










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.60 n.3 San José Sep. 2012
Who is who in the understory: the contribution of resident and transitory groups of species to plant richness in forest assemblages
*Dirección para correspondencia
Abstract
The forest understory is made up of resident and transitory species and can be much richer than the canopy. With the purpose to describe the contribution of these groups to the woody understory, five Atlantic Forest fragments were selected and studied in Northeastern Brazil. In order to analyze the understory’s structure, the sample included woody individuals with circumference at breast height (CBH) smaller than 15cm and circumference at ground level (CGL) greater than 3cm, regardless of height. The recorded species were quantified and classified into functional stratification categories (resident and transitory), and the floristic similarity between the understory and the tree stratum was calculated. Species’ importance in the understory was analyzed by height and total natural regeneration classes based on a regeneration index. The understory was richer in species (median=63.8, SD=21.72, n=5 fragments) than the tree stratum (43.8, 18.14, 5), and the similarity between these components was relatively high (median=0.54, SD=0.09). The results also showed that the studied understory in the forest fragments was mainly composed by transitory species (median=67.01%, SD=3.76), that were well distributed among height classes and had the highest densities, which may favor their future presence in the canopy’s structure and composition. The typical understory species were grouped into two strata: the lower understory, made up of species that generally do not reach more than 4m in height (mostly species from families Piperaceae, Rubiaceae and Melastomataceae); and the upper understory, with intermediate heights between the lower understory and the canopy, but with average heights that were not higher than 10m (mainly of species from families Anonnaceae, Clusiaceae and Myrtaceae). These families’ richness was commonly used as an indicator of the vegetation’s successional stage; however, such results must be seen with caution as they show that these families co-occurred and were highly important in different strata. Studying the understory is fundamental because it represents a floristically rich stratum with a unique structure, which promotes the natural regeneration of the tree stratum.
Key words: height classes, floristic composition, density, natural regeneration, richness, functional groups.
Resumen
El sotobosque forestal está compuesto por especies residentes y transitorias. Con el objetivo de describir la contribución de esos grupos en el sotobosque leñoso, cinco fragmentos de Bosque Atlántico fueron seleccionados en el nordeste de Brasil. El muestreo incluyó individuos con circunferencia a la altura del pecho (CPA)<15cm y con circunferencia a la altura del suelo (CAS)≥3cm. Las especies fueron cuantificadas y clasificadas en residentes o transitorias y la similitud florística entre el sotobosque y el dosel fue calculada. El sotobosque se mostró más rico en especies (mediana=63.8, DS=21.72, n=5 fragmentos) que el dosel (43.8, 18.14, 5) y la similitud entre esos componentes fue relativamente alta (0.54, 0.09). El sotobosque está compuesto principalmente de especies transitorias (mediana=67.01%, DS=3.76) las cuales están bien distribuidas entre las clases de altura y presentan las mayoresdensidades. Las especies residentes fueron agrupadas en dos estratos: el sotobosque bajo (<4m de altura, la mayoría Piperaceae, Rubiaceae y Melastomataceae); y el sotobosque superior (altura media <10m, principalmente Anonnaceae, Clusiaceae y Myrtaceae). El estudio del sotobosque es fundamental porque es un estrato florísticamente rico con un estructura única, lo cual promueve la regeneración natural del estrato arbóreo.
Palabras clave: clases de altura, composición florística, densidad, regeneración natural, riqueza de especies, Bosque Atlántico, grupo funcional.
Several life forms coexist in the forest understory, which contributes to high species richness (Gentry & Dodson 1987, Schnitzer & Carson 2000). There are two main approaches to view the regeneration dynamics of this stratum: either as a physiognomic component defined by a maximum height or diameter, including young trees, seedlings, saplings, and shrubs, regardless of their potential to occupy the forest vertically (for example, Campos & Landgraf 2001, Lima-Filho et al. 2002, Rayol et al. 2006, Silva et al. 2007, Marangon et al. 2008, Gomes et al. 2009, Lü et al. 2010, Sansevero et al. 2011); or as functional groups of plants that occupy the lowest level of the forest and might include resident or transitory species (Tabarelli & Mantovani 1999, Harms et al. 2004, Araújo et al. 2006, Santos et al. 2008, Aquino & Barbosa 2009, Onofre et al. 2010).
The resident group of species, which may be called the typical understory, is made up of herbaceous and sprawling species, as well as shrubs and small trees that remain in this stratum throughout their life cycle. Conversely, the transitory group - also called the natural regeneration group - includes species that have the potential to grow and reach the forest canopy, such as trees and vines (Gilliam et al. 1994, 1995). The transitory group is generally made up of tree individuals with dendrometric values lower than those defined for the tree stratum (normally diameter at breast height, DBH>5cm) (Finol 1971). Since these two groups concurrently occur (Gilliam et al. 1995), the understory’s composition may include much more species than the canopy (Galeano et al. 1998, Lü et al. 2010). Future canopy composition, in turn, is dependent on the density of transitory species in the understory, as well as on the recruitment of these species from the lower to the highest height classes (Clark & Clark 1992, Volpato 1994, Kobe 1999). Species that occur in all of the height classes have greatest survival chances, as natural mortality is greater in the lower height classes (Volpato 1994, Felfili et al. 2000), caused by physical damage, leaf litter, vertebrates (Scariot 2000, Ickes et al. 2001, Santos & Válio 2002) or by biological damage such as predation and parasitism (Cadenasso & Picket 2000).
The physiognomic and functional analyses, when added to the analysis of the understory’s vertical structure, help understanding the forest’s future development and composition (Gama et al. 2003) and allow us to: better estimate the richness and state of conservation (Richards 1996), diagnose the dynamics of natural forest fragments (Finol 1971), measure responses to soil and climatic variations and to environmental stress (Harms et al. 2004), plan management actions, forestry practices and vegetation restoration activities (Emborg 1998, Mattei & Longhi 2001, Gama et al. 2003, Silva et al. 2007, Sansevero et al. 2011).
In a mature tropical forest, the floristic composition of the canopy and understory is expected to be distinct from each other (Jardim & Hosokawa 1986) due to the low height of herbs and shrubs, which contributes to an increase of richness in the understory, and also to the dynamics of tree species in different successional groups. As shown by Nascimento (2010) in a 12-year secondary area, regenerated after clearcutting, the similarity between woody species in the canopy and in the understory was of 76% and diminished to 48% with successional maturity, in 20-year secondary forests. For a more mature stage, Alves & Metzger (2006) recorded 30% similarity in a secondary 80-year forest. Such low similarity in intermediate regeneration stages is due to the concentration of secondary species in the understory and pioneer species in the canopy, since the pioneer species grow faster than the secondary species until they reach maturity (Válio 2003, Bohlman & O’Brien 2006). Similarity may start increasing again during a more advanced successional stage, as the secondary species that once occupied the regeneration stratum become part of the tree stratum. This pattern was found by Silva et al. (2008) who recorded an increase in the similarity between the canopy and the understory from a more disturbed area (0.45; edge) to a more conserved one (0.59; forest interior) in a 300ha mature Atlantic forest fragment.
