










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.57 suppl.1 San José Nov. 2009
Dermatoglyphic traits of six Chibcha-speaking Amerindians of Costa Rica, and an assessment of the genetic affinities among populations
Maia Segura-WW Ramiro Barrantes
Escuela de Biología, Universidad de Costa Rica, 11501-22060, San José, Costa Rica; rabar@cariari.ucr.aac.ccr
Abstract:Dermatoglyphic traits have been used to evaluate population structure and microdifferentiation in several populations. For Chibcha-speaking groups of Lower Central America there are few dermatoglyphic studies, but extensive linguistic, anthropological and genetic data support their historical, cultural and biological relationships. The main objectives of this study were to describe new dermatoglyphic data for six Chibcha- speaking Amerindians of Costa Rica, and to assess the relationships between these and other Amerindian and Eskimo groups, at different levels of population differentiation by means of multivariate analyses of quantitative traits. Sexual (x2 =227.22, df=33, p<0.01),, and bimanual (x2 =554.45, df=33, p<0.01) differences were both significant for the overall population, as has been reported previously. Remarkably, higher frequencies of arches, lower frequencies of whorls and lower means of total ridge counts were observed in the tribes analyzed compared with other American indians. At the lowest level of population differentiation, two Cabecar subpopulations (Aatlantic and Chirripo) were compared and no significant differences were found (FF=0.001, p=0.72),, suggesting that dermatoglyphic variation might not reflect known genetic divergence at this level of association. Comparisons within the Chibchan dataset using Principal Components Analysis (PPCA) placed the Huetar and the Cabecar in close proximity, and separated the Guatuso and the Guaymi. Additionally, the Chibchan tribes, although showing nearer proximity to Non-Andean South American groups, can be separated from other Amerindian and Eskimo populations, confirming previous results based on extensive genetic surveys and linguistic analyses that have demonstrated the existence of a Chibchan cluster within a larger South American phylogenetic group. The results obtained support the use of dermatoglyphics to assess interpopulation affinities, even at the level of tribes. Rev. Biol. Trop. 57 (SSuppl. 1): 357-369. Epub 2009 November 30.
Key words: Dermatoglyphics, Chibcha-speaking Amerindians, genetic affinities, PCA, Costa Rica.
The genetic structure and microevolution of Chibcha-speaking Amerindian populations have been studied during the last decades through the analysis of linguistic (Constenla- Umaña 1990), archaeological (Cooke 2005, Hoopes & Fonseca 2003), and genetic (Barrantes 1993) data. Detailed descriptions of genetic variation and phylogenetic relationships among these populations have been obtained using several molecular markers, including blood groups, red cell and serum protein systems (Barrantes et al . 1990, Barrantes 1993, Azofeifa et al .2001), genetic analysis of mtDNA (Santos et al . 1994, Torroni et al.1994, Batista et al . 1995, Kolman et al . 1995, Melton et al . 2007) and Y chromosome markers (Ruiz- Narváez et al . 2005), demonstrating the relatively isolated development of these groups and their low genetic variability.
The existence of dermatoglyphic data for these Chibcha-speaking populations constitutes an opportunity, first to describe new facts on Chibchan groups, and second, to evaluate and compare the results in terms of the existing, well defined, historical associations among them. Most studies of dermatoglyphic traits have been descriptive, although some have analyzed population structure and microdifferentiation (Jantz 1987, Crawford & Duggirala 1992, Sokal & Livshits 1993) based on dermal traits which are believed to have high heritabilities (Meier 1980). Other studies have assessed the correlations between different sets of dermatoglyphic, genetic, and geographic distances obtaining both significant (Crawford & Duggirala 1992) and non-significant associations (Neel et al. 1974).
