










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.54 n.3 San José Sep. 2006
Abundancia de Diadema antillarum (Echinodermata: Echinoidea) en las costas de Venezuela
Nicida Noriega1, Sheila M. Pauls2 & Carlos del Mónaco2
1 Departamento de Estudios Ambientales, Universidad Simón Bolívar, Apdo. Postal. 89000, Caracas, Venezuela; nicidanoriega@gmail.com
2 Instituto de Zoología Tropical, Universidad Central de Venezuela, Apdo. Postal 47058, Caracas 1041-A, Venezuela; smarquez@strix.ciens.ucv.ve; carlosdelmonaco@gmail.com
Recibido 23-VII-2005. Corregido 20-III-2006. Aceptado 23-V-2006.
Abstract: Abundance of Diadema antillarum (Echinodermata: Echinoidea) in the coasts of Venezuela. Diadema antillarum is a shallow-water sea-urchin from the tropical Atlantic whose populations almost disappeared in 1983-84 because of widespread mortalities which reached 87-100 %. In Venezuela, urchin population densities before the mortality event were comparable to those of other Caribbean regions; however, later abundancies remain unknown. The objectives of this study were to evaluate the recent densities of certain D. antillarum populations along the Venezuelan coasts and compare the densities at the Parque Nacional Mochima before and after the mortality. At each location urchin densities were determined by means of transects using 1m2-quadrats as sampling units. The highest mean densities were observed at the sites on the central coast: Ensenada de Oricao, 0.28 ind/m2 (2002) and 1.05 ind/m2 (2003), and Chichiriviche de la Costa, 0.84 ind/m2 (2002) and 0.74 ind/m2 (2003). In Mochima, the mean density before the mortality for D. antillarum oscillated between 0.28 and 4 ind/m2, after the mortality event the mean density varied between 0.15 ind/m2 (2000) and 0.47 ind/m2 (2000). The populations of D. antillarum studied at Parque Nacional Morrocoy and Refugio de Fauna Silvestre Cuare showed highest densities at Playuela (0.43 ind/m2) and Cayo Sur (0.95 ind/m2) respectively, whereas other sites showed densities below 0.1 ind/m2. The density registered at Playuela in 2003 is lower than that reported before the mortality event (0.58-3.64 ind/m2). The density for Parque Nacional Archipiélago de Los Roques, specifically for the Arrecife de Herradura remained constant between 2002 and 2003 with values between 0.22-0.23 ind/m2 respectively. To conclude, the sea urchin abundancies observed at most of the Venezuelan coastal sites that we studied were higher than those reported for other areas of the northern Caribbean, even though the values have not yet returned to those preceding the 1984 mass-mortality event, due to the slow recovery of the populations. Rev. Biol. Trop. 54 (3): 793-802. Epub 2006 Sept. 29.
Key words: sea urchin, Diadema antillarum, population density, Venezuela, Caribbean.
Diadema antillarum Philippi es un erizo de aguas someras del Atlántico tropical que habita comunidades coralinas, aunque puede ser encontrado en ambientes rocosos, raíces de manglar, praderas de fanerógamas marinas y fondos arenosos (Randall et al. 1964). Fue uno de los organismos más comunes y conspicuos de los arrecifes coralinos del Caribe, con densidades que alcanzaban hasta 23 ind/m2 (Scoffin et al. 1980). Las observaciones de campo y manipulaciones de sus densidades produjeron evidencias de que D. antillarum altera la microestructura del substrato debido a sus hábitos alimenticios, controlando principalmente la abundancia de algas, reclutas de coral y otros organismos sésiles (Sammarco 1980, Liddell y Ohlhorst 1986).
Las poblaciones de D. antillarum del Caribe y del Atlántico oeste sufrieron una mortandad masiva que empezó en enero de 1983 y duró hasta febrero de 1984. Dichas poblaciones fueron prácticamente diezmadas con mortandades de hasta un 87-100 % (Lessios 1988a, Carpenter 1990), mientras que un segundo evento ocurrido en octubre de 1985, aunque menos severo, redujo todavía más las poblaciones sobrevivientes del primer evento. Posteriormente ocurrieron otros eventos de menor intensidad entre 1990 y 1992, afectando áreas muy localizadas al norte del Caribe. El agente que causó la mortalidad fue específico para D. antillarum y se dispersó por todo el Caribe con las corrientes superficiales dominantes; actualmente dicho patógeno todavía no ha sido identificado, aunque todo indica que puede ser una bacteria o un virus (Lessios et al. 1984, Bauer y Agerter 1987).
