgos (2002), Pérez-De la Cruz et al. (2009). Todos los especímenes quedaron depositados en la colección de insectos de la División Académica de Ciencias Biológicas de la Universidad Juárez Autónoma de Tabasco.
Resultados
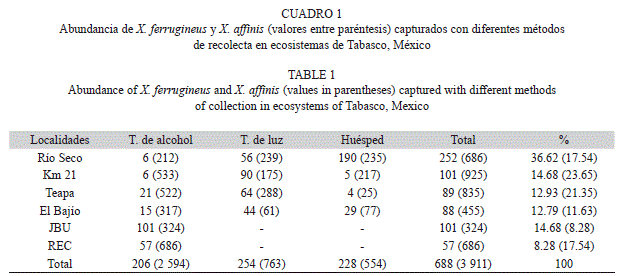
Se recolectaron 688 especímenes de X. ferrugineus con los tres métodos de recolecta, siendo las trampas de luz las que registraron el mayor número de organismos con 254 (37%), seguido de las plantas huésped con 228 (33%) y por último en las trampas de alcohol con 206 (30%). En la plantación de cacao de Río Seco se registró el mayor número de insectos con 36.62% y el menor lo presentó la REC con 8.28%. A diferencia de X. affinis que registró una abundancia de 3 911 especímenes con los tres métodos de recolecta, siendo las trampas de alcohol las que registraron el mayor número de organismos con 2 594 (66%), seguido de las trampas de luz con 763 (20%) y por último en las plantas huésped con 554 (14%). En el kilómetro 21 se registró el mayor número de insectos con el 24% y el menor lo presentó JBU con el 8% (Cuadro 1).
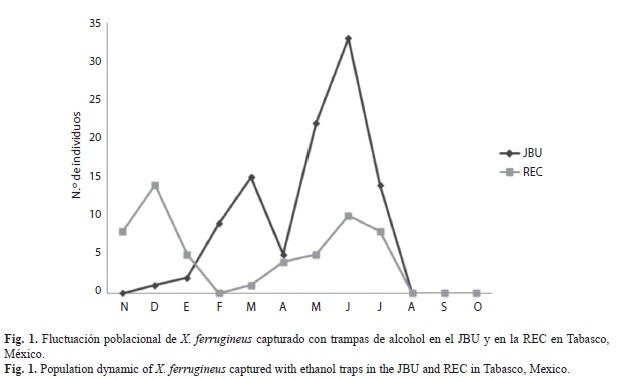
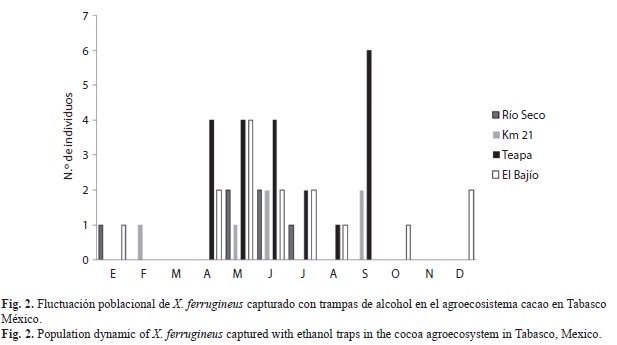
La fluctuación poblacional de X. ferrugineus capturado con trampas de alcohol en el JBU presentó su máximo pico poblacional durante el mes de junio (con 33-individuos), coincidiendo con temperaturas promedio de 28°C y alto porcentaje de humedad superior a 73%, estando presente durante la mayor parte del periodo de estudio (excepto en noviembre que no se recolectaron organismos) pero con poblaciones bajas. A diferencia de las poblaciones encontradas en la REC que presentó su máximo pico poblacional en diciembre (14-individuos) y otro en junio con un número de organismos relativamente bajo (Fig. 1). En el agro-ecosistema cacao las poblaciones de X. ferrugineus fueron bajas (menor a 10-individuos/mes) presentando picos poblacionales marcados en mayo con una abundancia no mayor a 11 individuos (este mes marca el fin de la época seca e inicio de la época lluviosa) en la mayoría de los sitios a diferencia de la localidad de Teapa que la presentó en septiembre con seis individuos (Fig. 2).