Descriptions of dermatoglyphic traits in Amerindians have focused in Eskimo populations, North American Indians and various Mayan and non Mayan groups (reviewed by Crawford 1998). Additionally, other South American populations have also been considered (reviewed by Salzano & Callegari-Jacques 1988). However, there is still a general lack of information of dermatoglyphic traits in Amerindian tribes located in Lower Central America. Preliminary descriptive studies of Chibchaspeaking Amerindians inhabiting this area have been performed in the Boruca, the Guaymi, the Bribri and the Cabecar (Quesada & Barrantes 1983, 1984, 1991), as separate investigations, but have not evaluated the associations, neither within these tribes, nor among these and other Amerindian populations. In this study these questions are addressed through the analysis of several ermatoglyphic variables, including new data of Chibcha-speaking Amerindians and comparisons among other American Indian populations. Therefore, the main objectives of the present study are twofold: (1) to present new results on digital and palmar dermatoglyphic characteristics of six Chibchaspeaking Amerindian groups of Costa Rica; and, (2) to assess the relationships among the tribes at diverse levels of population differentiation, i.e. at the individual level; among nearby Chibcha-speaking groups; and among Chibcha and non-Chibcha American Indian populations, by using Principal Components Analysis (PCA) of quantitative dermatoglyphic traits.
Materials and methods
The patterns of dermatoglyphic traits were studied in 743 individuals from six Chibchaspeaking populations of Costa Rica: Guaymi, Bribri, Boruca, Cabecar, Guatuso and Huetar. The records from these two last tribes, as well as two localities from the Cabecar constitute unpublished new data. Information of dermatoglyphics was obtained during field visits to different localities between 1979 and 1995. The geographic location of each sampling site is shown in Table 1, and the number of individuals analyzed in each group is given in Table 2. Finger and palmar prints were collected and characterized using the standard ink method and classifying systems described by Cummins & Midlo (1961) and by Penrose (1968).

For each individual an overall total of 44 digital and palmar dermatoglyphic variables were studied, distributed as follows: 1. Qualitative variables (10 for each hand): types of digital patterns on every finger; presence or absence of palmar patterns on I2, I3 and I4 interdigital areas; patterns on thenar/interdigital and hypothenar palmar areas. 2. Quantitative variables (12 for each hand): finger ridge counts for each digit, total ridge count, pattern intensity index, main line index, A-B, B-C and C-D ridge counts, and ATD angle.
Digital pattern types were classified, as usual, in four categories defined as: arches, ulnar loops, radial loops and whorls. The frequencies of each category were estimated for the total population, for the two sexes and for both hands separately. Similarly, the frequencies of palmar patterns were also calculated. Quantitative variables were described using the mean and standard deviation per population. Bimanual and sexual comparisons were performed using x2 test and Students t-test for qualitative and quantitative variables respectively.
To analyze if the differences in dermatoglyphic characteristics reflect currently known variation between populations, the Cabecar were subdivided in two groups following geographic, linguistic, genetic and ethnographic sources (Barrantes 1993): the Chirripo population comprising individuals from Chirripo and Talari (localities shown in Table 1); and the Atlantic group, including populations from Amubri and Mojoncito. The two groups haveshown particular genetic and linguistic characteristics that suggest certain divergence between populations (Azofeifa et al. 001). To test for differences in dermatoglyphic characteristics that could support the genetic results, a multivariate analysis of variance of the quantitative variables was used. Before performing the analysis, the variables were tested for the basic assumptions of normality and homoscedasticity. A posteriori analyses of variance were used to evaluate the differences between individual variables.
The affinities among the six Amerindian populations were assessed by multivariate analysis of the dermatoglyphic quantitative variables using Principal Components Analysis (PCA), as implemented in SPSS 15.0 (SPSS 2006). This analysis is helpful to reduce the correlation between variables when large data sets are analyzed, and to identify general trends of the data for a more efficient discrimination of the variation among tribes. The first three principal components were extracted from the correlation matrices, followed by varimax rotation. Factor score coefficients of each population for the first three principal components were calculated from the mean value of each variable for males and females separately, and were subsequently used to create plots to show the dermatoglyphic relationships among the populations studied (Arrieta et al. 1991, Micle & Kobyliansky 1986). To evaluate the differences between dermatoglyphic characteristics of hibcha-speaking Amerindians and other American Indian populations, the male means for the Chibchan sample were calculated. Comparisons were made by means of PCA using the variables reported by Garruto et al. (1979): means for pattern intensity index, total ridge counts, main line index, A-B ridge count, presence of palmar patterns in the hypothenar, thenar/interdigital, and interdigital areas. These authors summarized data of male Eskimo, North, Central and South American Indians and these last two groups were subclassified into Mayan and Non-Mayan; and into Andean and Non-Andean, respectively. As in the previous analysis, a plot using the first three principal components was used to show the relationship between these populations.