In fragmented landscapes, analyzing the understory’s structure and identifying its functional components has been crucial to diagnose the effects of disturbances (Martins & Rodrigues 2002, Ceccon et al. 2006), fragmentation and edge effects (Benitez-Malvido 1998, Laurance et al. 2007, Bouroncle & Finegan 2011). In the Brazilian Atlantic Forest, where most areas have been substituted for cultivated land, it is estimated that only 11.6% of the original cover remains (Ribeiro et al. 2009). Yet, despite its status as a world conservation hotspot (Mittermeier et al. 2005), and the fact that many plants spend all or a considerable part of their lives in the understory (Harms et al. 2004), few studies have analyzed the understory vertical structure (e.g. Silva et al. 2007, Marangon et al. 2008), described who is who in this stratum regarding plant habit (Souza et al. 2009) or successional category (Tabarelli & Mantovani 1999, Aquino & Barbosa 2009,Onofre et al. 2010, Sansevero et al. 2011) or compared understory composition and structure among forests (Gomes et al. 2009).
Only 4.6% of the original vegetation remains in the extreme North of the Atlantic Forest’s distribution area (Lima 1998), in the form of small, irregular fragments, most less than 500ha in size (Trindade et al. 2008). Such forests are located mainly at the bottom of deep valleys - areas where small springs and watercourses can be found - and have closed canopies. They are, thus, riparian forests (Naiman et al. 2005) made up of assemblages that border watercourses in areas where the interfluvial vegetation is also forest (Metzger et al. 1997). In addition to the typically riparian species, species from the upper forests can also be found in these habitats, since the riparian vegetation serves as an important source of diaspores for forest remnants’ natural regeneration and recolonization processes (Triquet et al. 1990).
With the purpose of describing the physiognomic and functional structure of the understory in Atlantic Forest fragments, five differently sized remnants were selected from a highly fragmented landscape. The study had five goals: 1) to uncover species richness and composition; 2) to discover how many and which canopy species are found in the understory; 3) to functionally classify species as resident or transitory; 4) to investigate the existence of a pattern in the understory’s structure among the different fragments; and 5) to discover the present and potential regenerative capacity of the remnants studied.
Material and methods
Study area: Five fragments were studied, all from the extreme North of the state of Pernambuco, Northeastern Brazil (07º41’05” - 07º54’17” S and 34º54’17” - 35º05’07” W), at 20-50m in elevation, within properties of the São José sugarcane plantation/processing plant. For the purposes of this study, the fragments were named: F1 (Piedade Forest, 305.78ha), F2 (Macacos Forest, 356.22ha), F3 (Zambana Forest, 387.85ha), F4 (Vespas Forest, 13.80ha), and F5 (Pezinho Forest, 29.89ha). The São José plant covers approximately 240km2; the landscape is dominated by sugarcane monoculture. The remnant forest vegetation (24% of the total area) is inserted within the Atlantic Forest domain called Dense Ombrophilous Lowland Forest (IBGE 1992) and covers 65.7km2 of this landscape, divided among 106 fragments (Trindade et al. 2008). Following Köppen’s classification, climate is of type As’ (tropical wet), with an average precipitation of 1 689mm and rain concentrated from April-August; the average temperature is 25.1ºC (data from the São José sugarcane processing plant for the period 1998-2006). The fragments are located on the Barreiras geological formation, which dates back to the Plio-Pleistocene and consists of unconsolidated sandy-clay sediments of continental origin. The terrain is made up of flattened tableland cut by deep, narrow valleys with sides that slope at 30% declivity or higher (CPRH 2003). The studied remnants house watercourses between one and three meters wide that are part of the Botafogo River water basin, which allows them to be classified as riparian forest (sensu Naiman et al. 2005).
Composition and structure data sampling: In order to analyze the understory, 20 non-contiguous 5x5m parcels were installed in each fragment, at the apex of larger 10x10 plots that had already been used for a study on trees available from Lins-e-Silva (2010), as part of the databank of the Fragments Project. The parcels were arranged to guarantee that both forest edge and interior environments be sampled (as well as both sides of the watercourse), at a minimum distance of 5m between the units.
To sample to tree stratum, we included all of the trees with CBH (circumference at breast height) greater or equal to 15cm. In the understory, we included the woody individuals with CBH lesser than 15cm and circumference at ground level (CGL) greater than 3cm, regardless of height. All of the individuals included in the tree stratum or understory had their CBH or CGL taken with a measuring tape and their height estimated with the aid of an 8m graduated ruler. Additionally, a fertile or sterile sample of each species’ branches was collected. Fieldwork was undertaken from 2007-2009. The data (CGL, CBH, height, botanical family and species) was organized into a database using the Mata Nativa 2 software (Souza et al. 2006).
The botanical material collected was added to the Herbarium Geraldo Mariz (UFP) of the Federal University of Pernambuco/Usina São José Collection. When possible, samples were identified up to the species level with the aid of specific literature, comparison with material from the UFP, PEUFR and IPA herbaria that had been previously identified by specialists. When necessary, the material was sent to specialists for identification and/or confirmation. All of the species collected were listed alphabetically according to family; the APG III (2009) classification system was used. Species names were updated regarding synonymies based on the Missouri Botanical Garden website (Tropicos 2010); author names and/or abbreviations followed the International Plant Names Index (IPNI 2010).
Analysis of data structure and composition: The woody flora of the studied understory was classified into functional stratification categories: 1) transitory species (TR), represented by young individuals of the trees that make up the forest’s canopy; and 2) typical understory species, i.e., resident species such as shrubs and small trees that develop under the canopy. The typical understory species were also divided into: a) lower understory (LU), which included species that make up the lowest forest stratum and do not surpass 5m in height, on average; and b) upper understory (UU), which included sciophilous species that are part of the intermediate stratum and do not reach the canopy.
The subdivision of typical species into the low understory and upper understory categories was based on the works of Vilela et al. (1995), Oliveira & Amaral (2005), Gomes et al. (2009), Souza et al. (2009) and Onofre et al. (2010). Upper limit was fixed at 10m based on the average height of the maximum heights of each species recorded in 370 plots studied by the Fragments Project, undergoing at São José Sugarcane Property since 2003. The number and percentage of species in each functional stratification category (SC) was determined for each fragment. As density varied amongst samples, richness of canopy and understory strata where estimated through rarefaction (100 permutations) using the software EstimateS 8.2 (Cowel 2009), for the minimal density value recorded in each stratum (160 trees; 531 understory plants). Observed and estimated values were then compared using T test. Homogeneity of species and individuals distribution was compared among the SCs using the G Test, and the floristic similarity between the understory and the tree stratum was calculated using Sørensen’s similarity index, with the purpose of defining which species recorded in the understory have regeneration potential for the overlying tree stratum. For test descriptions, see Magurran (2004).