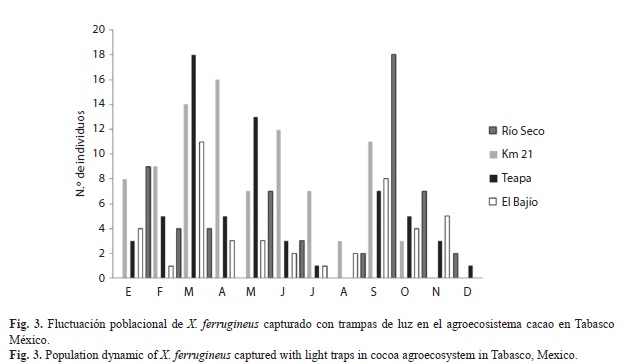
La fluctuación poblacional de X. ferrugineus capturado en trampas de luz en el agro-ecosistema cacao presentó varios picos poblacionales marcados a través del periodo de estudio siendo los más destacados en Teapa y Río Seco en los meses de marzo y octubre, respectivamente; en el kilómetro 21 en abril y en el Bajío en marzo, aunque los resultados sugieren que estos insectos se encuentran la mayor parte del año y pueden ser recolectados con mayor eficiencia durante los meses de marzo, abril y octubre con este método de captura. La localidad con el mayor número de insectos capturados fue el kilómetro 21 con 35% y el mínimo en El Bajío con 17% (Fig. 3).
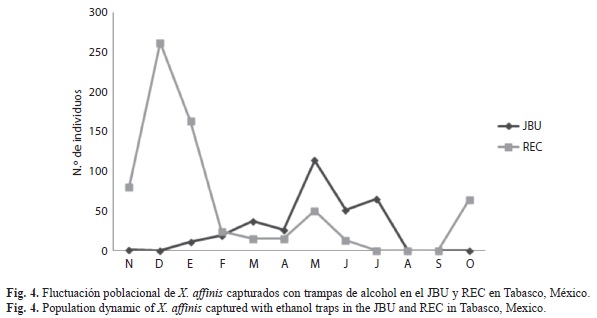
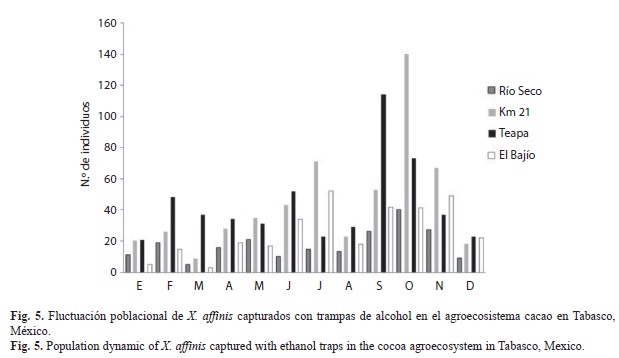
La fluctuación poblacional de X. affinis capturado en trampas de alcohol en el JBU presentó su máximo pico poblacional durante el mes de mayo, estando presente durante la mayor parte del periodo de estudio (excepto en diciembre) pero con poblaciones bajas. A diferencia de las poblaciones encontradas en REC que presentó su máximo pico poblacional en diciembre (Fig. 4). En el agro-ecosistema cacao las poblaciones de X. affinis registraron picos poblacionales marcados en octubre en el kilómetro 21 y en septiembre en Teapa. El Bajío y Río Seco presentaron poblaciones bajas y estables a lo largo del periodo de estudio (Fig. 5).