Después de la mortalidad de Diadema, en las comunidades donde el erizo era común, la cobertura y biomasa de las algas de aguas someras aumentó inmediatamente. Sin embargo, en las comunidades donde el erizo presentaba bajas densidades los cambios fueron menos evidentes (Ruyter y Breeman 1986, Hughes et al. 1987, Carpenter 1988, Morrison 1988, Forcucci 1994, Hughes 1994). En las comunidades coralinas ocurrieron cambios en la composición y abundancia de las especies de algas, pues las macroalgas pasaron a dominar sobre las algas filamentosas y costrosas coralináceas en la mayoría de los arrecifes. El crecimiento excesivo de las algas redujo la tasa de reclutamiento de coral y provocó una disminución de la cobertura coralina (Carpenter 1990).
Luego de varios años de la desaparición de Diadema se ha observado una lenta recuperación de las poblaciones en muchas localidades, por lo cual no han alcanzado todavía las densidades anteriores a la mortalidad. En Panamá, 10 años después de este evento, la densidad de Diadema era menor del 0.3 % de los niveles pre-mortalidad (Lessios 1988b, Lessios 1995) y 20 años después era de 6.5 % (Lessios 2005). En Costa Rica, Alvarado et al. (2004) registraron densidades de 0.7 ind/m2 en 1999 y 0.3 ind/m2 en el año 2003. En los cayos de Florida (EEUU) las densidades máximas registradas fueron de 0.05 ind/m2 en 1999 y 0.03 ind/m2 en el 2000, mientras que en Dry Tortugas las abundancias alcanzaron hasta 0.8 ind/m2 (Chiappone et al. 2001, 2002). En Jamaica se ha registrado la mayor recuperación de todo el Caribe con densidades de hasta 5-12 ind/m2 (Cho y Woodley 2000, Edmunds y Carpenter 2001).
En Venezuela, Diadema se distribuía por toda la costa continental e islas oceánicas asociada principalmente a las comunidades coralinas (Zoppi 1967, Pauls 1982, Weil et al. 1984). Las abundancias de D. antillarum registradas antes de la mortalidad de 1984 en algunas localidades de las costas venezolanas, como en la Isla La Orchila (Urich 1977), Bahía de Mochima (Pauls 1982), Parque Nacional Morrocoy (Weil et al. 1984) y Punta Morón (Losada et al. 1988) eran similares a las encontradas en el resto del Caribe. Para finales de noviembre de 1984 las poblaciones de este erizo también fueron diezmadas en el país (Lessios et al. 1984). Sin embargo, a diferencia de otras zonas del Caribe, en Venezuela no hay registros de cual fue el grado de afectación de las mismas y tampoco se conocen las abundancias después del evento de mortalidad. Posterior a este evento sólo existe la evaluación realizada por Noriega (1998) en dos localidades de la Bahía de Mochima. Por lo tanto, los objetivos del presente estudio fueron evaluar la densidad reciente de algunas poblaciones de D. antillarum a lo largo de las costas de Venezuela y comparar las densidades de la Bahía de Mochima antes y después de la mortandad. Ambos objetivos fueron llevados a cabo mediante la recopilación de diversos estudios realizados por los autores.