Results
The percentage frequencies of digital patterns in both hands in males and females for the six populations analyzed are shown in Table 2. For the overall population, significant differences were found between males and females (2=27.22, df=3, p<0.01), and between right and left hands (x2=54.45, df=3, p<0.01). Differences among sexes are mainly due to higher frequencies of arches, ulnar loops and whorls in females. The differences found for right and left hands can be attributed to a higher incidence of arches and whorls on the left hand. These results confirm previous reports of sexual and bimanual differences in several populations for both quantitative and qualitative traits (Quesada & Barrantes 1984, Micle & Kobyliansky 1986, Arrieta et al. 1987, 1991, Martín et al. 1996).
In most tribes, ulnar loops are the most frequent pattern in both sexes. In contrast, the least frequent patterns are radial loops for both males and females, except for the Guatuso, where arches show the lowest frequency. Similar results were observed and described by Quesada & Barrantes (1991) for samples from Bribri and Cabecar populations. South American Indians are also characterized by low frequencies of radial loops and high frequencies of ulnar loops (Demarchi & Marcellino 1998) and Eskimo populations show similar results (Crawford & Duggirala 1992). However, some important differences can also be addressed, including higher values for arches, lower for whorls, and lower mean total ridge count, as will be mentioned, in Chibchan tribes when compared to data describing other Central, North and South Americans (Garruto et al. 1979), and Caucasian, Oriental and African populations (Plato 1983),
Absolute frequencies of palmar dermatoglyphic patterns in the interdigital zones, thenar/I and hypothenar areas, in the Chibchan groups are presented in Table 3. The absence of patterns in the I2 interdigital area for almost all populations is not unique. In fact, this has been shown to be a ubiquitous characteristic in several reports (Garruto et al. 1979, Hoff et al. 1981). Conversely, the I4 area shows the highest frequency of patterns among the interdigital areas in all populations. No significant differences were found between sexes among all populations in the frequencies of patterns in the I3 (x2=14.12, df=8, p>0.05) nor I4 (x2=7.19, df=8, p>0.05) areas. Absence of small vestigial pattern in thenar/interdigital I palmar area and absence of whorls in the hypothenar area are the predominant characteristics among all populations.
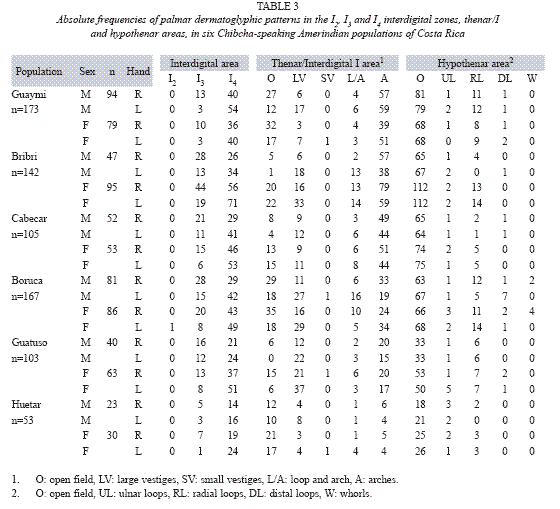
Table 4 shows the frequencies of quantitative variables in each Chibcha-speaking Amerindian tribe, for males and females, and both hands. The mean total ridge count for the overall population was 96.98, which is lower in these groups compared to other North, Central and South American Indians with values ranging from 127.8 to 133.5 (Garruto et al. 1979). For the other quantitative variables, no major differences were observed.
When the two Cabecar subgroups were analyzed, no significant differences were obtained (F=0.001, p=0.72) between the Chirripo and Atlantic populations. Even when a posteriori analyses of variance showed significant differences for ATD angle (F1,104=13.86, p<0.001), no other variable differed between the two localities. These results suggest that at this level of population differentiation, dermatoglyphics might not indicate actual genetic divergence.
Based on the multivariate analysis of dermatoglyphic characteristics three principal components were extracted (Table 5). These components account for 63.13% of the total variance of the traits considered, 57.00% infemales and 64.92% in males. For the overall population analysis, the first principal component is highly correlated with ridge counts and with pattern intensity index in both hands, total ridge count shows the highest correlation coefficient with this component. When sexes were analyzed independently similar results are found. Several previous studies have consistently reported that the first component eveals the size of the digital patterns and the variation in the pattern intensity index (Micle & Kobyliansky 1986, Arrieta et al. 1991).