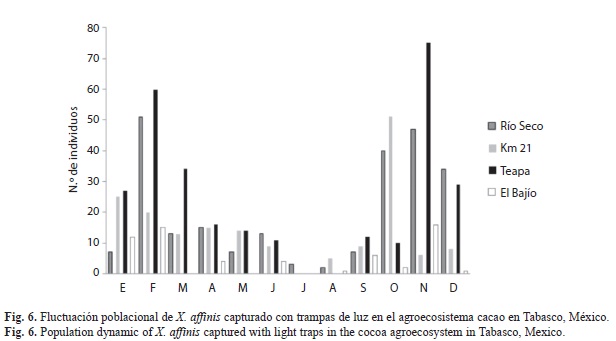
La fluctuación poblacional de X. affinis capturado en trampas de luz en el agro-ecosistema cacao presentó dos picos poblacionales marcados, uno en febrero y otro en octubre-noviembre, la localidad con el mayor número de insectos capturados fue Teapa con el 38% y el mínimo fue en El Bajío con 8% (Fig. 6).
Descripción de X. ferrugineus: Las hembras de X. ferrugineus son de cuerpo cilíndrico, de 2 a 3.3mm de longitud, color café rojizo. Declive elitral aplanado, con pendiente moderada; en la base de la interestría i hay uno o dos gránulos pequeños; en la parte media del declive en la interestría iii se encuentra un dentículo prominente y puntiagudo que sirve para reconocer con facilidad a esta especie (Fig. 7). El macho es más pequeño que la hembra, de 1.8mm de longitud. Con respecto al ciclo de vida y hábitos, se presentan múltiples generaciones por año, con estados sobrepuestos. Sólo las hembras son capaces de volar, los machos no debido a que éstos no pueden desplegar sus alas. Las hembras viven de manera comunal, haciendo galerías multi-ramificadas. Introducen hongos ambrosiales, los cuales colonizan la madera y se reproducen en ella, las estructuras de reproducción se forman en la luz de las galerías y constituyen el alimento para adultos y larvas; dichas larvas viven libremente dentro del sistema de galerías, sin estar en nichos especiales. Cuando maduran pupan en el interior de las galerías y los nuevos adultos salen por los orificios de entrada de las madres. Son insectos sub-sociales en donde hay reproducción por partenogénesis. Existe una baja producción de machos (Cibrián et al. 1995).
Huéspedes: X. ferrugineus es un insecto considerado polífago debido a la gran diversidad de plantas que utiliza como huéspedes, entre las cuales se encuentran: Colubrina arborescens, Erythrina americana, Arctocarpus altilis, Glyricidia sepium, Diphysa robinoides, Theobroma cacao, Mangifera indica, Spondias mombin, Tabebuia rosea, Swietenia macrophylla, Acacia gaumeri, Aspidosperma megalocarpon, Bursera simaruba, Bombacopsis quinatum, Brosimun alicastrum, Cecropia obtusifolia, Cededrela odorata, Citrus spp., Dendropanax arboreum, Inga spp., Lonchocarpus margaritensis, Lysiloma bahamensis, Manilkara zapota, Quercus spp., Heliocarpus pallidus, Ficus sp., Urera sp. Sciadodendro excelsum, Thouinidium decandrum, Acrocarpus fraxinifolius, Vitex guameri, Eucalyptus spp. Dialium guianense, Terminalia amazonica, Rheedia sp., Heliocarpus appendiculatus.
Descripción de X. affinis: El declive elitral de las hembras de X. affinis es gradualmente convexo, con superficie opaca, gránulos pequeños de igual tamaño en las interestrías decliviales I y III; vestidura de setas largas evidentes, coloración amarillenta a café rojizo; longitud de 2.0-2.7mm (Fig. 8).
Huéspedes de X. affinis: Dhiphysa robinoides, Theobroma cacao, Mangifera indica, Colubrina arborescens, Tabebuia rosea, Ficus tecolutensis, Swietenia macrophylla, Alexa imperatricia, Bombacopsis quinata, Brosimum sp., Brosimum alicastrum, Bursera simaruba, B. instabilis, Cecropia peltata, Cespedesia macrophila, Cleythra hondurensis, Couma macrocarpa, Dendropanax arboreum, Eschweilera corrugata, E. grata, Erytrina costaricensis, Icica allisima, Lecythia sp., Licania sp., Miconia globuliflora, Pouteria anibaefolia, P. egregia, Sacoglottis procera, Schizolobium sp., Sloanea multiflora, Spondias purpurea, S. mombin, Taxodium sp., Terminalia amazonia, Toulicia pulvinata, Trichilia propingon, Metopium brownei, Acrocarpus flaxinifolius,Cedrela odorata, Croton nitens, Acacia gaumeri, Manilkara zapota, Dialium guianense, Rheedia sp.