Materiales y métodos
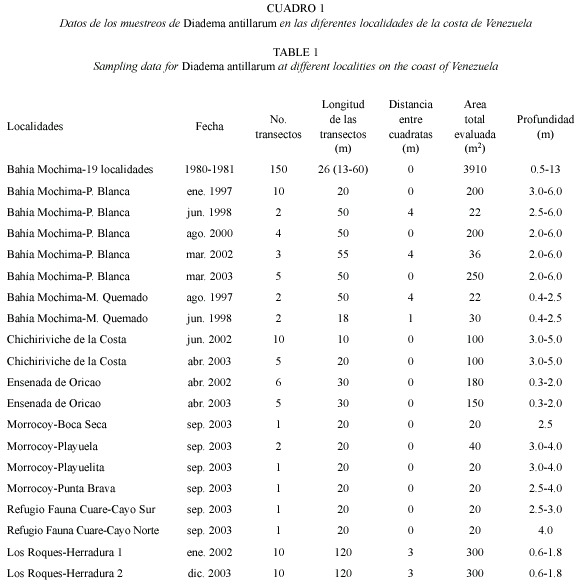
Se evaluaron un total de seis localidades de la costa de Venezuela, de las cuales cinco se ubican en la costa continental (Parque Nacional Mochima, Ensenada de Oricao, Ensenada de Chichiriviche de la Costa, Parque Nacional Morrocoy y Refugio de Fauna Silvestre Cuare) y una en el área oceánica (Parque Nacional Archipiélago Los Roques) (Fig. 1). La selección de las localidades se basó en el estudio de arrecifes y comunidades coralinas ubicados principalmente en parques nacionales y otras áreas donde actualmente existe Diadema. La Bahía de Mochima, ubicada en el Parque Nacional Mochima, se encuentra conformada por comunidades coralinas asociadas a fondos rocosos que alcanzan una profundidad de aproximadamente 10 m (Pauls 1982). Está localizada en la costa oriental del país (10º20-10º24 N, 64º1930"-64º2230" W), y posee aproximadamente 7.6 km de largo por 0.3 a 3.5 km de ancho (Okuda et al. 1968). Oricao es una pequeña ensenada ubicada en la costa central (10º3323" N-67º11 W) en las cercanías de la desembocadura del río Oricao y resguardada por rompeolas artificiales. El fondo es arenoso entremezclado con parches rocosos y alcanza una profundidad máxima de aproximadamente 4 m. La Ensenada de Chichiriviche de la Costa, también localizada en la costa central (10º3314"-10º3330" N, 67º1408"-67º1424" W), posee una dimensión aproximada de 450 x 300 m y presenta una playa arenosa con márgenes rocosos. El Parque Nacional Morrocoy y el Refugio de Fauna Silvestre Cuare se localizan en la costa occidental (10º43-10º59 N, 68º10-68º22W) y están formados por una serie de cayos coralinos que se extienden hasta una profundidad máxima de 18 m. Las áreas evaluadas en Morrocoy fueron: Punta Brava, Playuela, Playuelita y Boca Seca, además de Cayo Norte y Cayo Sur en el Refugio de Fauna Silvestre Cuare. El Parque Nacional Archipiélago Los Roques se encuentra aproximadamente a 166 km al norte de la costa central (11º4426"-11º5936" N, 66º3325"-66º5726" W). En esta localidad fueron evaluadas dos áreas distintas ubicadas en el Arrecife de Herradura, el cual se encuentra localizado entre los cayos Dos Mosquises Norte y Sur al suroeste del archipiélago. Dicho arrecife presenta una forma de arco y posee una plataforma coralina somera entre 0.1-1.3 m de profundidad.

Se realizaron muestreos diurnos en distintas fechas, evaluándose áreas que oscilaron entre 22 y 565 m2 (Cuadros 1 y 2). Para ello se colocaron transectos con una longitud variable que dependió de la extensión de la comunidad coralina o rocosa presente, así como de la distribución de las poblaciones de Diadema. Los erizos fueron cuantificados utilizándose como unidad muestreal un cuadrante de 1 m2, el cual fue colocado a lo largo del transecto, distanciados en función del tamaño del área evaluada y las características de la comunidad coralina o arrecife (Cuadro 1). La densidad de Diadema fue referida como el número de erizos por m2. No se realizaron análisis estadísticos debido a la diferencia en el tamaño de las áreas evaluadas y de las metodologías utilizadas, dado a que el presente trabajo fue una recopilación de diversos estudios realizados.