Studies on natural regeneration usually stratify vegetation into height classes. In this study we adjusted the classification applied by Oliveira-Filho et al. (1994) and Marangon et al. (2008). Marangon et al. (2008) used the minimum 1m height limit and justified that individuals with such a height have better morphological characteristics and, thus, allow for a more reliable identification. In this study, we also included plants that were less than 1m tall, as Oliveira-Filho et al. (1994) have stated that this makes it possible to sample more individuals. Individuals were grouped into the following height classes: class 1-height<1m; class 2-height≥1m and <2m; class 3-height≥2m and <3m; and class 4-height ≥3m with CBH less than 15cm.
Species importance in the understory was analyzed based on the Natural Regeneration Index by height class and Total Natural Regeneration, suggested by Finol (1971) and modified by Volpato (1994). The natural regeneration index includes parameters related to horizontal and vertical structure (density, frequency and distribution of individuals by height class). Over the last years the natural regeneration index has been used with the purpose of carrying out more complete diagnoses on the dynamics and development of forest succession. For example, Armesto & Martínez (1978) used the index by Finol for describing Mediterranean vegetation in Chile, whereas Silva et al. (2007), Marangon et al. (2008) and Santos et al. (2008) used Volpato’s modified version for describing natural regeneration in Brazilian forests. This latter incorporates a more detailed analysis per height class when calculating the index, which considers the relative density of the species in each class.
In order to calculate the Natural Regeneration Index, absolute and relative density and frequency parameters were estimated by height class for each species of each fragment. The natural regeneration estimate by height class (NRC) was made based on these parameters, according to Volpato’s (1994) equation: NRCij=(RDij+RFij)/2 where NRCij is the natural regeneration estimate of the ith species of the jth plant size class, as a percentage, RDij is the relative density for the ith species of the jth natural regeneration size class, RFij is the relative frequency for the ith species of the jth natural regeneration size class, i is the 1, 2, 3... pth species sampled, and j is the 1, 2, 3, and 4 classes.
The next step was to calculate the Total Natural Regeneration estimate (TNR) for each species by adding the natural regeneration indexes of each class according to the formula by Volpato (1994): TNRi=ΣNRCij, where TNRi is the total natural regeneration estimate of the ith species, NRCij is the natural regeneration estimate of the ith species of the jth plant size class, i is the 1, 2, 3... pth species sampled, and j is the 1, 2, 3 and 4 classes.
Results
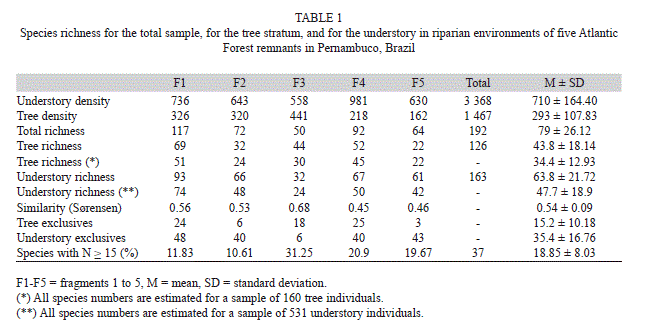
Species richness and composition: A total of 163 species distributed among 44 families were sampled from the understory of the five fragments analyzed (fragment median=63.8 species, SD=21.72, n=5), varying from 32 species in F3 to 93 species in F1 (Table 1). One hundred twenty-six species of 44 families were sampled from the tree stratum. The number of species in this stratum varied from 32 in F2 to 69 in F1 (43.8, 18.14, 6). When richness was estimated using rarefaction, mean richness calculated values were 34.4±12.93 (median±SD) from the canopy and 47.69±18.09 from the understory, not significantly different from observed values in both strata. All of the fragments except F3 were richer, although not significantly, in the understory. An average of 35% (SD=16.76) of the species occurred exclusively in the understory, while 15.2% (10.18) of the species only occurred within the tree stratum of the riparian forests. Of the total, 75.4% of the species found in the tree stratum were present in the understory. The similarity between the tree and understory strata averaged 0.54 (0.09) (Table 1). Fragment F3 had the highest similarity (0.68), while fragments F4 and F5 were the least similar (0.46 and 0.45, respectively).
Families Myrtaceae (17 species/227 individuals), Melastomataceae (13/327), Rubiaceae (10/247), Fabaceae-Mimosoideae (10/234), Annonaceae (9/461) and Clusiaceae (7/266) had the highest species richness and added up to 40.50% of the understory’s total species. The lower understory was made up mainly of species from families Rubiaceae and Melastomataceae, which totalled 52.17% of the species and 24.35% of individuals in this category, while species from families Annonaceae, Clusiaceae and Myrtaceae predominated in the upper understory (66.66% of the species and 56.95% of individuals).
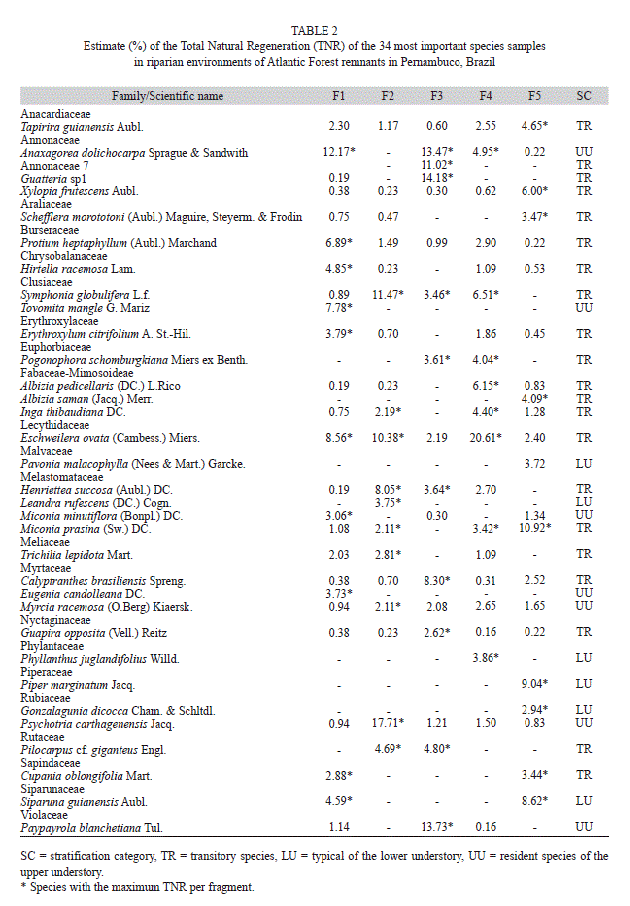
In addition to the families cited above, despite its lower richness, the family Lecythidaceae (3 species/447 individuals) was high in density. Anaxagorea dolichocarpa Sprague & Sandwith (Annonaceae), Eschweilera ovata (Cambess.) Miers. (Lecythidaceae) and Symphonia globulifera L.f. (Clusiaceae) stood out due to the number of individuals (24.10% of the total) and for being among the ten species with the highest natural regeneration indexes (TNR) in three of the five remnants studied.