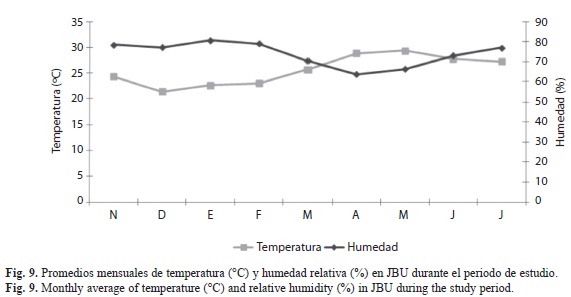
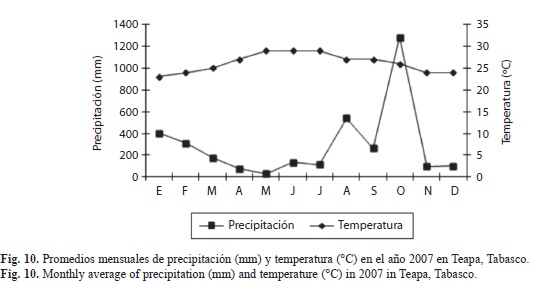
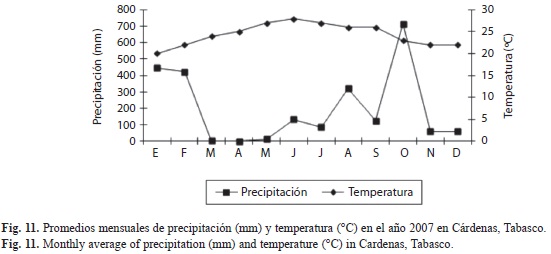
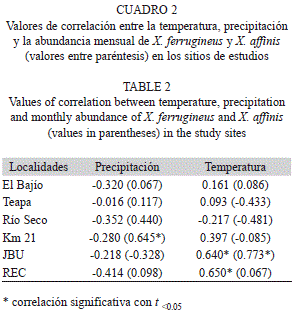
Correlación entre los factores climáticos y la abundancia de X. ferrugineus y X. affinis: En las figuras 9, 10 y 11 se muestran los factores climáticos (temperatura, precipitación y humedad) de los sitios de estudio, tomados de las estaciones meteorológicas más cercanas. En ellas se observan dos periodos bien definidos en la región, el de sequía con lluvias escasas (marzo-mayo) y el periodo de precipitación (es variable de junio-febrero), alcanzando la máxima precipitación en el mes de octubre. Las temperaturas promedio fluctúan entre los 24 y los 29°C. El análisis de correlación mostró diferentes grados de ocurrencia en X. ferrugineus y X. affinis en función de la precipitación y temperaturas medias, registrándose valores positivos y negativos en sus correlaciones, con algunas diferencias significativas (Cuadro 2). Estos resultados nos indican que las poblaciones de estos insectos están influenciadas de manera tal que pueden promover el incremento o en su momento el decremento de las poblaciones mediante el aumento o disminución de algún factor, como la temperatura que presentó mayor grado de asociación con la abundancia de estos insectos. Ambas especies pueden verse afectadas de manera positiva o negativa por factores abióticos tales como; la temperatura y precipitación los cuales son determinantes en la distribución y abundancia de los escolítinos en los ecosistemas donde habitan aunado a los factores bióticos.