Resultados
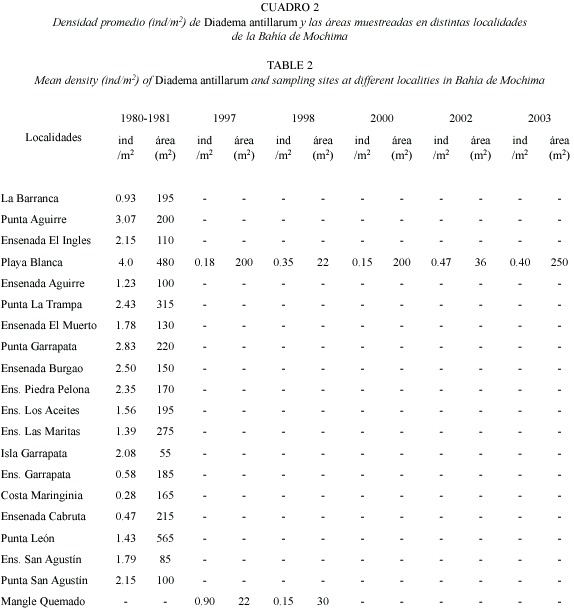
Las evaluaciones realizadas entre 1980 y 1981 en 19 áreas de la Bahía de Mochima arrojaron densidades promedio que oscilaron entre 0.28 y 4 ind/m2. De éstas, nueve áreas ubicadas en zonas cercanas a la boca de la bahía presentaron abundancias superiores a 2 ind/m2, siendo Playa Blanca el área con mayor densidad (promedio de 4 ind/m2) (Cuadro 2). Evaluaciones posteriores al evento de mortalidad de 1984 en Playa Blanca arrojaron densidades por debajo de las registradas antes de la mortalidad, con valores promedio entre 0.15 ind/m2 (2000) y 0.47 ind/m2 (2002), y una densidad máxima de 4 ind/m2 (2003) (Cuadros 2 y 3), distribuidos entre 2 y 6 m de profundidad. Por otra parte, en Mangle Quemado se presentó una densidad de 0.9 ind/m2 en 1997 y 0.15 ind/m2 en 1998, localizados entre 0.4 y 2.5 m de profundidad (Cuadro 2) formando agregaciones entre rocas y colonias de corales de Millepora alcicornis, Porites astreoides y Diploria strigosa.
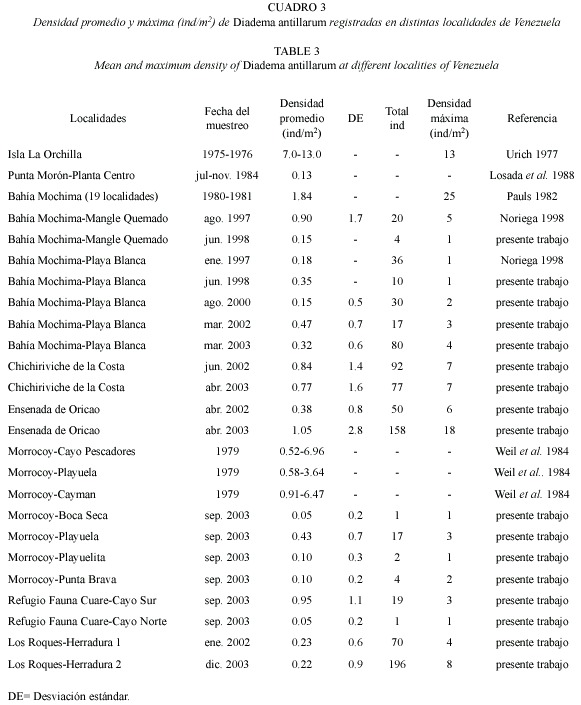
La Ensenada de Oricao, presentó la mayor densidad de Diadema, con un valor promedio de 0.28 ind/m2 en el 2002 y una densidad máxima de 6 ind/m2, mientras que en el 2003 la densidad promedio aumentó a 1.05 ind/m2 y la densidad máxima incrementó a 18 ind/m2 (Cuadro 3). La Ensenada de Chichiriviche de la Costa presentó en el 2002 una densidad promedio de 0.84 ind/m2 y un máximo de 7 ind/m2, mientras que en el 2003 el promedio fue de 0.77 ind/m2 y el máximo de 7 ind/m2 (Cuadro 3). Estos organismos fueron observados agrupados en cavidades entre rocas y corales.
En el Parque Nacional Morrocoy y en el Refugio de Fauna Silvestre Cuare, Diadema fue localizada entre 2.5 y 4.0 m de profundidad en las seis áreas evaluadas. Las máximas densidades promedio fueron de 0.43 ind/m2 en Playuela y 0.95 ind/m2 en Cayo Sur, mientras que en las demás áreas las abundancias fueron menores a 0.1 ind/m2 (Cuadro 3).
En el Arrecife Herradura (Parque Nacional Archipiélago de Los Roques) D. antillarum fue encontrada formando agregaciones en la plataforma del arrecife, cercanas a la pendiente, entre los 0.6 y 1.8 m de profundidad y localizadas principalmente entre colonias de coral de Millepora y Acropora palmata. En el 2002 la densidad promedio para el área Herradura 1 fue de 0.23 ind/m2 con un máximo de 4 ind/m2 y en el 2003 en Herradura 2 la abundancia fue de 0.22 ind/m2 con un máximo de 8 ind/m2 (Cuadro 3).