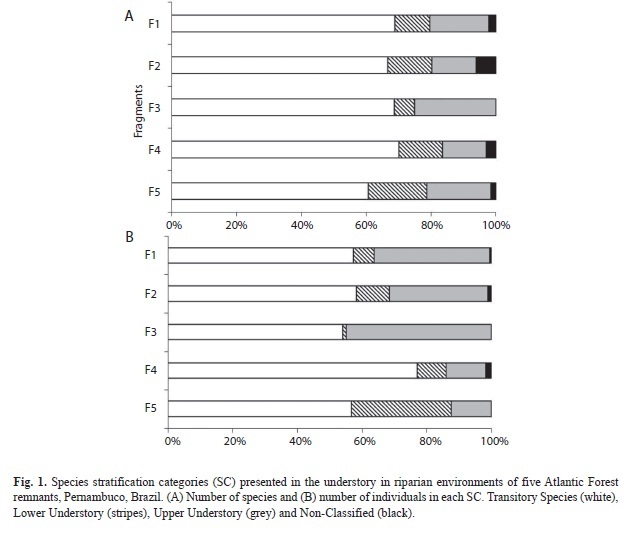
Species classification into stratification categories: The percentage of transitory species was greater than that of typical understory species (Fig. 1A). An average of 67.01% (SD=3.76) transitory species were recorded, while lower and upper understory species added up to 12.42% (4.33) and 18% (4.79) of the total species sampled, respectively. Between fragments, differences in species distribution into stratification categories were only detected for F3, which was different from fragments F2 (G=14.63, p<0.05), F4 (G=10.37, p<0.05) and F5 (G=9.38, p<0.05) because it had the smallest percentage of lower understory species (6.25%) and the greatest percentage of upper understory species (25%).
Transitory species averaged 60.73% (9.26) of the understory sampled individuals (Fig. 1B). Conversely, the lower and upper understory species represented 11.52% (11.33) and 27.06% (14.50) of the individuals, respectively. Fragment F5 differed from the others because it had a greater percentage of individuals in the lower understory (30.79%) and a smaller percentage of individuals in the upper understory (12.22%) in relation to fragments F1 (G=29.30, p<0.05), F2 (G=19.50, p<0.05) and F3 (G=54.82, p<0.05). Fragment F4 was different due to its higher percentage transitory species (76.06%) in comparison to the other fragments. In this remnant, the upper understory had the lowest value (12.23%) and was different from fragments F1 (G=16.10, p<0.05), F2 (G=10.74, p<0.05) and F3 (G=33.13, p<0.05). Significant differences were also found between fragments F2 and F3 (G=13.06, p<0.05), which showed opposite trends in relation to the percentage of resident species: F3’s lower understory had a smaller percentage (1.08%), while the percentage of upper understory was larger (44.80%).
Among the 163 species recorded in the five fragments’ understory, looking at the 10 with highest regeneration indexes (TNR) in each fragment, 34 species are listed (Table 2). These species include the most important in at least one fragment, in which they added up to 10.75-31.25% of the total natural regeneration. Transitory species were the majority, with 21 recorded species (61.76%), while the lower and upper understory had six (17.65%) and seven species (20.59%), respectively.
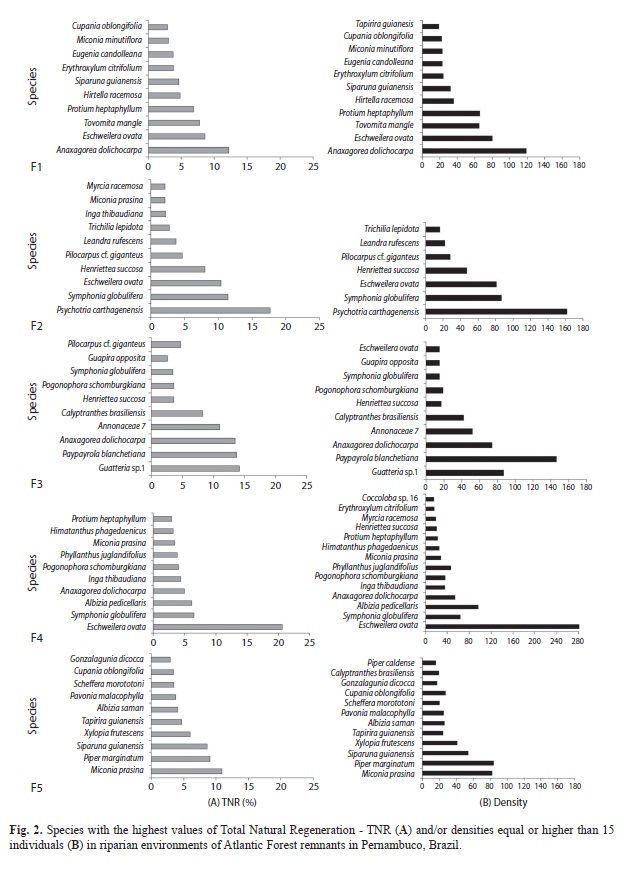
Regenerative capacity and vertical structure of the understory: Most species with high total natural regeneration indices (TNR) were also well distributed among height classes and had the highest densities (Fig. 2). Only 37 species - less than 21% of the total richness - had densities equal to or higher than 15 individuals in the total sample (Table 1); the highest percentage was recorded for F3, which stood out for housing 31.25% of the species with 15 or more individuals. Species such as Inga thibaudiana DC., Myrcia racemosa (O. Berg) Kiaersk., Miconia prasina (Sw.) DC. (F2) and Pilocarpus cf. giganteus Engl. (F3) all had less than 15 individuals each. Nevertheless, they were among those with the highest TNR, as they were recorded in all of the height classes (the exceptions were M. prasina and P. cf. giganteus, which were found in three classes, yet had high relative frequencies in relation to the others). Tapirira guianensis Aubl. (F1), E. ovata (F3), Henriettea succosa (Aubl.) DC., M. racemosa, Erythroxylum citrifolium A. St.-Hil.,Coccoloba sp16 (F4), Calyptranthes brasiliensis Spreng. and Piper caldense C.DC. (F5), although not being among the species with the highest regeneration, represent the regeneration potential of the fragment in which they occur, as they stand out in number of individuals. These species had low relative frequency and - with the exception of T. guianensis and M. racemosa - were absent from one of the height classes.