Discusión
De los métodos de recolecta utilizados, la trampa de luz registró un mejor efecto para atraer a X. ferrugineus de acuerdo a lo observado en los sitios del agro-ecosistema cacao donde fueron implementados ambos métodos (Trampa de luz: 254 especímenes y trampa de alcohol: 48), a diferencia de lo observado para X. affinis donde la trampa de alcohol registró mayor captura en los mismos sitios (Trampa de alcohol: 1 584 especímenes y trampas de luz: 763). Cabe mencionar que las trampas de alcohol permanecieron activas durante todo el periodo de estudio y las de luz sólo se colocaron una vez por mes, por lo que esto podría ayudar a explicar estas diferencias. Ambos métodos de recolecta pueden ser complementarios para el monitoreo de las poblaciones de estos insectos y tener una idea más precisa para sugerir el momento adecuado de implementar un manejo.
La fluctuación poblacional de X. ferrugineus mostró de manera general poblaciones bajas en los ecosistemas estudiados con picos poblacionales sin alguna estacionalidad marcada, registrándose picos poblaciones tanto en época seca (marzo-abril-mayo) como en lluviosa (junio, septiembre-octubre, diciembre) aunque con tendencia a un mayor número de organismos en periodos de precipitación y alta humedad. A diferencia de lo reportado por Morales et al. (2000) en zonas reforestadas con Eucalyptus grandis en Minas Gerais, Brasil, donde ésta especie presentó su máxima abundancia en época lluviosa y disminuyó fuertemente durante la época seca. Las poblaciones de X. affinis fueron más notables en los ecosistemas estudiados mostrando en la mayoría de los sitios (excepto en JBU que se presentó en mayo) picos poblacionales en épocas lluviosas (septiembre-diciembre) a diferencia de lo reportado por Morales et al. (2000), que reportó picos poblacionales máximos para la época seca y con disminución en la época lluviosa. En general ambas especies estudiadas se encuentran presente durante la mayor parte del año y su fluctuación poblacional es dependiente de los recursos alimenticios y sobre todo de factores ambientales como la temperatura y humedad los cuales influyen en la abundancia de ambas especies, aunado a las características de cada sitio, como la diversificación vegetal, edad, competidores, depredadores, manejo de los sitios, los cuales son factores determinantes en la distribución y abundancia de los escolítinos (Rudinsky 1962, Wood 1982, Pérez-De la Cruz et al. 2009). Por lo que se sugiere que X.affinis se ha adaptado y explotado los recursos de mejor manera que X. ferrugineus en estos ecosistemas a juzgar por la abundancia observada. Ambas especies tienen una amplia distribución con una lista considerable de huéspedes las cuales se podrían ir incrementando de acuerdo al interés por conocer la amplia gama de plantas que pueden llegar a utilizar como recurso alimenticio.
Los resultados obtenidos muestran el estado de las poblaciones de éstos insectos barrenadores, sin embargo, hacen falta estudios detallados sobre ciclos de vida, horas de vuelo, depredadores, así como otros factores ambientales locales que ayuden a entender el comportamiento de las poblaciones, para establecer un manejo apropiado. Sin embargo, se puede sugerir como método de control para ambas especies el derribo y destrucción de los árboles que estén atacados por estos barrenadores, además la incineración o bien la fragmentación de la madera de los troncos dañados; con ello se logra un secado rápido y la muerte de los insectos y los hongos asociados o bien establecer los métodos de recolecta aquí empleados como una herramienta preventiva, con el propósito de disminuir las poblaciones de estos barrenadores en la región, durante los meses en donde las poblaciones tienden a incrementarse. En aprovechamientos forestales se recomienda no dejar la trocería almacenada en el campo por periodos prolongados; la misma recomendación es extensiva a los patios de almacenamiento en aserraderos. Cuando se requiera mantener trocería durante largo tiempo, ésta se puede proteger con un insecticida de contacto de persistencia moderada mezclado con un adherente; la aplicación se debe hacer antes de que los insectos ataquen (Cibrián et al. 1995).
