










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Variación temporal y espacial de aves playeras en la laguna Barra de Navidad, Jalisco, en tres temporadas no reproductivas
*Dirección para correspondencia
Abstract
Temporal and spatial variation of shorebirds in Barra de Navidad lagoon, Jalisco, during three non-breeding seasons. Resident and migratory shorebirds inhabit different kinds of wetlands such as lagoons, rivers and seashores among others. In recent years, these areas have been importantly affected by urban, agriculture and touristic activities, such as the Barra de Navidad lagoon, for which little information is available to support conservation programs. The aim of this work was to describe shorebirds temporal and spatial distribution in Barra de Navidad lagoon during three non-breeding seasons (1999-2000, 2006-2007 and 2008-2009). For this, monthly censuses were performed from November-April with the purpose of registering all the shorebirds species. We were able to identify 19 shorebirds species (three residents and 16 winter visitors), of which Charadrius wilsonia, Limosa fedoa and Tringa semipalmata were the most abundant. The greater number of species was registered for November, December and March of the first and third seasons. The greater number of individuals was registered when birds were feeding during low tides, mainly in December, January and February of the first and third seasons. At low tide, there was a great number of species and individuals in zone C. This area had muddy substrates that were exposed during low tides and were used to feed. Barra de Navidad lagoon provided suitable habitats for feeding and resting for resident and migratory birds. Twelve of the 19 species were considered as priority within the Mexican bird conservation strategy. However, these habitats are threatened by human activities performed in the nearby areas of the lagoon that may have negative consequences for the distribution, abundance and conservation of these species.
Key words: shorebirds, conservation, wetlands, migratory birds, México.
Resumen
Hay un escaso conocimiento de las aves playeras en los humedales costeros de Jalisco, y en particular en la laguna Barra de Navidad. El presente trabajo contribuye al conocimiento de este grupo de aves y describe su distribución temporal y espacial en la laguna Barra de Navidad durante tres temporadas no reproductivas (1999-2000, 2006-2007 y 2008-2009). Se realizaron censos mensuales de noviembre-abril en las tres temporadas con el fin de registrar todas las especies de aves playeras. Se identificaron 19 especies (tres residentes y 16 visitantes de invierno), de las cuales Charadrius wilsonia, Limosa fedoa y Tringa semipalmata presentaron la mayor abundancia. Doce especies son consideradas como prioritarias en la “Estrategia para la Conservación y Manejo de las Aves Playeras y su Hábitat en México”. El mayor número de especies fue registrado en noviembre, diciembre y marzo en la primera y tercera temporada. El mayor número de individuos fue registrado alimentándose en marea baja, principalmente en diciembre, enero y febrero de la primera y tercera temporada. En marea baja hubo un mayor número de especies e individuos alimentándose en la zona C. Esta zona se caracterizó por tener sustratos lodosos expuestos durante marea baja y que fueron aprovechados por las aves para alimentarse. La laguna Barra de Navidad proporcionó hábitats de alimentación y descanso para las aves residentes y migratorias. Sin embargo, estos hábitats se ven amenazados por las actividades humanas realizadas dentro de la laguna, que sin duda tendrán consecuencias negativas para la distribución y abundancia de las aves playeras.
Palabras clave: aves playeras, conservación, humedal costero, aves migratorias, México.
A nivel mundial, las aves playeras están representadas por 217 especies, de las cuales 53 habitan en México, siendo la región de la Península de Baja California el lugar que posee la mayor riqueza y abundancia de estas aves en el país (O’Brien et al. 2006, SEMARNAT 2008). Para la costa de Jalisco (19°11’ - 20°44’ N y 104°40’ - 105°24’ W) se han registrado 34 especies de aves playeras (Cupul-Magaña 2000a,b, Hernández-Vázquez et al. 2010), que representan el 35.8% de las especies reportadas para México.
Las aves playeras se encuentran en una gran variedad de hábitats, tales como: lagunas, ríos, lagos, pantanos, playas, llanuras salinas y áreas de cultivo (O’Brien et al. 2006). En los hábitats costeros este grupo de aves está relacionado con las variaciones en las mareas, debido a que tienden a concentrarse en áreas expuestas durante mareas bajas en donde la densidad y disponibilidad de las presas es alta y la energía requerida para capturarlas es relativamente baja (Goss-Custard et al. 1977). Las variaciones en el nivel del agua, causado por las diferencias en las mareas, es un factor importante que influye a escala local en la distribución y abundancia de las aves playeras (Butler et al. 2001, Warnock et al. 2002).