For the second component almost all the variables have positive correlation coefficients, which indicate that they all contribute to the formation of this component, although B-C and C-D ridge counts for both right and left hands have higher scores and their contribution is more important than the other variables. The third component describes the variability of the ATD angle measurement.
Plots showing the affinities among populations for males and females based on the first three components are presented in Fig. 1. In males, the Guatuso and the Bribri show positive scores for both components, consistent with the high total ridge counts and B-C/D ridge counts in the these tribes (Table 4). The male samples of the Boruca are separated mainly due to high mean values of B-C and C-D ridge counts in the former, and higher total ridge counts in the later. The Guaymi males and females have the most negative scores for the three components, which can be attributed to their low means for the variables describing these components.
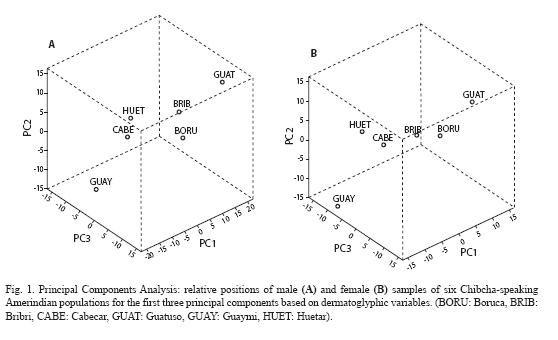
In females, a cluster similar to that described for males is formed by the Guatuso, the Bribri and the Boruca populations, exhibiting high positive scores with components I and II. The Huetar and the Cabecar can be grouped in another cluster defined by negative scores for the first component and positive values for the second component.
A separate PCA was performed to analyze the relationships between several American Indian populations and the Chibcha-speaking Amerindians. The results obtained from this analysis are plotted in Fig. 2. The first three principal components explain 86.21% of the variance. PC1 shows the highest correlation with total ridge count and digital patterns, particularly whorls, PC2 is highly correlated with patterns in I2 area and A-B ridge count, and PC3 is correlated with patterns in the I4 interdigital area, indicating the value of these variables to assess the associations between Amerindian populations. As can be seen, there is clear separation between Central, North American groups and Eskimo populations. As expected, the Chibcha-speaking Amerindians show relatively higher affinity with South American populations, although maintaining their autonomy as a separate group, confirming previous results obtained from genetic, archaeological and linguistic approaches (Barrantes et al. 1982, Layrisse et al. 1995, Hoopes & Fonseca 2003) and more recent analysis of larger sets of genetic markers (Wang et al. 2007).

Discussion
The frequencies of digital pattern types presented for the Chibcha-speaking Amerindian tribes in this study are generally in agreement with published data for other Amerindians and Eskimo populations. High frequencies of ulnar loops and lower of radial loops, as well as the distribution of patterns between fingers, are consistent with other investigations. For example, Crawford (1998) reviewed several studies of dermatoglyphics in Eskimo and North American Indian populations finding higher incidences of ulnar loops, while Salzano and Callegari-Jacques (1988) reviewed the data available for South American Indians and observed the same trend. This outcome is not exclusively Amerindian as it seems to be common among groups worldwide such as Israeli (Micle & Kobyliansky 1986) and Spanish (Martín et al. 1996) populations. The frequencies of patterns in interdigital zones, thenar/I and hypothenar areas are also in agreement with other tribes (Hoff et al. 1981). On the other hand, Chibcha-speaking tribes can be distinguished from other American populations by their high frequency of arches, low frequency of whorls and low mean total ridge count.
The significant sexual and bimanual differencesare not unexpected results due to previous reports of similar outcomes for several populations (Micle & Kobyliansky 1986,Arrieta et al. 1987, 1991, Martín et al. 1996). The reasons for sexual dimorphism observed in the analyzed variables are still controversial, although some evidences support the fact that differences in heritability and developmental variation among sexes might account for these patterns (Meier 1980). On the other hand, bimanual differences have been attributed to developmental instability, measured by fluctuating asymmetry of bilateral traits. In the particular case of dermatoglyphics, fluctuating asymmetry must result from environmental assaults during early embryonary stages, as dermal ridges adopt fixed configurations during the first 12 to 16 weeks of fetal development, making subsequent alterations highly improbable (Cummins & Midlo 1961).