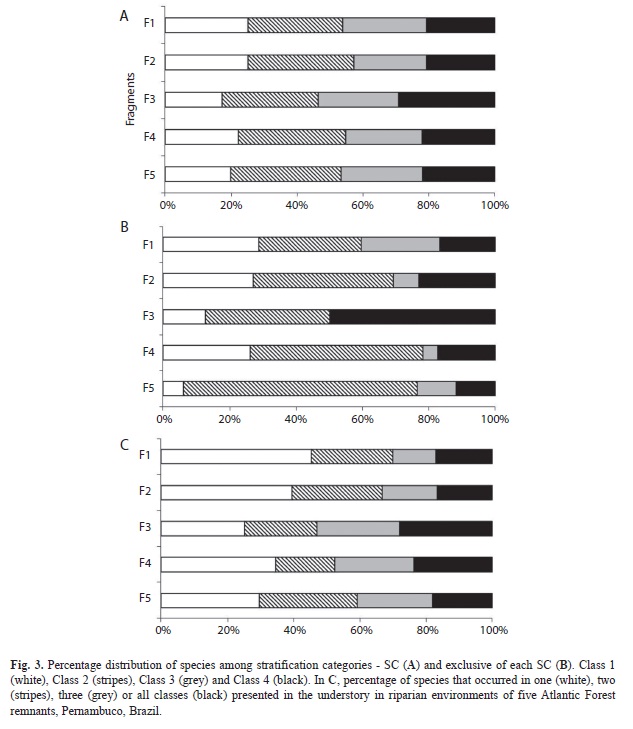
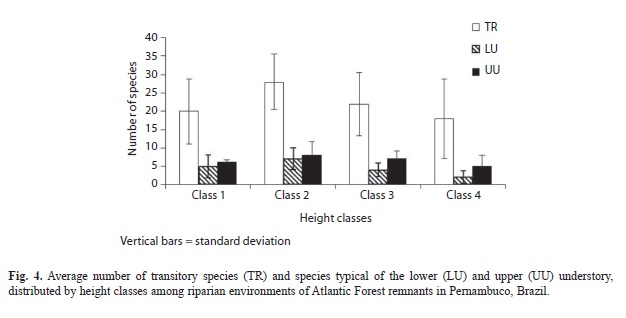
The percentages of species with densities of 15 or more individuals had the following distribution among functional stratification categories (SC): 23 transitory species (62.16%); six lower understory species (16.22%), and seven upper understory species (18.92%). Species’ distribution between the different height classes was similar among the fragments. The second regeneration class encompassed the largest number of species (Fig. 3A). The transitory species were the best represented in all of the classes, followed by those from the upper and lower understory (Fig. 4). The average number of typical lower and upper understory species was similar for classes 1 and 2.
In all of the forest remnants, the species that occurred in only one class were concentrated in the second height class (except for F3, where most occurred solely in class 4) (Fig. 3B). The distribution of transitory and typical understory species exclusively in one height class revealed that these species do not generally occur in classes 3 (2-3m height) and 4 (height >3m and CBH<15cm).
Few species occurred in all of the height classes - percentages of such species varied from 16.67% in F2 to 28.13% in F3 (Fig. 3C). Most species only occurred in one class, with percentages varying from 25% in F3 to 45.16% in F1. Transitory species obtained the greatest averages in all of the situations, while the average number of species in the lower and upper understory varied from 1-3.
Discussion
Species richness and composition: In this study, the understory generally contributed to an enhancement on plant richness in the forest fragments. This confirms the pattern that is normally recorded in the literature on tropical forests (Gentry & Dodson 1987, Galeano et al. 1998, Schnitzer & Carson 2000, Lü et al. 2010), as the lower stratum encompasses transitory and resident species. In these forests, the understory flora may represent 50% or more of the total species (Schnitzer & Carson 2000) and augment the list of tree species up to 30% (Lins-e-Silva 2010).
According to Jardim & Hosokawa (1986), the floristic composition of tropical forests is very different for the upper stratum and understory. Nevertheless, the 54% average similarity among the understory and tree stratum of the different fragments studied here was higher than what is commonly found in the literature (Alves & Metzger 2006). It is important to point out that the herbaceous understory was not sampled and the upper understory was included, which may have contributed to the high similarity found between the canopy and understory. The high richness found by this study for the understory, associated with an intermediate similarity between the understory and the canopy, may also indicate the absence of large scale disturbances in the past (Bohlman & O’Brien 2006), since even higher similarities are expected in early secondary areas (Nascimento 2010).
With the exception of Family Clusiaceae, the high richness and abundance of individuals recorded for families such as Rubiaceae, Melastomataceae, Annonaceae, and Myrtaceae seems to be a standard for flora understory in more humid forests (Schorn & Galvão 2006, Gomes et al. 2009, Onofre et al. 2010, Lü et al.2010), as well as in seasonal forests and savannah (Vilela et al. 1995, Cardoso-Leite et al. 2004, Oliveira & Felfili 2005). Although these families’ richness in the understory is commonly used as an indicator of succession stage, our results show that these families co-occur and are highly important in distinct strata. Thus, in studies with less inclusive size criteria, the family Myrtaceae (which has a higher richness of shade tolerant species) (Tabarelli & Mantovani 1999) is indicative of more mature successional stages. When individuals of the smaller height classes are sampled, Melastomataceae and Rubiaceae are cited as representative of pioneer species (Tabarelli & Mantovani 1999), i.e., of forests in an initial successional stage. Thus, in a more detailed analysis of forest strata, the richness or abundance of these families do not seem to be good indicators of Atlantic Forest successional stages; but yet dependent on the sampling criteria used and on complex characteristics of species’ life history (Clark & Clark 1992).
Species classification into stratification categories: In a manner similar to that already recorded by other authors (Vilela et al. 1995, Salles & Schiavini 2007, Gomes et al. 2009), the understory of the studied fragments is also composed primarily by species of the canopy which are passing through the lower strata. Due to the different methodologies used for sampling this stratum, widely varied percentages for tree species have been recorded in the understory- ranging, for example, from 54.54% (Vilela et al. 1995) to 89.18% (Gomes et al. 2009). Despite this variation, all studies show that the understory is an important transitional environment that serves as an ecological filter for the species that will be part of the canopy, i.e. an environment that selectively determines which individuals and species may survive under its conditions (George & Bazzaz 1999, Harms et al. 2004).
Few species were abundant and most had low density values; this pattern is expected both for the understory (Lü et al. 2010) and for the tree stratum (Richards 1996). These species, classified as rare or locally rare, generally occur in abundance percentages above 25% (Nappo et al. 2004, Oliveira & Amaral 2005); differently from abundant species, these are more likely to be substituted by others as the forest develops, either for natural reasons or due to disturbances in the area (Campos & Landgraf 2001).