Muchas especies son migratorias y realizan grandes desplazamientos desde el norte del continente, donde se reproducen, hasta el sur del continente donde pasan el invierno. Durante este viaje las aves playeras usan los recursos disponibles en los diferentes humedales, y las condiciones ambientales encontradas en ellos son de gran relevancia para la sobrevivencia de este grupo de aves (Myers et al. 1987).
La costa de Jalisco tiene varios humedales relativamente pequeños que sirven como sitios de parada para las aves migratorias, donde se alimentan y descansan para continuar su viaje (Hernández-Vázquez et al. 2010). En la zona sur de dicha costa se ubica la laguna Barra de Navidad, la cual, a diferencia de los humedales cercanos, ha sufrido grandes cambios en los últimos años, debido al crecimiento urbano y turístico que se ha desarrollado en sus márgenes.
Hay poca información publicada que haga referencia de las aves playeras en esta laguna. En un estudio realizado por Hernández- Vázquez et al. (2010), se da información de las aves acuáticas en varios humedales de la costa de Jalisco y se aborda de forma general algunos aspectos de la distribución y abundancia de aves playeras en la laguna Barra de Navidad. Se han realizado otros estudios en humedales cercanos a esta laguna, tal es el caso del estero La Manzanilla (Hernández-Vázquez 2000), Majahuas y El Chorro (Hernández-Vázquez & Mellink 2001) y Agua Dulce-Ermitaño (Hernández-Vázquez 2005). Aunque estos estudios se enfocaron a monitorear los diferentes grupos de aves acuáticas, también abordan aspectos generales de abundancia y distribución de las aves playeras. Sin embargo, en ninguno de estos estudios se presenta información de las abundancias y distribución en una escala temporal más amplia, ni se presenta información de la distribución espacial de las aves playeras. Este estudio tiene como objetivo identificar las especies de aves playeras en la laguna Barra de Navidad, así como evaluar su distribución temporal y espacial en tres temporadas no reproductivas (1999-2000, 2006-2007 y 2008-2009). Con esta información se pretende contribuir al conocimiento de las aves playeras en el pacífico central mexicano y particularmente en la laguna Barra de Navidad, además de ser una importante herramienta de conservación que permitirá conocer los efectos del desarrollo desmedido que se está dando en la zona, sus consecuencias a futuro y su efecto sobre las aves playeras.
Materiales y métodos
Área de estudio: La laguna Barra de Navidad se localiza en la costa del pacífico central mexicano, en el estado de Jalisco (19°10’ - 19°12’ N y 104°39’ - 104°41’ W). Esta laguna se encuentra dentro de las Regiones Marinas Prioritarias (RMP-27, Punta Graham-El Carrizal) (Arriaga-Cabrera et al. 1998) y Región Hidrológica Prioritaria (RHP-25, Río Purificación-Armería) (Arriaga-Cabrera et al. 2002) para México. En 2008 fue decretada como sitio RAMSAR por la Convención Internacional de Humedales (Ramsar 2011). En 2009 fue considerada dentro de los Manglares Prioritarios para la Conservación por la CONABIO (Silva- Bátiz et al. 2009).
La laguna tiene una superficie de 382 hectáreas de espejo de agua y 620 hectáreas de manglar. Está conectada permanentemente con el mar por un canal de 80m de ancho con profundidades de hasta 7-8m, cuya parte central es periódicamente dragada para permitir la navegación. La masa de agua de la laguna se encuentra fuertemente relacionada con mareas baroclínicas y barotrópicas y la circulación de la misma depende de ellas. Los principales aportes de agua dulce son por el Arroyo Seco y un canal artificial que la conecta con el Río Marabasco (Silva-Bátiz et al. 2009). La laguna Barra de Navidad, a diferencia de las demás, se caracteriza por estar rodeada de asentamientos humanos; al noroeste se encuentra el poblado Barra de Navidad, al oeste Colimilla y el desarrollo turístico Isla Navidad que cuenta con campos de golf al margen de la laguna. En su interior se encuentra la isla Los Puercos y El Tepelolote (Fig. 1).