Interpopulation comparisons yielded contrasting results depending on the hierarchical levels of population differentiation tested. At the lowest level of differentiation, i.e. the comparisons between the two Cabecar subgroups, the dermatoglyphic traits analyzed do not provide enough information to separate the populations, even when the evaluation of genetic variability suggests a population subdivisionamong the Cabecar (Barrantes 1993). The Chirripo subpopulation is located on the Atlantic side of the Cordillera de Talamanca (Talamanca Mountains) near, although not overlapping, the other Atlantic subpopulation described in this paper. The geographical distribution could account for the genetic divergence of the twogroups; however, these differences are not reflected in the dermatoglyphic traits measured in this study. Only ATD angle was significantly ifferent between the two subpopulations. Other studies have found that ATD angle is oneof the most important variables for population discrimination (Reddy et al. 2001), probably due to its low population variation, or because it is among the least environmentally affected dermatoglyphic traits, as measured by reduced fluctuating asymmetry (Karmakar & Malhotra 1995).
At a higher level of differentiation, based on the multivariate analysis of quantitative variables it has been shown that the first three principal components allow the definition of population clusters among the six Chibchaspeaking Amerindians of Costa Rica as plotted in Fig. 1. The associations obtained are not particularly related to the extant geographical distribution of the screened tribes, and consequently no clear geographical tendencies are detected. On the other hand, as revealed by the PCA scaling plot, these patterns are not inconsistent with molecular markers and linguistic data describing the separate position of the Guatuso and the Guaymi (Barrantes et al. 1990); neither with known ethnohistorical affiliations, suggesting certain affinities between the Cabecar and the Huetar, even when these relations were not confirmed by genetic analysis (Azofeifa et al. 2001). It is possible that these results might have arisen from shared ancestry or admixture processes in the past and not from recent gene flow between neighboring tribes. Rothhammer et al. (1977) also reported non-significant correlations between genetics and dermatoglyphics at the inter-tribal level of variation in Yanomama Indian villages. Additionally, these authors found varying degrees of concordance between genetic markers and dermal traits, with increasing significance of this relationship when comparing more distant groups. For example, when comparing South American Indian tribes with other populations of different continental regions, good congruence between the two sets of characteristics was achieved.
In the same way, it has been shown that clear distinction between Chibchan groups and other tribes along the American continent can be accomplished by using dermatoglyphics, and the patterns revealed are consistent with associations defined on the basis of genetic and linguistic evidence. For instance, the observed separation between the examined Chibchan populations and other non- Chibcha-speaking Central American Indians (Mayan & Non- Mayan groups), as shown in Fig. 2, has been demonstrated by genetic markers (Wang et al. 2007). Furthermore, at this level of population differentiation, the PCA revealed a nearer relationship between Chibcha-speaking Amerindian tribes and South American populations. This result is in agreement with evidence from further studies that indicate the presence of cultural and biological similarities between Lower Central American and northern South American Chibcha-speaking populations, probably resulting from the expansion of the former into the south, as has been suggested by the possible geographic origin of the protochibcha language (Constenla-Umaña 1995) and by extensive genetic analyses showing a strongly supported separate subcluster of Chibcha speakers within a South American cluster (Melton et al. 2007, Wang et al. 2007).
The obtained associations support the importance of dermatoglyphics for the study of interpopulation relationships to elucidate patterns of population structure of Amerindian populations, as has been demonstrated in by other investigations (Micle & Kobyliansky 1985, Gualdi-Russo et al. 1994), although it should be noticed that not all dermatoglyphic traits are equally effective in separating the tribes analyzed; the types of digital patterns are among the most important traits accounting for the variation between the groups, similar to results previously found for other populations (Hoff et al. 1981, Micle & Kobyliansky 1986, Arrieta & Lostao 1988). Earlier studies have pointed out the large variability of Amerindians regarding various anthropological characters, including dermatoglyphics (Leguebe & Vrydagh 1981, Salzano & Callegari-Jacques 1988, Crawford 1998), explained in part by the differential degrees of admixture. Given the generally accepted fact that dermatoglyphics are under genetic control it is possible that demographic processes and the interaction between genetics and environment have diverse consequences on dermatoglyphic traits and could account for the differences observed. Nevertheless, contrasting arguments still exist concerning the effects of environment, gene flow, genetic drift, and consequently population size, over dermatoglyphic variables (Meier 1980, Arrieta & Lostao 1988).