Discusión
Las abundancias del erizo registradas en los últimos años en la localidad de Playa Blanca (Bahía de Mochima) todavía son bajas en comparación con la densidad promedio anterior a la mortalidad. Esta situación podría ser interpretada como fluctuaciones naturales de las poblaciones o como una lenta recuperación de la población debido al ligero incremento en sus densidades máximas.
En las otras localidades costeras evaluadas, la abundancia de Diadema fue muy variable, lo que podría deberse principalmente a las diferencias existentes en las distintas áreas en cuanto al tipo y composición del sustrato, complejidad topográfica, parámetros físico-químicos del agua, hidrodinámica (corrientes y oleaje), abundancia y flujo de larvas, entre otros factores.
La ensenada de Oricao registró un notorio incremento en la densidad promedio de Diadema aumentando de igual forma los valores máximos de abundancia. En esta localidad se registraron las mayores densidades de todas las localidades evaluadas, las cuales son similares con las registradas en Jamaica en años recientes (5-12 ind/m2) (Cho y Woodley 2000, Edmunds y Carpenter 2001). El incremento observado en Oricao podría ser debido a la incorporación de nuevos organismos a la población por efecto de migración o altas tasas de reclutamiento, dado que las evaluaciones fueron realizadas bajo las mismas condiciones de muestreo. Las densidades registradas en Chichiriviche de la Costa fueron similares a las observadas en Dry Tortugas (0.8 ind/m2), tales densidades han sido consideradas como altas por Chiappone et al. (2001).
En las áreas muestreadas en el Parque Nacional Morrocoy y Refugio de Fauna Silvestre Cuare, las densidades promedio todavía son muy bajas, con abundancias por debajo de 0.1 ind/m2. Una posible explicación de estos bajos valores de densidad podría ser debido a una baja densidad de larvas en el plancton aunado al deterioro de los arrecifes y a las altas coberturas de algas, las cuales podrían inhibir el reclutamiento de los erizos como lo ha demostrado Bak (1985).
En las islas oceánicas como Los Roques, los arrecifes coralinos son estructuralmente complejos con una alta heterogeneidad espacial, por lo que se esperaría encontrar una mayor densidad de D. antillarum, no obstante en ambas áreas del Arrecife Herradura, la densidad promedio fue inferior a la mayoría de las localidades costeras evaluadas. En esta localidad las densidades máximas fueron observadas en la plataforma del arrecife, donde dominan corales ramificados (A. palmata y M. alcicornis) que son de importancia para el abrigo y protección de los erizos. Además, este sector del arrecife es somero y está bajo la influencia de un oleaje moderado, condiciones que son consideradas significativas para el establecimiento de Diadema (Ogden y Carpenter 1987).
Antes del evento de mortalidad masiva de D. antillarum, las densidades registradas en Venezuela eran similares a las de otras regiones del Caribe. Para la Isla La Orchila los registros señalan entre 7-13 ind/m2 para la plataforma arrecifal y menos de 5 ind/m2 para el talud (Urich 1977) y para el Parque Nacional Morrocoy Weil et al. (1984) informaron densidades de 2.6 a 3.7 ind/m2 en el año 1979. En Punta Morón (Golfo Triste), sobre los muros de contención del canal de toma de la Planta Termoeléctrica del Centro, la población de Diadema cuantificada entre julio y noviembre de 1984 alcanzó una densidad promedio de 0.13 ind/m2 (Losada et al. 1988). Los valores de densidad informados por todos los autores mencionados anteriormente fueron mayores a los registrados después de la mortandad en este estudio en toda la costa venezolana incluyendo el Parque Nacional Morrocoy.
En el Caribe en general se observa una lenta recuperación de las poblaciones de Diadema, con densidades bajas en la mayoría de las localidades estudiadas. Forcucci (1994) registró densidades de 0.02 a 0.6 ind/m2 en cuatro arrecifes estudiados en los cayos de Florida en 1990, las cuales fueron las mayores registradas después de la mortalidad de 1984 para la región. En 1991 hubo un nuevo evento de mortalidad local y las densidades disminuyeron a 0.03 ind/m2. Posteriormente, en 1999-2000, en los mismos cayos de Florida se evaluaron 125 localidades, registrándose Diadema en sólo 24 de ellas, con densidades menores a 0.05 ind/m2 (Chiappone et al. 2001, 2002). En Cuba, en los arrecifes al oeste de Cayo Largo del Sur se registraron en 1999 densidades de 0.04 y 0.19 ind/m2 (Alcolado et al. 2001).