Métodos de muestreo: Se realizaron visitas mensuales a la laguna de Barra de Navidad durante tres temporadas no reproductivas: noviembre 1999-abril 2000, noviembre2006- abril 2007 y noviembre 2008-abril 2009. Los conteos se realizaron por las mañanas entre las 07:00-10:00 am, independientemente de la condición de la marea. Con el fin de monitorear todos los hábitats de la laguna y registrar el mayor número de especies, cada censo se realizó a lo largo de un transecto dentro del espejo de agua y siguiendo una ruta paralela al margen de la laguna. La distancia entre el transecto y la orilla de la laguna fue de 50m (en un estudio previo se observó que a esta distancia no se causaba disturbio en la actividad de las aves). Los recorridos fueron realizados con una embarcación de tres metros de eslora con un motor fuera de borda de ocho hp a una velocidad 10km/h. A lo largo del transecto se contaron todas las especies e individuos observados a 50m a la izquierda y 50m a la derecha de la embarcación, cubriendo una franja de 100m de ancho y 10 400m de largo, con un área total de barrido de 104 hectáreas en toda la laguna. Con el fin de conocer la distribución espacial de las aves, toda el área de barrido fue dividida en cuatro zonas (A, B, C y D) con un área de 26 hectáreas cada una.
Las características de las cuatro zonas son diferentes. Las zonas A y B presentaron mucha actividad humana. En sus márgenes hay asentamientos humanos y desarrollos turísticos, originando una alta actividad turística, pesquera y un frecuente tráfico de embarcaciones pequeñas y veleros. En estas zonas se encuentran tres bajos o áreas expuestos durante mareas bajas: La Playa, El Remanso y Colimilla. La zona C es la más alejada de los asentamientos humanos y se encuentra rodeada por manglar. En esta zona se da el aporte de agua dulce provenientes del río Marabasco y Arroyo Seco lo que origina una alta concentración de sedimento y áreas someras. En marea baja queda expuesto el bajo La Soledad. En la zona D se realizan actividades principalmente de pesca artesanal, y su angosto margen es cubierto por el mangle, lo que limita las áreas de sustratos blandos usados por las aves, principalmente durante marea alta. En marea baja queda expuesto El Conchero, que es un bajo con sedimentos arenosos (Fig. 1).
Las aves fueron identificadas con ayuda de binoculares 10x50 y un telescopio 15-60x, con base en las guías de campo de National Geographic Society (1987, 2006). El nombre científico y el orden del listado de las especies se realizaron con base en la lista de aves de Norteamérica de la American Ornithologist’s Union (A.O.U. 2011).
Las mediciones del nivel de marea se obtuvieron de las cartas de marea del Centro de Investigación Científica y de Educación Superior de Ensenada (CICESE) para la región de Manzanillo, Colima, México.
Los datos fueron analizados con el software Statistical 7 para Windows. Debido a que los datos no presentaron una distribución normal (Kolmogorov-Smirnov, p>0.05), se usaron pruebas de comparación de medias para determinar las diferencias del número de especies e individuos entre temporadas (1999-2000, 2006-2007 y 2008-2009) y entre zonas (A, B, C y D). Para comparaciones de más de dos medias se utilizó la prueba de Kruskal-Wallis y para comparar dos medias se usó la prueba de Mann-Whitney. La correlación de rango de Spearman se usó para ver la posible relación entre el número de especies e individuos con el nivel del agua. En todas las pruebas se usó un nivel de significancia de 0.05 (Zar 1974).
Resultados
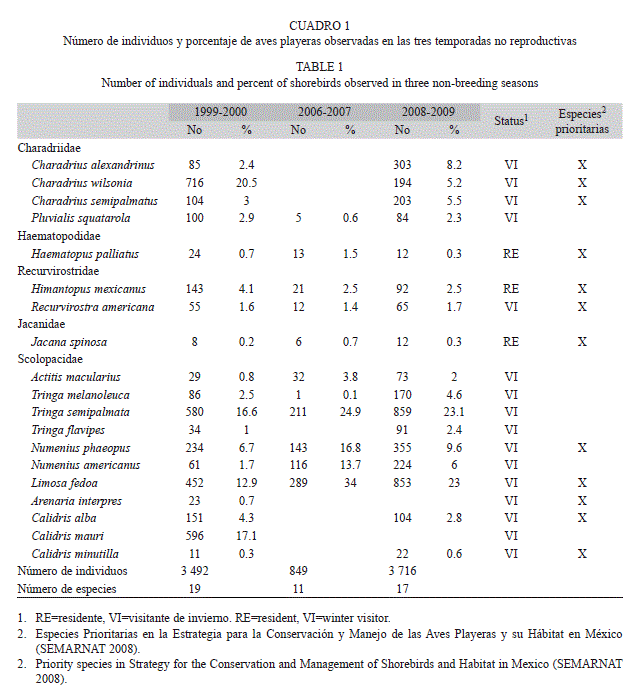
Composición de especies: Se identificaron 19 especies de aves playeras en las tres temporadas de estudio: tres residentes y 16 visitantes de invierno. En 1999-2000 se registró una abundancia acumulada de 3 492 individuos y 19 especies, de las cuales Charadrius wilsonia (716 individuos, 20.5%), Calidris mauri (596 individuos, 17.1%) y Tringa semipalmata (580 individuos, 16.6%) fueron las especies con la mayor abundancia. En 2006-2007 se contabilizaron 849 individuos y 11 especies; las especies más representativas fueron Limosa fedoa (289 individuos, 34%) y T. semipalmata (211 individuos, 24.9%). Para la temporada 2008-2009 se registró el mayor número de individuos, con una abundancia acumulada de 3 716 individuos y 17 especies. Las especies dominantes fueron: T. semipalmata (859 individuos, 23.1%) y L. fedoa (853 individuos, 23%). De las 19 especies registradas 12 son consideradas como prioritarias en la Estrategia para la Conservación y Manejo de las Aves Playeras y su Hábitat en México (SEMARNAT 2008) (Cuadro 1).