Regenerative capacity and vertical structure of the understory: Regarding species’ vertical distribution, most were between 1-2m high, similar to what was found by Silva et al. (2007) and Marangon et al. (2008). The species that were found in all height classes - despite their lower occurrence percentages - must have their growth accompanied and their successional characteristics observed because these are the species with the greatest potential to establish themselves and assure their presence in the forest’s structure and composition (Clark & Clark 1992, Kobe 1999, Silva et al.2007). The exceptions are the species that – due to particular characteristics - are resident and never surpass the lower stratum (Finol 1971).
Species that scored high in relative density and were well distributed vertically also had the highest regeneration rates, as other authors had already recorded (Volpato 1994, Silva et al. 2007, Marangon et al. 2008), with few exceptions. Some species remained among those with high TNR, despite low relative density, because of their homogeneous vertical distribution and higher relative frequency in relation to the others. Nevertheless, some species did not have a high regeneration rate despite their high relative density, which is due to their low relative frequency and absence from at least one height class. In fact, since the TNR index is based upon three species descriptors, it showed to be adequate for ranking understory species according to their importance in the plant assemblage. Furthermore, the species ranking based on the index maintained the same balance between transitory and understory typical species, thus allowing an appropriate interpretation of the data for understanding the contribution of each group to the richness of this stratum.
The results of this study reinforced the idea that the understory of forest fragments is made up mainly of transitory species that are well distributed among the height classes and have higher densities, which favors their future presence in the structure and composition of the canopy. Typical understory species, however, occur in two strata: the lower understory (which includes the species that don’t generally surpass four meters, such as those from families Piperaceae, Rubiaceae and Melastomataceae) and the upper understory (intermediate between the low understory and the canopy and made up of species from families Anonnaceae, Clusiaceae and Myrtaceae, which do not surpass an average of ten meters in height). Our findings support the importance of studying the understory’s vegetation because it is floristically rich, structurally unique, and promotes the natural regeneration of both the lower and tree strata and the recovery from natural or anthropic disturbances. The fundamental aspects of understory assemblages must be prioritized in order to correctly understand the role of transitory and resident species in this stratum.
Acknowledgments
This study is part of the Fragments Project/Phase II - “Sustainability of Atlantic Forest Remnants in Pernambuco and their Implications for Local Conservation and Development”, a Brazil-Germany scientific collaboration program (“Science and Technology for the Atlantic Forest”) financed by CNPq (590039/2006-7) and BMBF (01 LB 0203 A1) with the support of the São José S.A. Sugarcane Mill/ Cavalcanti Petribú Group. The authors thank the support received from the teams of the UFRPE Plant Ecology and Phytosociology Laboratory and the UFPE Plant Morphotaxonomy Laboratory, the Ceará Foundation for Scientific and Technological Development (FUNCAP) and the Brazilian National Council for Scientific Development (CNPq) for the research scholarship provided to the first author. We also thank Lenilson Santos and Marcos Chagas for their indispensable aid in the field.
References
Alves, L.F. & J.P. Metzger. 2006. A regeneração florestal em áreas de floresta secundária na Reserva Florestal do Morro Grande, Cotia, SP. Biota Neotrop. 6: 1-10. [ Links ]
APG III. 2009. Angiosperm Phylogeny Group (APG III). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 161: 105-121. [ Links ]
Aquino, C. & L.M. Barbosa. 2009. Classes sucessionais e síndromes de dispersão de espécies arbóreas e arbustivas existentes em vegetação ciliar remanescente (Conchal, SP), como subsídio para avaliar o potencial do fragmento como fonte de propágulos para enriquecimento de áreas revegetadas no rio Mogi-guaçu, SP. Rev. Árvore 33: 349-358. [ Links ]
Araújo, F.S., S.V. Martins, J.A.A. Meira-Neto, J.L. Lani & I.E. Pires. 2006. Estrutura da vegetação arbustivo-arbórea colonizadora de uma área degradada pós-mineração de caulim, em Brás Pires, MG. Rev. Árvore 30: 107-116. [ Links ]
Armesto, J.J. & J.A. Martínez. 1978. Relations between vegetation structure and slope aspect in the Mediterranean region of Chile. J. Ecol. 66: 881-889. [ Links ]
Benitez-Malvido, J. 1998. Impact of forest fragmentation on seedling abundance in a tropical rain forest. Conserv. Biol. 12: 380-389. [ Links ]
Bohlman, S. & S. O’Brien. 2006. Allometry, adult stature and regeneration requirement of 65 tree species on Barro Colorado Island, Panama. J. Trop. Ecol. 22: 123-136. [ Links ]
Bouroncle, C. & B. Finegan. 2011. Tree Regeneration and Understory Woody Plants Show Diverse Responses to Forest-Pasture Edges in Costa Rica. Biotropica 43: 562-571. [ Links ]
Cadenasso, M.L. & S.T.A. Pickett. 2000. Linking forest edge structure to edge function: mediation of herbivore damage. J. Ecol. 88: 31-44. [ Links ]
Campos, J.C. & P.R.C. Landgraf. 2001. Análise da regeneração natural de espécies florestais em matas ciliares de acordo com a distância da margem do lago. Ci. Fl. 11: 143-151. [ Links ]
Cardoso-Leite, E., T.B. Covre, R.G. Ometto, D.C. Cavalcanti & M.I. Pagani. 2004. Fitossociologia e caracterização sucessional de um fragmento de mata ciliar, em Rio Claro/SP, como subsídio à recuperação da área. Rev. Inst. Flor. 16: 31-41. [ Links ]