Por otro lado, existen islas donde la recuperación ha sido más rápida como en Barbados donde las poblaciones de Diadema se habían recuperado hasta en un 57 % de los niveles pre-mortalidad para 1985 (Hunte y Younglao 1988). En Saint Croix (Islas Vírgenes) evaluaciones realizadas entre 2000 y 2001 arrojaron densidades promedio de 0.004-0.37 ind/m2 en el arrecife y de 0.24-0.52 ind/m2 en parches arrecifales (Miller et al. 2003). En Jamaica han ocurrido los mayores incrementos poblacionales, por ejemplo, en 1992 existían pequeñas agregaciones de Diadema y para 1995-96 las densidades locales alcanzaron hasta 1.8 ind/m2 (Woodley 1999, Woodley et al. 1999), en 1997 se registraron densidades de 0.1 a 3.4 ind/m2 con un promedio de 1.54 ind/m2 (Cho y Woodley 2000) y en el 2000 se registraron densidades promedio alrededor de 5 ind/m2 y en algunas áreas hasta 12 ind/m2 (Edmunds y Carpenter 2001).
Las densidades registradas recientemente en la mayoría de las localidades de Venezuela son superiores a las informadas en las áreas evaluadas al norte del Caribe, exceptuando la informada por Woodley 1999, Woodley et al. 1999 en Jamaica; sin embargo todavía no se han alcanzado los niveles anteriores a la mortalidad de 1984. Es importante señalar que en algunas localidades de la costa central como Oricao y Chichiriviche de la Costa los incrementos poblacionales, en los últimos años, se han acentuado. Debido a la importancia ecológica de D. antillarum y a la gran relevancia de las localidades estudiadas, es necesario no solo incrementar la intensidad de los monitoreos sino abarcar otras áreas a lo largo de la costa venezolana. Estas evaluaciones permitirán predecir posibles cambios en la dinámica poblacional de estos organismos.
Agradecimientos
Se agradece al Instituto Nacional de Parques (INPARQUES) y a la Fundación Científica Los Roques por el apoyo técnico en el Parque Nacional Archipiélago Los Roques. A los revisores anónimos por los comentarios y observaciones realizadas al manuscrito.
Resumen
Diadema antillarum es un erizo de aguas someras del Atlántico tropical cuyas poblaciones fueron diezmadas entre los años 1983-1984 con mortandades de hasta 87-100 %. En Venezuela, las densidades registradas antes de la mortalidad presentaban valores similares a los de otras regiones del Caribe, sin embargo se desconoce las abundancias posteriores al evento. Los objetivos del presente estudio fueron evaluar las densidades recientes de algunas poblaciones de D. antillarum a lo largo de las costas de Venezuela y comparar las densidades de la Bahía de Mochima antes y después de la mortandad. Se utilizaron transectos, y cuadrantes de 1m2 como unidad muestreal. Se registraron las mayores densidades promedio en las localidades del litoral central: Ensenada de Oricao con 0.28 ind/m2 (2002) y 1.05 ind/m2 (2003), y Chichiriviche de la Costa con 0.84 ind/m2 (2002) y 0.74 ind/m2 (2003). En la Bahía de Mochima, la densidad promedio antes de la mortandad de D. antillarum osciló entre 0.28 y 4 ind/m2, luego del evento de mortandad la densidad promedio varió entre 0.15 ind/m2 (2000) y 0.47 ind/m2 (2002). Las poblaciones de D. antillarum estudiadas en el Parque Nacional Morrocoy y Refugio de Fauna Silvestre Cuare presentaron las densidades promedio más altas en Playuela (0.43 ind/m2) y Cayo Sur (0.95 ind/m2), mientras que las demás áreas presentaron densidades inferiores a 0.1 ind/m2. La densidad registrada en Playuela en el 2003 es menor a la informada antes del evento de mortandad (0.58-3.64 ind/m2). La densidad del Parque Nacional Archipiélago Los Roques, específicamente en el Arrecife de Herradura, se mantuvo constante durante 2002-2003, con valores entre 0.22 y 0.23 ind/m2 respectivamente. En conclusión, las abundancias registradas en la mayoría de las localidades evaluadas en Venezuela son superiores a las informadas para áreas al norte del Caribe, aunque todavía no se ha alcanzado los niveles anteriores a la mortalidad de 1984 por el lento incremento de las poblaciones.