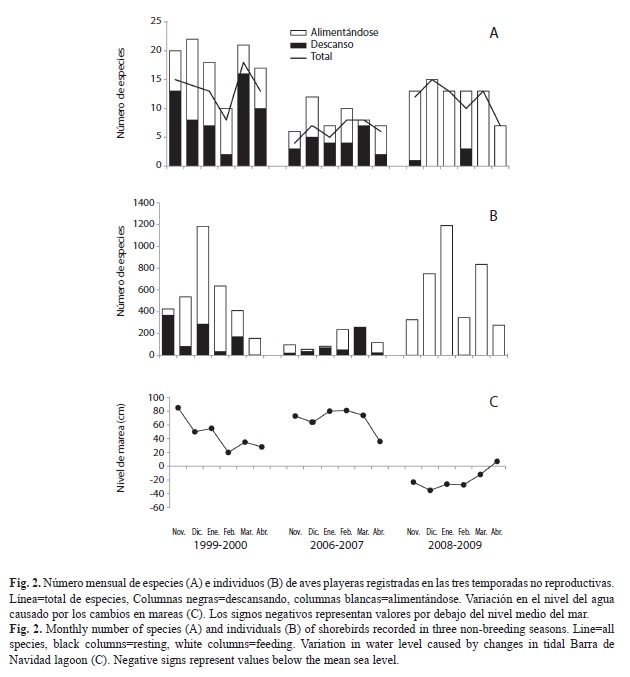
Distribución temporal: El número total de especies observado mensualmente presentó diferencias significativas entre las tres temporadas (Kruskal-Wallis, p=0.003). El mayor número de especies se registró en noviembre, diciembre y marzo en la temporada 1999-2000 y 2008-2009. En la temporada de 2006-2007, el número de especies fue relativamente bajo y no mostró un patrón definido. En cuanto a la actividad del número de especies, en la primera temporada (1999-2000) se registró una mayor abundancia de especies en descanso en noviembre, marzo y abril y más especies alimentándose de diciembre, enero y febrero. En 2006-2007 no hubo un patrón definido y el número de especies alimentándose y en descanso fue muy similar. A diferencia de las dos primeras temporadas, en 2008-2009 la mayoría de las especies se encontraban alimentándose (Fig. 2A).
El número total de individuos también presentó diferencias significativas entre las tres temporadas (Kruskal-Wallis, p=0.007). En 1999-2000 se registró un mayor número en enero, con 1 184 individuos, mientras que en 2006-2007 se registró un ligero incremento en febrero (235) y marzo (257). Para 2008-2009 hubo dos incrementos importantes, uno en enero con 1 191 individuos y otro en marzo con 834 individuos. El mayor número de individuos se observó principalmente alimentándose, esto fue más evidente en las temporadas 1999-2000 y 2008-2009 (Fig. 2B).
Al comparar el número de especies con el nivel de marea no se observó un correlación significativa entre ellos (Spearman, p>0.05), pero si hubo una correlación negativa significativa con el número de individuos (Spearman, p<0.05), es decir que el número de individuos se incrementó al disminuir el nivel del agua. El nivel de marea fue más alto en la primera y segunda temporada (10-88 y 36-81cm, respectivamente), mientras que en la tercera temporada se registraron los niveles más bajos (7-35cm) (Fig. 2C), coincidiendo con la mayor abundancia de especies e individuos que se encontraban alimentándose (Fig. 2A,B).