Ceccon, E., P. Huante & E. Rincón. 2006. Abiotic factors influencing tropical dry forests regeneration. Braz. Arch. Biol. Techn. 49: 305-312. [ Links ]
Clark, D.A. & D.B. Clark. 1992. Life history diversity of canopy and emergent trees in a neotropical rain forest. Ecol. Monogr. 62: 315-344. [ Links ]
Cowel, R.K. 2009. EstimateS 8.2: Statistical estimator of species richness and shared species from samples. Department of Ecology & Evolutionary Biology, University of Connecticut, Storrs, USA. (Downloaded: 20 October 2011, http://viceroy.eeb.uconn.edu/estimates). [ Links ]
CPRH. 2003. Diagnóstico sócio-ambiental do Litoral Norte de Pernambuco. CPRH, Recife, Pernambuco, Brasil. [ Links ]
Emborg, J. 1998. Understorey light conditions and regeneration with respect to the structural dynamics of a near-natural temperate deciduous forest in Denmark. Forest Ecol. Manage. 106: 83-95. [ Links ]
Felfili, J.M, J.F. Ribeiro, C.W. Fagg & J.W.B. Machado. 2000. Recuperação de Matas de Galeria. EMPRAPA Cerrados: Série Técnica 21: 1-45. [ Links ]
Finol, H.U. 1971. Nuevos parametros a considerarse en el analisis structural de las selvas virgenes tropicales. Rev. Forest. Venez. 14: 29-42. [ Links ]
Galeano, G., S. Suárez & H. Balslev. 1998. Vascular plant species count in a wet forest in the Chocó area on the Pacific Coast of Colombia. Biodivers. Conserv. 7: 1563-1575. [ Links ]
Gama, J.R.V., S.A. Botelho, M.M. Bentes-Gama & J.R.S. Scolforo. 2003. Estrutura e potencial futuro de utilização da regeneração natural de floresta de várzea alta no município de Afuá, Estado do Pará. Ci. Fl. 13: 71-82. [ Links ]
Gentry, A.H. & C. Dodson. 1987. Contribuition of nontrees to species richness of a tropical rain forest. Biotropica 19: 149-156. [ Links ]
George, L.O. & F.A. Bazzaz. 1999. The fern understory as an ecological filter: emergence and establishment of canopy-tree seedlings. Ecology 80: 833-845. [ Links ]
Gilliam, F.S., N.L. Turril & M.B. Adams. 1995. Herbaceous-layer and overstory species in clear-cut and mature central Appalachian hardwood forests. Ecol. Appl. 5: 947-955. [ Links ]
Gilliam, F.S., N.L. Turrill, S.D. Aulick, D.K. Evans & M.B. Adams. 1994. Herbaceous layer and soil response to experimental acidification in a central Appalachian hard- wood forest. J. Environ. Qual. 23: 835- 844. [ Links ]
Gomes, J.S., A.C.B. Lins-e-Silva, M.J.N. Rodal & H.C.H. Silva. 2009. Estrutura do sub-bosque lenhoso em ambientes de borda e interior de dois fragmentos de floresta atlântica em Igarassu, Pernambuco, Brasil. Rodriguésia 60: 295-310. [ Links ]
Harms, K.E., J.S. Powers & R.A. Montgomery. 2004. Variation in small sapling density, understory cover, and resource availability in four Neotropical Forests. Biotropica 36: 40-51. [ Links ]
IBGE. 1992. Manual Técnico da vegetação Brasileira.IBGE (Manuais Técnicos em Geociências), Rio de Janeiro, Rio de Janeiro, Brasil. [ Links ]
Ickes, K., S.J. Dewalt & S. Appanah. 2001. Effects of native pigs (Sus scrofa) on woody understory vegetation in a Malasian lowland rain forest. J. Trop. Ecol. 17: 191-206. [ Links ]
Jardim, F.C.S. & R.T. Hosokawa. 1986. Estrutura da floresta equatorial úmida da estação experimental de silvicultura tropical do INPA. Acta Amaz. 16: 411-508. [ Links ]
Kobe, R.K. 1999. Light gradient partioning among tropical tree species through differential seedling mortality and growth. Ecology 80: 187-201. [ Links ]
Laurance, W.F., H.E.M. Nascimento, S.G. Laurance, A. Andrade, R.M. Ewers, K.E. Harms, R.C.C. Luizão & J.E. Ribeiro. 2007. Habitat Fragmentation, Variable Edge Effects, and the Landscape-Divergence Hypothesis. PLoS ONE 2 : 1-8. [ Links ]
Lima, M.L.F.C. 1998. A reserva da biosfera da Mata Atlântica em Pernambuco- situação atual, ações e perspectivas. Caderno 12, Gráfica do Instituto Florestal, São Paulo, São Paulo, Brasil. [ Links ]
Lima-Filho, D.A., J. Revilla, L.S. Coelho, J.F. Ramos, J.L. Santos & J.G. Oliveira. 2002. Regeneração natural de três hectares de Floresta Ombrófila Densa de Terra Firme na região do rio Urucu-AM, Brasil. Acta Amaz. 32: 555-569. [ Links ]
Lins-e-Silva, A.C.B. 2010. Influência da área e da heterogeneidade de habitats na diversidade vegetal em fragmentos de floresta atlântica. Ph.D. Thesis, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brasil. [ Links ]
Lü, X.T., J.X. Yin & J.W. Tang. 2010. Diversity and composition of understory vegetation in the tropical seasonal rain forest of Xishuangbanna, SW China. Rev. Biol. Trop. 59: 455-463. [ Links ]
Magurran, A.E., 2004. Measuring Biological Diversity. Blackwell Science, Oxford, England. [ Links ]
Marangon, L.C., J.J. Soares, A.L.P. Feliciano & C.F. Brandão. 2008. Regeneração natural em um fragmento de floresta estacional semidecidual em Viçosa, Minas Gerais. Rev. Árvore 32: 183-191. [ Links ]
Martins, S.V. & R.R. Rodrigues. 2002. Gap-phase regeneration in a semideciduous mesophytic forest, Southeastern Brazil. Plant Ecol. 163: 51-62. [ Links ]
Mattei, V.L. & S.J. Longhi. 2001. Avaliação da regeneração natural de Eucalyptus paniculata Smith. Ci. Fl. 11: 55-65. [ Links ]
Metzger, J.P., L.C. Bernacci & R. Goldenberg. 1997. Pattern of tree species diversity in riparian forest fragments of different widths (SE Brazil). Plant Ecol. 133: 135-152. [ Links ]
Mittermeier, R.A., P.R. Gil, M. Hoffman, J. Pilgrim, T. Brooks, C.G. Mittermeier, J. Lamoreux & G.A.B. Fonseca. 2005. Hotspots Revisited - Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions. Chicago, Chicago, Illinois, USA. [ Links ]
Naiman, R.J., H. Décamps & M.E. McClain. 2005. Riparia: ecology, conservation, and management of streamside communities. Elsevier, San Diego, California, USA. [ Links ]
Nappo, M.E., J.J. Griffith, S.V. Martins, P. Marco-Júnior, A.L. Souza & A.T. Oliveira-Filho. 2004. Dinâmica da estrutura fitossociológica da regeneração natural de espécies arbóreas e arbustivas no sub-bosque de povoamento de Mimosa scabrella Bentham, em área minerada, em Poços de Caldas, MG. Rev. Árvore 28: 811-829. [ Links ]