Acknowledgments
To Misael Quesada, pioneer in dermatoglyphic research in Costa Rica, for collecting and characterizing finger and palmar prints. This project was funded by the University of Costa Rica through Program 742-88-065, and the Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICIT).
Resumen
Los dermatoglifos se han utilizado para evaluar la estructura poblacional y microdiferenciación de varias poblaciones. Para los grupos chibcha de Baja Centroamérica hay pocos estudios sobre dermatoglifos pero los datos lingüísticos, antropológicos y genéticos muestran la existencia de relaciones históricas, culturales y biológicas. Los objetivos del presente estudio fueron describir nuevos datos de dermatoglifos para seis tribus amerindias chibcha de Costa Rica y evaluar las relaciones entre estas y otros grupos amerindios y esquimales, a diferentes niveles de diferenciación poblacional por medio de análisis multivariados. Se encontraron diferencias significativas entre ambos sexos (2=27.22, df=3, p<0.01) y ambas manos (2=54.45, df=3, p<0.01), similar a lo descrito para otras poblaciones. Las tribus estudiadas se caracterizan por presentar alta frecuencia de arcos, baja frecuencia de verticilos y bajo conteo total de líneas. Al nivel más bajo de diferenciación poblacional, se compararon dos subpoblaciones cabécar (Atlántico y Chirripo) y no se encontraron diferencias significativas (F=0.001, p=0.72) lo cual sugiere que los dermatoglifos no permiten discriminar entre grupos a este nivel. Las comparaciones entre las tribus chibcha estudiadas por medio de análisis de componentes principales (PCA) ubican a los huetar cercanos a los cabécar; mientras que los guatuso y guaymí aparecen como grupos más aislados. Adicionalmente, el grupo chibcha, aunque muestra mayor afinidad con poblaciones suramericanas, puede separarse de otras tribus amerindias y esquimales, confirmando los resultados de estudios genéticos y lingüísticos que han colocado a los chibchas dentro del un grupo filogenético mayor formado por tribus amerindias de Suramerica. Dichos resultados confirman el valor de las características dermatoglíficas para evaluar las afinidades interpoblacionales aún a nivel de tribus.
Palabras clave: dermatoglifos, amerindios chibcha, afinidades genéticas, PCA, Costa Rica.
Received 03-X-22007. Corrected 30-VIII-22009. Accepted 04-X-22009.
References
Arrieta, M. & C. Lostao. 1988. Principal components analysis of digital dermatoglyphic patterns in a Basque population. Hum. Biol. 60: 191-204. [ Links ]
Arrieta, M., B. Criado, B. Martínez, A. Simon, L. Salazar & C. Lostao. 1991. Characterization of digital dermatoglyphic features in Salazar Valley: comparison with other Basque valleys. Ann. Hum. Biol. 18: 221-233. [ Links ]
Arrieta, M., M. Ibarrondo & C. Lostao. 1987. Digital dermatoglyphics in the Basque Population: univariate and multivariate comparisons with other Spanish populations. Am. J. Phys. Anthropol. 73: 89-98. [ Links ]
Azofeifa, J., E. Ruiz & R. Barrantes. 2001. Blood group, red cell, and serum protein variation in the Cabecar and Huetar, two Chibchan Amerindian tribes of Costa Rica. Am. J. Hum. Biol. 13: 57-64. [ Links ]
Barrantes, R. 1993. Diversidad genética y mezcla racial en los amerindios de Costa Rica y Panamá. Rev. Biol. Trop. 41: 379-384. [ Links ]