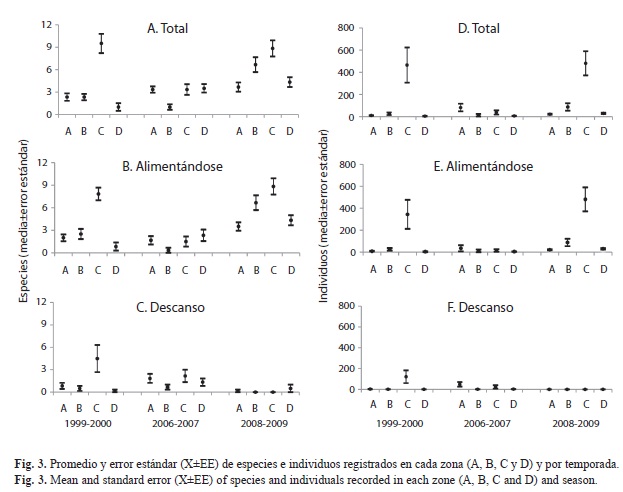
Distribución espacial: El promedio de especies registrado en las cuatro zonas (A, B, C y D) presentó diferencias significativas en cada una de la temporadas: 1999-2000 (Kruskal- Wallis, p=0.0006), 2006-2007 (Kruskal-Wallis, p=0.011) y 2008-2009 (Kruskal-Wallis, 10.93, p=0.012). En las tres temporadas se registró el promedio más alto en la zona C (Media±Error Estándar: 9.5±1.3, 3.3±0.7 y 8.83±1.1, respectivamente) (Fig. 3A).
La actividad de las especies (alimentándose o en descansando) no presentó diferencias significativas en las dos primeras temporadas (1999-2000, Mann-Whitney, p>0.05 y 2006-
2007, Mann-Whitney, p>0.05), mientras que en la tercera temporada hubo diferencias significativas entre las dos actividades (Mann- Whitney, p<0.05), siendo mayor el número de especies que se encontraban alimentándose (Fig. 3B,C).
El promedio de individuos también presentó diferencias significativas en las tres temporadas: en 1999-2000 (Kruskal-Wallis, p=0.0011), 2006-2007 (Kruskal-Wallis, p=0.007) y 2008-2009 (Kruskal-Wallis, p=0.0019). En la primera y tercera temporada la zona C registró los promedios más altos, con 465±158.6 y 481.3±109.06, respectivamente (Fig. 3D). En las comparaciones entre actividad, no se observaron diferencias significativas entre las aves alimentándose y en descanso en 1999-2000 (Mann-Whitney, p>0.05) y 2006-2007 (Mann-Whitney, p>0.05). Sin embargo, en la temporada 2008-2009 hubo diferencias entre las dos actividades (MannWhitney, p<0.05), siendo mayor el número de individuos que se encontraba alimentándose (Fig. 3E y 3F).
Discusión
Las 19 especies registradas en la laguna Barra de Navidad representan el 35.8% de las especies de aves playeras reportadas para México y el 64.1% de las aves playeras reportadas para la costa de Jalisco. Estos números son muy similares a los observados en otros humedales del pacífico central mexicano, tal es el caso de la laguna Agua Dulce (25 especies, 2 277 aves), Ermitaño (19 especies, 1 298 aves) (Hernández-Vázquez 2005), Majahuas (22 especies, 2707 aves), El Chorro (22 especies, 3 471 aves) (Hernández- Vázquez & Mellink 2001). Sin embargo, estas abundancias son inferiores a las observadas en humedales al norte del pacífico mexicano, como Guerrero Negro-Ojo de Liebre, Baja California (268 000 individuos), Marismas Nacionales, Nayarit (206 000 individuos), estero Río Colorado, Sonora (164 000 individuos) (SEMARNAT 2008).
Aunque la laguna Barra de Navidad, al igual que otros humedales cercanos, se caracterizan por ser relativamente pequeños y por concentrar abundancias bajas de aves playeras, es importante mencionar que estos humedales son usados como sitios de parada por las aves playeras migratorias, donde se alimentan y descansan para continuar su vuelo al norte y sur del continente (Hernández-Vázquez et al. 2010).
