










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.51 n.2 San José Jun. 2003
Apolinar Santamaría Miranda 1, Juan F. Elorduy Garay 2 & Agustín A. Rojas Herrera 3
Recibido 02-V-2001. Corregido 12-IX-2002. Aceptado 08-X-2002.
Abstract
In this paper we describe the feeding habits of the Red snapper (Lutjanus peru) in the southern coast of Guerrero state, Mexico. This data could be relevant to local fisheries management. In total 385 full stomachs (206 from females and 179 from males) were collected monthly in 1993 and 1994, from commercial catches. Stomach contents were identified to species level when possible. Food items were grouped into taxonomically coherent groups, and their numbers, weight and frequency of occurrence obtained. The index of relative importance (IRI) was calculated for the total sample, by month, sex and size class. In 1993, a total of 42 different food items were obtained, and 43 in 1994. Food items consisted of fishes, crustaceans, and mollusks. The grouping of prey species, genera, or families rendered 14 food groups, as follows: Mollusks, Amphipods, Copepods, Stomatopods, Penaeus vannamei, Brachiuran Larvae, Stomatopods Larvae, Family Albuneidae Larvae, Other crustaceans, Anchoa ischana, Anchoa lucida, Other fishes, Salps and Unidentified organic matter. Anchoa ischana dominated the trophic spectrum in both years. Although, Amphipods, Anchoa lucida and Other fishes had important indexes in 1994. There was high variability in the diet composition by month. Nevertheless, in 1993, from February to August, crustaceans as a whole dominated the diet. In 1994, crustaceans only dominated in March and August. No differences in food habits were found between immature fish, males and females. The diet of the Red snapper varies as they grow, although the same food groups are found always. At the adult phase, in length classes over 261 mm FL, there is some degree of specialization, as they consume more fish, particularly engraulids.
Key words: Lutjanus peru, feeding habits, Numeric Index, Gravimetric Index, Index of Relative Importance, West central Mexico.
La actividad pesquera es un renglón muy importante en la economía local y regional del estado de Guerrero. Las capturas comerciales más importantes de las pesquerías ribereñas se basan fundamentalmente en peces de las familias Lutjanidae, Carangidae, Haemulidae y Ariidae. Entre éstas destacan, por su valor y volumen de captura, las especies de Lutjanidae Lutjanus peru (1 000 tm al año) y Lutjanus guttatus (650 tm al año), y las de Carangidae Caranx caballus (1 000 tm al año) y Selar crumenophthalmus (550 tm al año) (Rojas-Herrera 1990).Los miembros de la familia Lutjanidae son depredadores con hábitos alimentarios diferentes, según determinaron Polovina y Ralston (1987). Todos son carnívoros, principalmente ictiófagos, aunque consumen una gran variedad de organismos bentónicos, entre ellos los crustáceos (Randall 1967). Por otro lado, sin embargo, Sierra y Popova (1997) mencionan que los juveniles de cinco especies de pargos del Golfo de Nicoya, Costa Rica, presentan una alta similitud en la composición de sus dietas.Rojas (1997b) estableció que en la dieta del pargo colorado L. colorado en el Golfo de Nicoya, Costa Rica, se presentan 16 especies de peces y 14 de crustáceos, así como moluscos, poliquetos y materia vegetal, observando diferencias ontogénicas.
En el Pacifico sur de México, Santamaría-Miranda y Rojas-Herrera (1994b) observaron un descenso marcado de la intensidad de alimentación al inicio de la actividad reproductiva de L. guttatus en la costa del estado de Guerrero. Santamaría-Miranda y Rojas-Herrera (1994a) encontraron una estrecha relación entre la alimentación y la actividad reproductiva de L. peru en la costa del estado de Guerrero. Díaz-Uribe (1994) determinó que L. peru, en las bahías de La Paz y la Ventana, en B.C.S., basa su alimentación en organismos planctónicos.