Agradecimientos
Los dos primeros autores agradecen a las personas que han influido en la conservación de especies de árboles tropicales en el Jardín Botánico José Narciso Rovirosa dentro de las instalaciones de la División Académica de Ciencias Biológicas con el proyecto “El Jardín Botánico José Narciso Rovirosa un área prioritaria de rescate y conservación ex situ de especies en la NOM-059, clave TAB-2009-C18-122207, financiado por el Fondo Mixto-CONACYTGobierno del Estado de Tabasco. Así como, se agradece a Ena Mata Zayas y a los revisores por las contribuciones hechas al manuscrito.
Referencias
Ascencio, J. & F. Maldonado. 2002. El jardín Botánico Universitario “José Narciso Rovirosa” un esfuerzo para la conservación de la flora tabasqueña. Kuxulkab Rev. Divul. 7: 34-47. [ Links ]
Atkinson, T. & A. Equihua. 1985a. Notes on biology and distribution of Mexican and Central American Scolytidae (Coleoptera). I. Hylesininae, Scolytinae except Cryphalini and Corthylini. Coleopts. Bull. 39: 227-238. [ Links ]
Atkinson, T. & A. Equihua. 1985b. Notes on biology and distribution of Mexican and Central American Scolytidae (Coleoptera). II. Scolytinae: Cryphalini and Corthylini. Coleopts. Bull. 39: 355-363. [ Links ]
Atkinson, T. & A. Equihua. 1988. Notas sobre la biología de Scolytidae y Platypodidae (Coleoptera) de México y Centroamérica. Folia Entomol. Mex. 76: 83-105. [ Links ]
Bright, D. & R. Skidmore. 1997. A Catalog of Scolytidae and Platypodidae (Coleoptera), Suplement 1 (1990-1994). NRC, Ottawa, Canada. [ Links ]
Bright, D. & R. Skidmore. 2002. A Catalog of Scolytidae and Platypodidae (Coleoptera), Suplement 2 (1995-1999). NRC, Ottawa, Canada. [ Links ]
Bustamante, F. & T. Atkinson. 1984. Biología del barrenador de las ramas del peral Corthylus fuscus Blandford (Coleoptera: Scolytidae), en el norte del Estado de Morelos. Folia Entomol. Mex. 60: 83-101. [ Links ]
Cibrián, D., J. Méndez, R. Campos, O. Yates III & J. Flores. 1995. Insectos Forestales de México/Forest insects of Mexico. Universidad Autónoma Chapingo, Estado de México, México. [ Links ]
Equihua, A. & T. Atkinson. 1986. Annotated checklist of bark and ambrosia beetles (Coleoptera: Scolytidae and Platipodidae) associated with a tropical deciduous forest at Chamela, Jalisco, Mexico. Florida Entomol. 69: 619-635. [ Links ]
Equihua, A. & A. Burgos. 2002. Scolytidae, p 539-557. In J. Llorente & J.J. Morrone (eds.). Biodiversidad, Taxonomía y Biogeografía de artrópodos de México: Hacia una síntesis de su conocimiento. Vol. III. CONABIO-IBUNAM, México. [ Links ]
Estrada, A. & T. Atkinson. 1988. Scolytidae y Platypodidae (Coleoptera) de Escárcega, Campeche, México. Biogeografía, biología, importancia económica y una lista comentada de especies. An. Inst. Biol. Univ. Nac. Autón. Méx. Ser. Zool. 58: 199-220. [ Links ]
INEGI (instituto Nacional de Estadística Geografía e informática). 2005. Cuaderno estadístico Municipal, Cárdenas, Tabasco, México. [ Links ]
INEGI (instituto Nacional de Estadística Geografía e informática). 2006. Cuaderno estadístico Municipal, Teapa, Tabasco, México. [ Links ]
Infante, S. & P. Zárate. 1990. Métodos estadísticos un enfoque interdiciplinario. Trillas, México. [ Links ]