Palabras clave: erizo de mar, Diadema antillarum, densidad de población, Venezuela, Caribe.
Referencias
Alcolado, P.M., R. Claro, B. Martínez, G. Menéndez, P. García, K. Cantelar, M. Hernández & R. del Valle. 2001. Evaluación ecológica de los arrecifes coralinos del oeste de Cayo Largo del Sur, Cuba: 1998-1999. Bol. Invest. Mar. Cost. 30: 109-132. [ Links ]
Alvarado, J.J., J. Cortes & E. Salas. 2004. Population densities of Diadema antillarum Philippi at Cahuita National Park (1977-2003) Costa Rica. Carib. J. Sci. 40: 257-259. [ Links ]
Bak, R.P.1985. Recruitment patterns and mass mortalities in the sea urchin Diadema antillarum . Proc. 5th Int. Coral Reef Congress 5: 267-272. [ Links ]
Bauer, J.C. & C.J. Agerter. 1987. Isolation of bacteria pathogenic for the sea urchin Diadema antillarum (Echinodermata: Echinoidea). Bull. Mar. Sci. 40: 161-165. [ Links ]
Carpenter, R.C. 1988. Mass mortality of a Caribbean sea urchin: immediate effects on community metabolism and other herbivores. Proc. Natl. Acad. Sci. USA 85: 511-514. [ Links ]
Carpenter, R.C. 1990. Mass mortality of Diadema antillarum. I. Long-term effects on sea urchin population-dynamics and coral reef algal communities. Mar. Biol. 104: 67-77. [ Links ]
Chiappone, M., S.L. Miller, D.W. Swanson, J.S. Ault & S.G. Smith. 2001. Comparatively high densities of the long-spined sea urchin in the Dry Tortugas, Florida. Coral Reefs 20: 137-138. [ Links ]
Chiappone, M., D.W. Swanson & S.L. Miller. 2002. Density, spatial distribution and size structure of sea urchins in Florida Keys coral reef and hard-bottom habitats. Mar. Ecol. Prog. Ser. 235: 117-126. [ Links ]
Cho, L.L. & J.D. Woodley. 2000. Recovery of reefs at Discovery Bay, Jamaica and the role of Diadema antillarum. Proc. 9th Int. Coral Reef Symp. 1: 331-337. [ Links ]
Edmunds, P.J. & R.C. Carpenter. 2001. Recovery of Diadema antillarum reduces macroalgal cover and increases abundance of juvenile corals on a Caribbean reef. Proc. Nat. Acad. Sci. USA 98: 5067-5071. [ Links ]
Forcucci, D. 1994. Population density, recruitment and 1991 mortality event of Diadema antillarum in the Florida Keys. Bull. Mar. Sci. 54: 917-928. [ Links ]
Hughes, T.P. 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265: 1547-1551. [ Links ]
Hughes, T.P, D.C. Reed & M.J. Boyle. 1987. Herbivory on coral reefs: community structure following mass mortalities of sea urchins. J. Exp. Mar. Biol. Ecol. 113: 39-59. [ Links ]
Hunte, W. & D. Younglao. 1988. Recruitment and population recovery of Diadema antillarum (Echinodermata, Echinoidea) in Barbados. Mar. Ecol. Prog. Ser. 45: 109-119. [ Links ]
Lessios, H.A. 1988a. Mass mortality of Diadema antillarum in the Caribbean: what have we learned? Ann. Rev. Ecol. Syst. 19: 371-393. [ Links ]
Lessios, H.A. 1988b. Population dynamics of Diadema antillarum (Echinodermata: Echinoidea) following mass mortality in Panama. Mar. Biol. 99: 515-526. [ Links ]
Lessios, H.A. 1995. Diadema antillarum 10 years after mass mortality: still rare, despite help from a competitor. Proc. R. Soc. Lond. B 259: 331-337. [ Links ]
Lessios, H.A. 2005. Diadema antillarum populations in Panama twenty years following mass mortality. Coral Reefs 24: 125-127. [ Links ]
Lessios, H.A., J.D. Cubit, D.R. Robertson, M.J. Shulman, M.R. Parker, S.D. Garrity & S.C. Levings. 1984. Mass mortality of Diadema antillarum on the Caribbean coast of Panama. Coral Reefs 3: 173-182. [ Links ]