El número mensual de especies e individuos, así como la actividad de las aves playeras presentaron un patrón diferente entre temporadas. En 1999-2000 el número de especies e individuos fue relativamente alto en los meses que se registraron mareas altas (noviembre, diciembre y enero). En estas condiciones de marea se esperaría que el número de aves fuera menor, debido a que los hábitats de alimentación y descanso se encuentran cubiertos por el agua. Sin embargo, durante los censos se pudo observar que aparentemente las aves no se desplazaron a otros humedales, sino a una pequeña área rodeada por manglares ubicada en el margen noreste de la laguna en la zona C y D. En marea alta esta área es ligeramente inundada, creando un hábitat somero y protegido donde las aves se concentraron para alimentarse y descansar durante marea alta. En la segunda temporada (2006-2007) también se registraron mareas altas, sin embargo, el número de especies e individuos fue mucho menor. Aunque en el presente estudio no fue evaluado el efecto antropogénico en la distribución de las aves, una posible causa de estas diferencias puede deberse al azolvamiento y destrucción del mangle que rodean las pequeñas áreas someras y protegidas por el mangle de la zona C y D. La alteración de estas áreas causó el bloqueo de agua durante marea alta y muy probablemente la disminución de invertebrados que forman parte de la dieta de las aves playeras. Aparentemente la destrucción de este hábitat obligó a las aves a desplazarse a otros humedales cercanos o playas donde el alimento es más accesible. En la tercera temporada (2008-2009) se registraron las mareas más bajas y consecuentemente un mayor número de individuos que se alimentaron en los sustratos blandos expuestos.
En el nivel de marea más bajo registrado (-35cm bajo el nivel medio del mar) quedó expuesta una mayor área de sustratos blandos: en las zona A fueron 2.8 hectáreas de sustratos arenosos (La Playa y El Remanso), en la zona B con 9.3 hectáreas de sustratos arenosos (Colimilla), zona C con 80.6 hectáreas de sustratos lodosos (La Soledad) y zona D sólo 2.6 hectáreas de áreas arenosas (El Conchero). Esta disponibilidad de hábitat de forma temporal es un factor importante que influyó en la distribución y actividad de las aves playeras.
La zona C presentó una mayor área de sustratos lodoso (La Soledad) y consecuentemente el mayor número de especies e individuos, esto fue más evidente en la primera y tercera temporada. En esta zona el arroyo Seco y un canal del Río Marabasco descargan sus aguas en la laguna, acarreando una gran cantidad de sedimento arcilloso que son depositados en esta zona. Este tipo de sustrato es hábitat de una gran variedad de invertebrados, como poliquetos, moluscos y crustáceos que forman parte importante en la dieta de las aves playeras (Skagen & Oman 1996, Butler et al. 2001). Este incremento en la densidad de los invertebrados, así como la disponibilidad de los mismos durante la marea baja son factores que influyen a escala local en la distribución y abundancia de las aves playeras (Warnock et al.2002, Austin & Rehflsch 2003).
La baja abundancia de aves en las zonas A, B y D se debe a la reducción de áreas con sustratos blandos expuestas en marea baja. Aunado a esto, gran parte de estas zonas se encuentran rodeadas por asentamientos urbanos y turísticos, siendo sometidas a una constante actividad humana, como tráfico de embarcaciones, pesca y actividades turísticas. Ya ha sido documentado en varios trabajos que estas actividades tienen un efecto negativo en la distribución y abundancia de las aves playeras (Burger 1986, Burger et al. 1995, Burger & Gochfeld 1991a,b 1998, Cursach & Rau 2008). Estas actividades reducen la eficiencia de alimentación y las oportunidades de descanso de las aves, por lo que el costo energético es mayor al escapar frecuentemente de las actividades humanas (Burger 1986), obligando a las aves a buscar áreas donde el disturbio sea menor, y donde la disponibilidad de alimento sea mayor, como se pudo observar en la zona C.
En general, la laguna Barra de Navidad presentó abundancias de aves playeras similares a otros humedales del pacífico central mexicano, pero inferiores a humedales de mayor tamaño ubicados en norte del país. Sin embargo, la variabilidad en el nivel de agua causado por la marea dejó varias áreas expuestas temporalmente y fueron aprovechadas por las especies migratorias y residentes para alimentarse y descansar. Las mayores abundancias de aves registradas en la primera y tercera temporada se observaron en sustratos arcillosos ubicados en la zona C, donde las aves se concentraron para alimentarse durante marea baja. Esta información es relevante para la protección de este grupo de aves, partiendo de la premisa de que, si no se puede proteger toda la laguna si se puede dedicar un mayor esfuerzo de protección en las zonas de mayor concentración de aves, como puede ser la zona C. Lo anterior toma mayor importancia al considerar que gran parte de la laguna Barra de Navidad se encuentra rodeada por infraestructura urbana y turística, dada la tendencia del crecimiento urbano de los últimos años es probable que se extiendan a otras áreas de la laguna que aún conservan condiciones favorables para las aves. Aunado a lo anterior, actualmente existe un proyecto en el que se pretende dragar toda la laguna (seis metros de profundidad) con el fin de facilitar la navegación dentro de ella. Es claro que esta actividad acabará con los sustratos blandos expuestos en marea baja, disminuyendo así la disponibilidad del alimento, y consecuentemente la abundancia y distribución de este grupo de aves. Con los resultados generados en este trabajo, además de proveer de información básica para identificar las zonas críticas de la laguna que no deben ser modificadas, servirá de comparación con estudios futuros realizados después del dragado y evaluar el posible daño causado por esta actividad.