Barrantes, R., P. Smouse, H. Mohrenweiser, H. Gershowitz, J. Azofeifa, T. Arias & J. Neel. 1990. Microevolution in lower Central America: genetic characterization of the Chibcha-speaking groups of Costa Rica and Panamá, and a consensus taxonomy based on genetic and linguistic affinity. Am. J. Hum. Genet. 46: 63-84. [ Links ]
Barrantes, R., P. Smouse, J. Neel, H. Mohrenweiser & H. Gershowitz. 1982. Migration and genetic infrastructure of the Central American Guaymi and their affinities with other tribal groups. Am. J. Phys. Anthropol. 58: 201-214. [ Links ]
Batista, O., C. Kolman & E. Bermingham. 1995. Mitochondrial DNA diversity in the Kuna Amerinds of Panamá. Hum. Mol. Genet. 4: 921-929. [ Links ]
Constenla-Umaña, A. 1990. Hipótesis sobre la localización del protochibcha y la dispersión de sus descendientes. Revista de Filología y Lingüística de la Universidad de Costa Rica 16: 111-123. [ Links ]
Constenla-Umaña, A. 1995. Sobre el estudio diacrónico de las lenguas chibchenses y su contribución al conocimiento del pasado de sus hablantes. Boletín Museo del Oro 38-39: 13-55. [ Links ]
Crawford, M. 1998. The Origins of Native Americans. Evidence from Anthropological Genetics. Cambridge, New York, USA. [ Links ]
Crawford, M. & R. Duggirala. 1992. Digital dermatoglyphic patterns of Eskimo and Amerindian populations: Relationships between geographic, dermatoglyphic, genetic, and linguistic distances. Hum. Biol. 64: 683-704. [ Links ]
Cooke, R. 2005. Prehistory of Native Americans on the Central American land bridge: colonization, dispersal, and divergence. J. Archaeol. Res. 13: 129-187. [ Links ]
Cummins, H. & C. Midlo. 1961. Finger Prints, Palms and Soles. An Introduction to Dermatoglyphics. Dover Publications, New York, USA. [ Links ]
Demarchi, D. & A. Marcellino 1998. Dermatoglyphic relationships among South Amerindian populations. Hum. Biol. 70: 579-596. [ Links ]
Garruto, R., C. Plato, C. Hoff, M. Newman, D. Gajdusek & P. Baker. 1979. Characterization and distribution of dermatoglyphic features in Eskimo and North, Central, and South American Indian populations. Birth Defects-Orig. 15: 277-334. [ Links ]
Gualdi-Russo, E., F. Facchini, P. Brasili-Gualandi, G. Gruppioni & D. Pettener. 1994. Digital and palmar dermatoglyphics and the population structure of the Upper Bologna Apennine. Ann. Hum. Biol. 21: 519-532. [ Links ]
Hoff, C., C. Plato, R. Garruto & J. Dutt. 1981. Dermatoglyphic assessment of the genetic relationships of Native American populations. Am. J. Phys. Antropol. 55: 455-461. [ Links ]
Hoopes, J. & O. Fonseca. 2003. Goldwork and Chibchan Identity: Endogenous Change and Diffuse Unity in the Isthmo-Colombian Area, p. 49-89. In J. Quilter & J. Hoopes (eds.). Gold and Power in Ancient Costa Rica, Panama, and Colombia. Dumbarton Oaks, Washington D.C, USA. [ Links ]
Jantz, R. 1987. Anthropological dermatoglyphic research. Ann. Rev. Anthropol. 16: 161-177. [ Links ]
Karmakar, B. & K. Malhotra. 1995. Variation in maximal angle. Int. J. Anthropol. 10: 163-174. [ Links ] olman, C., E. Bermingham, R. Cooke, R. Ward, T. Arias & F. Guionneau-Sinclair. 1995. Reduced mtDNA diversity in the Ngöbé Amerinds of Panamá. Genetics 140: 275-283.