Nascimento, L.M. 2010. Sucessão secundária em área de Mata Atlântica de Pernambuco: mudanças florísticas e estruturais. Ph.D. Thesis, Universidade Federal Rural de Pernambuco, Pernambuco, Brasil. [ Links ]
Oliveira, A.N. & I.L. Amaral. 2005. Aspectos florísticos, fitossociológicos e ecológicos de um sub-bosque de terra firme na Amazônia Central, Amazonas, Brasil. Acta Amaz. 35: 1-16. [ Links ]
Oliveira, E.C.L. & J.M. Felfili. 2005. Estrutura e dinámica da regeneração natural de uma mata de galeria no Distrito Federal, Brasil. Acta Bot. Bras. 19: 801-811. [ Links ]
Oliveira-Filho, A.T., R.J. Almeida, J.M. Mello & M.L. Gavilanes. 1994. Estrutura fitossociológicas e variáveis ambientais em um trecho de mata ciliar do córrego dos Vilas Boas, Reserva Biológica do Poço Bonito, Lavras (MG). Rev. Bras. Bot. 17: 67-85. [ Links ]
Onofre, F.F., V.L. Engel & H. Cassola. 2010. Regeneração natural de espécies da mata atlântica em sub-bosque de Eucalyptus saligna Smith. em uma antiga unidade de produção florestal no Parque das Neblinas, Bertioga, SP. Sci. For. 38: 39-52. [ Links ]
Rayol, P.B., M.F.F. Silva & F.O. Alvino. 2006. Dinâmica da regeneração natural de florestas secundárias no município de Capitão Poço, Para, Brasil. Amazônia: Ciência e Desenvolvimento 2: 93-109. [ Links ]
Ribeiro, M.C., J.P. Metzger, A.C. Martensen, F.J. Ponzoni & M.M. Hirota. 2009. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conserv. 142: 1141-1153. [ Links ]
Richards, P.W. 1996. The tropical rain Forest: an ecological study. Cambridge, Cambridge, England. [ Links ]
Salles, J.C. & I. Schiavini. 2007. Estrutra e composição do estrato de regeneração em um fragmento florestal urbano: implicações para a dinâmica e a conservação da comunidade arbórea. Acta Bot. Bras. 21: 223-233. [ Links ]
Sansevero, J.B.B., P.V. Prieto, L.F.D. Moraes & P.J. Rodrigues. 2011. Natural Regeneration in Plantations of Native Trees in Lowland Brazilian Atlantic Forest: Community Structure, Diversity, and Dispersal Syndromes. Restor. Ecol. 19: 379-389. [ Links ]
Santos, R., V. Citadini-Zanette, L.S. Leal-Filho & W.T.Hennies. 2008. Spontaneous Vegetation on Overburden Piles in the Coal Basin of Santa Catarina, Brazil. Restor. Ecol. 16: 444-452. [ Links ]
Santos, S.L. & I.F.M. Válio. 2002. Litter accumulation and its effects on seedling recruitment in a Southeast Brazilian Tropical Forest. Rev. Bras. Bot. 25: 89-92. [ Links ]
Scariot, A. 2000. Seedling mortality by litterfall in Amazonian forest fragments. Biotropica 32: 662-669. [ Links ]
Schnitzer, S.A. & W.P. Carson. 2000. Have we forgotten the forest because of the trees? Trends Ecol. Evol. 15: 375-376. [ Links ]
Schorn, L.A. & F. Galvão. 2006. Dinâmica da regeneração natural em três estágios sucessionais de uma floresta ombrófila densa em Blumenau, SC. Floresta 36: 59-74. [ Links ]
Silva, H.C.H., A.C.B. Lins-e-Silva, J.S. Gomes & M.J.N. Rodal. 2008. The effects of internal and external edges on vegetation physiognomy and structure in a remnant of atlantic lowland rainforest in Brazil. Biorem. Biodiv. Bioavail. 2: 47-55. [ Links ]
Silva, W.C., L.C. Marangon, L.C. Ferreira, A.L.P. Feliciano & R.F. Costa-Júnior. 2007. Estudo da regeneração natural de espécies arbóreas em fragmento de floresta ombrófila densa, Mata das Galinhas, no município de Catende, Zona da mata sul de Pernambuco. Ci. Fl. 17: 321-331. [ Links ]
Souza, A.C.R., E.B. Almeida Jr. & C.S. Zickel. 2009. Riqueza de espécies de sub-bosque em um fragmento florestal urbano, Pernambuco, Brasil. Biotemas 22: 57-66. [ Links ]
Souza, A.L., G.F. Silva, J.F. Chichorro & R.L. Caraciolo. 2006. Mata Nativa 2: Manual do usuário. Cientec, Viçosa, Minas Gerais, Brasil. [ Links ]
Tabarelli, M. & W. Mantovani. 1999. A regeneração de uma Floresta Tropical Montana após corte e queima (São Paulo-Brasil). Rev. Bras. Biol. 59: 239-250. [ Links ]
The Internacional Plant Names Index. 2010. (Downloaded: 10 January 2011, http://www.ipni.org. [ Links ]).
Trindade, M., A.C.B. Lins-e-Silva, H.P. Silva, S.B. Figueira & M. Schessl. 2008. Fragmentation of the Atlantic Rainforest in the northern coastal region of Pernambuco, Brazil: recent changes and implications for conservation. Biorem. Biodiv. Bioavail. 2: 5-13. [ Links ]
Triquet, A.M., G.A. Mcpeek & W.C. Mccomb. 1990. Songbird diversity in clear cuts with or without a Riparian Buffer Strip. J. Soil Water Conserv. 45: 500-503. [ Links ]
Tropicos. 2010. Missouri Botanical Garden, Missouri, USA. (Downloaded: 10 January 2011, http://www.tropicos.org). [ Links ]
Válio, I.F.M. 2003. Seedling Growth of Understorey Species of a Southeast Brazilian Tropical Forest. Braz. Arch. Biol. Techn. 46: 697-703. [ Links ]
Vilela, E.A., A.T. Oliveira-Filho, D.A. Carvalho & M.L. Gavillanes. 1995. Flora arbustivo-arbórea de um fragmento de mata ciliar no Alto Rio Grande, Itutinga, Minas Gerais. Acta Bot. Bras. 9: 87-100. [ Links ]
Volpato, M.M.L. 1994. Regeneração natural em uma floresta secundária no domínio de Mata Atlântica: uma análise fitossociológica. Master Thesis, Universidade Federal de Viçosa, Minas Gerais, Brasil. [ Links ]
*Correspondencia:
Juliana Silva Gomes-Westphalen:Universidade Federal do Ceará, Campus do Pici, Programa de Pós-Graduação em Ecologia e Recursos Naturais, Bloco 906, 60455-760, Fortaleza, CE, Brasil. juli.ufrpe@gmail.com,
Ana Carolina Borges Lins-e-Silva: Universidade Federal Rural de Pernambuco, Departamento de Biologia, Área de Ecologia, 52171-900, Recife, PE, Brasil. anacarol@db.ufrpe.br
Francisca Soares de Araújo: Universidade Federal do Ceará, Campus do Pici, Programa de Pós-Graduação em Ecologia e Recursos Naturais, Bloco 906, 60455-760, Fortaleza, CE, Brasil. tchesca@ufc.br
1. Universidade Federal do Ceará, Campus do Pici, Programa de Pós-Graduação em Ecologia e Recursos Naturais, Bloco 906, 60455-760, Fortaleza, CE, Brasil; juli.ufrpe@gmail.com, tchesca@ufc.br
2. Universidade Federal Rural de Pernambuco, Departamento de Biologia, Área de Ecologia, 52171-900, Recife, PE, Brasil; anacarol@db.ufrpe.br
Received 11-VIII-2011. Corrected 06-II-2012. Accepted 05-III-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}