Agradecimientos
A la Federación Regional de Sociedades Cooperativas de la Industria Pesquera del Estado de Jalisco, F.C.L por el apoyo logístico requerido durante el trabajo de campo. A la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad “CONABIO” (Convenio Núm. FB425/L292/97), el Canadian Wildlife Service/Latin American Program, Centro Universitario de la Costa Sur, Universidad de Guadalajara y al Programa de Mejoramiento del Profesorado (PROMEP: 103.5/07/2449) por el financiamiento en sus diferentes fases. A los revisores anónimos por sus acertados cometarios que ayudaron a mejorar el manuscrito.
Referencias
American Ornithologists’Union (A.O.U.). 2011. Check-list of North American Birds. http://www.aou.org/). The American Ornithologists’ Union, Washington, D.C. EEUU. (Consultado: 20 octubre 2011). [ Links ]
Arriaga-Cabrera, L., E. Vázquez., J.G. Cano, R.J. Rosenberg, E.M. López & V.A. Sierra (coordinadores). 1998. Regiones marinas prioritarias de México. Comisión Nacional para el Conocimiento y uso de la Biodiversidad. México. (Consultado: 31 octubre 2011, http://www.conabio.gob.mx/conocimiento/regionalizacion/doctos/marinas.html). [ Links ]
Arriaga-Cabrera, L., V. Aguilar & J. Alcocer. 2002. Aguas continentales y diversidad biológica de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. (Consultado: 31 octubre 211, http://www.conabio.gob.mx/conocimiento/regionalizacion/doctos/hidrologicas.html). [ Links ]
Austin, G.E. & M. Rehfisch. 2003. The likely impact of sea level rise on waders (Charadrii) wintering on estuaries. J. Nat. Conservat. 11: 43-58. [ Links ]
Burger, J. 1986. The effect of human activity on shorebirds in two coastal bays in northeastern Unites States. Environ. Conservat. J. 13: 123-130. [ Links ]
Burger, J. & M. Gochfeld. 1991a. Human distance and birds: tolerance and response distances of resident and migrant species in India. Environ. Conservat. J. 18: 158-165. [ Links ]
Burger, J. & M. Gochfeld. 1991b. Human Activity Influence and Diurnal and Nocturnal Foraging of Sanderlings (Calidris alba). Condor 93: 259-265. [ Links ]
Burger, J., M. Gochfeld & L.J. Niles. 1995. “Ecotourism and birds in coastal New Jerset: Contasting responses of birds, tourists, and managers”. Environ. Conservat. J. 22: 56-64. [ Links ]
Burger, J. & M. Gochfeld. 1998. Effects of ecotourists on bird behavior at Loxahatchee National Wildlife Refuge, Florida. Environ. Conservat. J. 25: 13-21. [ Links ]
Butler, R.W., N.C. Davison & R.I.G. Morrison. 2001. Global-scala shorebirds distribution in relation to productivity of near-shore ocean water. Colon. Waterbird 24: 224-232. [ Links ]
Cupul-Magaña, F. 2000a. Aves acuáticas del estero El Salado, Puerto Vallarta, Jalisco. Huitzil 1: 3-8. [ Links ]
Cupul-Magaña, F. 2000b. Notas sobre la avifauna acuática de las islas y los humedales costeros de Bahía de Banderas, Jalisco-Nayarit, México. Mexicoa 2: 85-87. [ Links ]
Cursach, J.A. & J.U. Rau. 2008. Influencia de las perturbaciones humanas sobre la diversidad del ensamble de aves costeras en el seno de Reloncoví, Sur de Chile. Bol. Chil. Ornitol. 14: 92-97. [ Links ]
Goss-Custard, J.D., R.A. Jenyon, R.E. Jones, P.E. Newbery & R.L.E. Williams. 1977. The ecology of The Wash II. Seasonal variation in the feeding conditions of wading birds (Charadrii). J. Appl. Ecol. 14: 701-719. [ Links ]
Hernández-Vázquez, S. 2000. Avifauna acuática del estero La Manzanilla, Jalisco, México. Act. Zool. Mex. 80: 143-153. [ Links ]
Hernández-Vázquez, S. & E. Mellink. 2001. Coastal waterbirds of El Chorro and Majahuas, México, during the non-breeding season, 1995-1996. Rev. Biol. Trop. 49: 357-365. [ Links ]