Iturre, M. & E. Darchuk. 1996. Registro de escolítidos relacionados al género Eucalyptus en Santiago del Estero. Rev. Quebracho 4: 11-16. [ Links ]
Morales, N., C. Zanuncio, D. Pratissoli & S. Fabres. 2000. Fluctuación poblacional de Scolytidae (Coleoptera) en zonas reforestadas con Eucalyptus grandis (Myrtaceae) en Minas Gerais, Brasil. Rev. Biol. Trop. 48: 101-107. [ Links ]
Palma-López, D., E. Garcia, A. Sol & F. Juárez. 1999. Plan de manejo de la Reserva Ecológica de la Chontalpa. Colegio de Postgraduados, Campus Tabasco y SEDESPA, Dirección de Ecología. H. Cárdenas, Tabasco, México. [ Links ]
Pérez-De la Cruz, M., A. Equihua, J. Romero, S. Sánchez & E. García. 2009. Diversidad, fluctuación poblacional y plantas huésped de escolítinos (Coleoptera: Curculionidae) asociados con el agroecosistema cacao en Tabasco, México. Rev. Mex. Biodiversidad 80: 779-791. [ Links ]
Romero, J., S. Anaya, A. Equihua & H. Mejía. 1997. Lista de Scolytidae de México (insecta: Coleoptera). Acta Zool. Mex. 70: 35-53. [ Links ]
Rudinsky, L. 1962. Ecology of Scolytidae. Annu. Rev. Entomol. 7: 327-348. [ Links ]
Schedl, K. 1940. Scolytidae, Coptonotidae y Platypodidae mexicanos. An. Esc. Nac. Cienc. Biol. IPN. 1: 317-378. [ Links ]
Wood, S. 1982. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), a Taxonomic Monograph. Great Basin Naturalist Memoirs 6: 1-1327. [ Links ]
Wood, S. 1986. A reclassification of the genera of Scolytidae (Coleoptera). Great Basin Naturalist Memoirs 10: 1-126. [ Links ]
Wood, S. & D. Bright. 1992a. A catalog of Scolytidae and Platypodidae (Coleoptera), Part 2: Taxonomic index. Great Basin Naturalist Memoirs 13: 1-833. [ Links ]
Wood, S. & D. Bright. 1992b. A catalog of Scolytidae and Platypodidae (Coleoptera), Part 2: Taxonomic index. Great Basin Naturalist Memoirs 13: 835-1553. [ Links ]
*Correspondencia: Ricardo Rangel: Universidad Juárez Autónoma de Tabasco, División Académica de Ciencias Biológicas, Código Postal 86150, Carretera Villahermosa-Cárdenas km 0.5 s/n entronque a Bosque de Saloya, Villahermosa, Tabasco, México; richardrm_monitoreo@hotmail.com
Manuel Pérez: Universidad Juárez Autónoma de Tabasco, División Académica de Ciencias Biológicas, Código Postal 86150, Carretera Villahermosa-Cárdenas km 0.5 s/n entronque a Bosque de Saloya, Villahermosa, Tabasco, México; perezmandoc@hotmail.com
Saúl Sánchez: Colegio de Postgraduados, Campus Tabasco, Apartado postal 24 86500, H. Cárdenas Tabasco, México; sssoto@colpos.mx
Silvia Capello: Universidad Juárez Autónoma de Tabasco, División Académica de Ciencias Biológicas, Código Postal 86150, Carretera Villahermosa-Cárdenas km 0.5 s/n entronque a Bosque de Saloya, Villahermosa, Tabasco, México;capellogs@hotmail.com
1. Universidad Juárez Autónoma de Tabasco, División Académica de Ciencias Biológicas, Código Postal 86150, Carretera Villahermosa-Cárdenas km 0.5 s/n entronque a Bosque de Saloya, Villahermosa, Tabasco, México; richardrm_monitoreo@hotmail.com, perezmandoc@hotmail.com, capellogs@hotmail.com
2. Colegio de Postgraduados, Campus Tabasco, Apartado postal 24 86500, H. Cárdenas Tabasco, México; sssoto@colpos.mx
Recibido 13-XII-2011. Corregido 20-V-2012.Aceptado 20-VI-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}