Layrisse, Z., M. Layrisse, A. Rodríguez-Larralde. 1995. Estudios genéticos en poblaciones de lengua chibcha. Boletín Museo del Oro 38-39: 71-85. [ Links ] Leguebe, A. & S. Vrydagh 1981. Geographic variability of digital ridge-counts: principal components analysis of male and female world samples. Ann. Hum. Biol. 8: 519-528. [ Links ]
Martín, J., M. Mesa, V. Fuster & P. Moral. 1996. Dermatoglyphics of inhabitants of Alberche/Tormes Valley (Sierra de Gredos-Central Spain): I. Finger pattern types and pattern intensity. Am. J. Hum. Biol. 8: 305-316. [ Links ]
Meier, R. 1980. Anthropological dermatoglyphics: a review.Yearbook Phys. Anthropol. 23: 147-178. [ Links ]
Melton, P., I. Briceño, A. Gómez, E. Devor, J. Bernal & M. Crawford. 2007. Biological relationship between Central and South American Chibchan speaking populations: evidence from mtDNA. Am. J. Phys. Anthropol. 133: 753-770. [ Links ]
Micle, S. & E. Kobyliansky. 1985. Dermatoglyphic distances between Israeli Jewish population groups of different geographical extraction. Hum. Biol. 57: 97-111. [ Links ]
Micle, S. & E. Kobyliansky. 1986. Dermatoglyphic sexual dimorphism in Israelis: principal components and discriminant analyses applied to quantitative traits. Hum. Biol. 58: 485-498. [ Links ]
Neel, J., F. Rothhammer & J. Lingoes. 1974. The genetic structure of a tribal population, the Yanomama Indians. X. Agreement between representations of village distances based on a different set of characteristics. Am. J. Hum. Genet. 26: 281-303. [ Links ]
Penrose, L. 1968. Memorandum on dermatoglyphic nomenclatura. Birth Defects-Orig. 4: 1-12. [ Links ] Plato, C. 1983. The Worldwide Distribution of Dermatoglyphics, p. 365-376. In I. Bansal (ed.). Human Biology- Recent Advances-Vol. 2. Today & Tomorrows Printers and Publishers. New Delhi, India. [ Links ]
Quesada, M. & R. Barrantes. 1983. Dermatoglifos en dos poblaciones indígenas guaymí de Costa Rica. Rev. Biol. Trop. 31: 269-275. [ Links ]
Quesada, M. & R. Barrantes. 1984. Rasgos dermatoglíficos de los indígenas de Boruca, Costa Rica, p. 185-189. In R. Barrantes, M. Bozzoli & P. Gudiño (eds.). Memorias del Primer Simposio Científico sobre Pueblos Indígenas de Costa Rica. Instituto Geográfico de Costa Rica. San José, Costa Rica. [ Links ]
Quesada, M. & R. Barrantes. 1991. Dermatoglifos en los amerindios Bribri y Cabécar de Costa Rica. Rev. Biol. Trop. 39: 63-70. [ Links ]
Reddy, B., A. Pfeffer, M. Crawford & B. Langstieh. 2001. Population substructure and patterns of quantitative variation among the Gollas of Southern Andhra Pradesh, India. Hum. Biol. 73: 291-306. [ Links ]
Ruiz-Narváez, E., F. Santos, D. Carvalho-Silva, J. Azofeifa, R. Barrantes & S. Pena. 2005. Genetic variation of the Y chromosome in Chibcha-speaking Amerindians of Costa Rica and Panama. Hum. Biol. 77: 71-91. [ Links ]
Rothhammer, F., R. Chakraborty & E. Llop. 1977. A collation of gene and dermatoglyphic diversity at various levels of population differentiation. Am. J. Phys. Anthropol. 46: 51-60. [ Links ]
Salzano, F. & S. Callegari-Jacques. 1988. South American Indians. A Case Study in Evolution. Oxford, New York, USA. [ Links ]
Santos, M., R. H. Ward & R. Barrantes. 1994. mtDNA variation in the Chibcha Amerindian Huetar from Costa Rica. Hum. Biol. 66: 963-977. [ Links ]
Sokal, R. & G. Livshits. 1993. Geographical variation of six dermatoglyphic traits in Eurasia. Am. J. Phys. Anthropol. 90: 393-407. [ Links ]
Statistical Package for the Social Sciences (SPSS). 2006. SPSS 15.0 Base Users Guide. SPSS Inc., Chicago, USA. [ Links ]
Torroni, A., J. Neel, R. Barrantes, T. Schurr & D. Wallace. 1994. Mitochondrial DNA "clock" for the Amerinds and its implications for timing their entry into North America. Proc. Natl. Acad. Sci. USA 91: 1158-1162. [ Links ]
Wang, S., C. Lewis Jr., M. Jakobsson, S. Ramachandran, N. Ray, G. Bedoya, W. Rojas, M. Parra, J. Molina, C.Gallo, G. Mazzotti, G. Poletti, K. Hill, A. Hurtado, D. Labuda, W. Klitz, R. Barrantes, M. Bortolini,F. Salzano, M. Petzl-Erler, L. Tsuneto, E. Llop, F.
Rothhammer, L. Excoffier, M. Feldman, N. Rosenberg & A. Ruiz-Linares. 2007. Genetic variation and population structure in Native Americans. Plos Genet. 3: 2049-2067.


{kind=link}
{kind=link}