Liddell, W.D. & S.L. Ohlhorst. 1986. Changes in benthic community composition following the mass mortality of Diadema at Jamaica. J. Exp. Mar. Biol. Ecol. 95: 271-278. [ Links ]
Losada, F., A. Martín, W. Feragotto & C. Alamo. 1988. Interacciones biológicas en el canal de toma de la Planta Termoeléctrica del Centro en Punta Morón, Venezuela. Ecotropicos 1: 55-70. [ Links ]
Miller, R.J., A.J. Adams, N.B. Ogden, J.C. Ogden & J.P. Ebersole. 2003. Diadema antillarum 17 years after mass mortality: is recovery beginning on St. Croix? Coral Reefs 22: 181-187. [ Links ]
Morrison, D. 1988. Comparing fish and urchin grazing in shallow and deeper coral reef algal communities. Ecology 69: 1367-1382. [ Links ]
Noriega, N. 1998. Caracterización de las comunidades de erizos presentes en praderas de Thalassia testudinum y comunidades coralinas de la Bahía de Mochima, Parque Nacional Mochima. Trabajo de Grado, Universidad Central de Venezuela, Caracas, Venezuela. 97 p. [ Links ]
Ogden, J.C. & R.C. Carpenter. 1987. Species profiles: Life histories and environmental requirements of coastal fishes and invertebrates (South Florida). Long-spined black sea urchin. U.S. Fish Wildl. Serv. Biol. Rep. 82: 1-17. [ Links ]
Okuda, T., J. Benitez, A.J. García & E. Fernández. 1968. Condiciones hidrográficas y químicas en la Bahía de Mochima y la Laguna Grande del Obispo desde 1964 a 1966. Bol. Inst. Oceanogr. Univ. Oriente 7: 7-37. [ Links ]
Pauls, S.M. 1982. Estructura de las comunidades coralinas de la Bahía de Mochima, Venezuela. Tesis Maestría, Universidad de Oriente, Cumaná, Venezuela, 124 p. [ Links ]
Randall, J.E., R.E. Schroeder & W.A. Starck. 1964. Notes on the biology of the echinoid Diadema antillarum. Carib. J. Sci. 4: 421-433. [ Links ]
Ruyter van Steveninck, E. & A.M. Breeman. 1986. Deep water vegetation of Lobophora variegata (Phaeophyceae) in the coral reef of Curacao: populations dynamics in relation to mass mortality of the sea urchin Diadema antillarum. Mar. Ecol. Prog. Ser. 36: 81-90. [ Links ]
Sammarco, P.W. 1980. Diadema and its relationship to coral spat mortality: grazing, competition, and biological disturbance. J. Exp. Mar. Biol. Ecol. 45: 245-272. [ Links ]
Scoffin, T.P., C.W. Stearn, D. Boucher, P. Frydl, C.M. Hawkins, I.G. Hunter & J.K. MacGeachy. 1980. Calcium carbonate budget of a fringing reef on the west coast of Barbados. Part. 2. Erosion, sediments and internal structure. Bull. Mar. Sci. 30: 475-508. [ Links ]
Urich, J.F. 1977. Estudio de la estructura comunitaria de un arrecife coralino al sur-oeste de la Orchila. Tesis de Grado, Universidad Central de Venezuela, Caracas, Venezuela, 157 p. [ Links ]
Weil, E., F. Losada & D. Bone. 1984. Spatial variations in density and size of the echinoid Diadema antillarum Philippi on some Venezuelan coral reefs. Bijdragen tot de Dierkunde 54: 73-82. [ Links ]
Woodley, J.D. 1999. Sea-urchins exert top-down control of macroalgae on Jamaican coral reefs (1). Coral Reefs 18: 192. [ Links ]
Woodley, J.D., P.M. Gayle & N. Judd. 1999. Sea-urchins exert top-down control of macroalgae on Jamaican coral reefs (2). Coral Reefs 18: 193. [ Links ]
Zoppi, E. 1967. Contribución al estudio de los equinodermos de Venezuela. Acta Biol. Venezuelica 5: 267-333. [ Links ]