Hernández-Vázquez, S. 2005. Aves acuáticas de la laguna de Agua Dulce y estero El Ermitaño, Jalisco, México. Rev. Biol. Trop. 53: 229-238. [ Links ]
Hernández-Vázquez, S., R. Rodríguez-Estrella, J.H. Vega- Rivera, F. Hernández-Vázquez, J.A. Rojo-Vázquez & V.H. Galván-Piña. 2010. Estructura, dinámica y reproducción de las asociaciones de aves acuáticas de la costa de Jalisco, México, p. 11-27. In E. Godínez- Domínguez, C. Franco-Gordo, J.A. Rojo-Vázquez, F. Silva-Bátiz & G. González-Sansón (eds.). Ecosistemas marinos de la costa Sur de Jalisco y Colima. Universidad de Guadalajara. Guadalajara, México. [ Links ]
Myers, J.P., R.I.G. Morrison, P.Z. Antas, B.A. Harrington, T.E. Lovejoy, M. Sallaberry, S.L. Senner & A. Tarak. 1987. Conservation strategy for migratory species. Am. Sci. 75: 19-26. [ Links ]
National Geographic Society. 1987. Field Guide to the Birds of North America. Washington D.C., EEUU. [ Links ]
National Geographic Society. 2006. Field Guide to the Birds of North America. Washington D.C., EEUU. [ Links ]
O’Brien, M., R. Crossley & K. Karlson. 2006. The Shorebirds guide. Houghton Miffin, Nueva York, EEUU. [ Links ]
Ramsar. 2011. The list of wetlands of international importance. (Consultado: 22 octubre 2011, http://www.ramsar.org/pdf/sitelist.pdf). [ Links ]
SEMARNAT. 2008. Estrategia para la Conservación y Manejo de las Aves Playeras y su Hábitat en México. Gobierno Federal, México, D.F, México. [ Links ]
Silva-Bátiz, F.A., S. Hernández-Vázquez, A.J. Nené-Preciado & A.D. Vázquez-Lule. 2009. Caracterización del sitio de manglar Laguna Barra de Navidad, en Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). Sitios de manglar con relevancia biológica y con necesidades de rehabilitación ecológica. CONABIO, México, D.F. (Consultado: 5 julio 2011, http://www.conabio.gob.mx/conocimiento/manglares/doctos/caracterizacion/C11_Barra_de_Navidad_caracterizacion.pdf). [ Links ]
Skagen, S.K. & H.D. Oman. 1996. Dietary flexibility of shorebirds en the western hemisphere. Can. Field. Nat. 110: 419-444. [ Links ]
Warnock, N., C. Elphick & M.A. Rubega. 2002. Shorebirds in the marine environment, p. 581-615. In A. Schreiber & J. Burger (eds.). Biological of marine birds. CRC, Florida, EEUU. [ Links ]
Zar, J.H. 1974. Biostatistical Analysis. Prentice Hall, Nueva York, EEUU. [ Links ]
*Correspondencia:
Salvador Hernández: Departamento de Estudios para el Desarrollo Sustentable de Zona Costera. Universidad de Guadalajara. Gómez Farías No. 82, San Patricio-Melaque, Municipio de Cihuatlán, Jalisco. CP: 48980. México. sahernan@costera.melaque.udg
Sergio Serrano: Departamento de Estudios para el Desarrollo Sustentable de Zona Costera. Universidad de Guadalajara. Gómez Farías No. 82, San Patricio-Melaque, Municipio de Cihuatlán, Jalisco. CP: 48980. México. mx. seseguz@hotmail.com
Xóchitl A. Hernández: Departamento de Estudios para el Desarrollo Sustentable de Zona Costera. Universidad de Guadalajara. Gómez Farías No. 82, San Patricio-Melaque, Municipio de Cihuatlán, Jalisco. CP: 48980. México. xahg_serch@hotmail.com
María Isabel Robles: Presidencia Municipal, Juan M. Díaz No. 3, C.P. 48680, Ejutla, Jalisco. isa_2301@hotmail.com
1. Departamento de Estudios para el Desarrollo Sustentable de Zona Costera. Universidad de Guadalajara. Gómez Farías No. 82, San Patricio-Melaque, Municipio de Cihuatlán, Jalisco. CP: 48980. México; sahernan@costera.melaque.udg. mx, seseguz@hotmail.com, xahg_serch@hotmail.com
2. Presidencia Municipal, Juan M. Díaz No. 3, C.P. 48680, Ejutla, Jalisco; isa_2301@hotmail.com
Recibido 21-VII-2011. Corregido 09-III-2012. Aceptado 12-IV-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}