La revisión bibliográfica de varias fuentes demostró que el nivel de conocimiento sobre la biología, dinámica poblacional y ecología de la especie Lutjanus peru para el estado de Guerrero es mínima. Sólo existen los trabajos de Aguilar Salazar (1986) sobre edad y crecimiento; Santamaría y Rojas (1994a, b) sobre alimentación y sobre reproducción.
En los estudios de alimentación la información más importante es la determinación de los tipos de alimento consumidos, lo que proporciona los eslabones tróficos que ligan a los peces con el ecosistema, e incluso información de la presencia y abundancia de las especies que consume, por lo que constituye una base importante en el manejo de los recursos pesqueros. El objetivo del presente trabajo es describir los hábitos alimentarios de L. peru y determinar su variabilidad anual y mensual, por sexos y tallas.
Materiales y métodos
Los datos para el presente estudio corresponden a la pesquería de Punta Maldonado, al sur del estado de Guerrero, entre 17°20 y 17°32 N, y 99°08 y 99°43 W (Fig. 1).
Los ejemplares de L. peru se recolectaron con periodicidad mensual durante dos años, desde enero de 1993 hasta diciembre de 1994; sin embargo, no se pudieron tomar muestras en los meses de marzo, junio, julio y septiembre de 1993. Se revisaron 385 estómagos llenos (206 de hembras y 179 de machos). Se realizó una prueba de hipótesis (a = 0.05) sobre la proporción de sexos (M/H), asumiendo como hipótesis nula una proporción de 0.5 (Sokal y Rohlf 1979).
Donde: P es el estimador de la hipótesis nula, p la proporción de machos, q la proporción de hembras y n el tamaño de la muestra.
Los componentes alimentarios se identificaron hasta el mínimo taxon posible, con la ayuda de las claves taxonómicas de Brusca (1980), Morris et al. (1980) y Abbott (1987), para los crustáceos y moluscos; y para peces las de Thomson et al. (1979) y Fisher et al. (1995), así como la ayuda de especialistas en cada uno de los grupos. Dichos componentes se agruparon posteriormente por grupos taxonómicos coherentes, siguiendo la metodología propuesta por Elorduy-Garay y Caraveo-Patiño (1994). Se contaron los individuos presa y se pesaron en una balanza analítica. Se calcularon, para cada uno de los años, tallas y sexos, los índices: Numérico (N), Gravimétrico (W) y su Frecuencia de Aparición (FA). Con el conjunto de ellos se calculó el Índice de Importancia Relativa (IIR) (Pinkas et al. 1971). Se realizaron pruebas de órdenes con signo de Wilcoxon (a = 0.05) (Sokal y Rohlf 1979) para detectar diferencias en las dietas de juveniles, machos, hembras y los espectros alimentarios globales.
Resultados
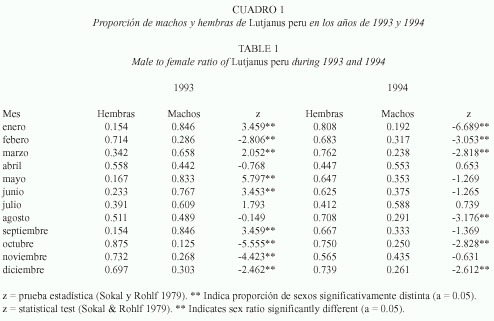
Para evaluar la proporción de sexos se consideraron 1 033 ejemplares de L. peru, de los cuales 481 fueron organismos de sexo indeterminado (denominados inmaduros de aquí en adelante), 375 hembras y 177 machos. La proporción general fue de 1:2.12 entre machos y hembras. En general se encontraron diferencias significativas en la proporción de machos y hembras, ya que sólo en nueve meses de los 24 analizados no existieron diferencias significativas. Sin embargo, no se puede asignar un dominio claro de ninguno de los sexos en 1993, aunque en 1994 dominaron las hembras durante seis meses (Cuadro 1).
El espectro alimentario de L. peru estuvo integrado por al menos 51 componentes en total, pertenecientes a 3 grandes táxones: moluscos, crustáceos y peces (Cuadro 2). Los crustáceos fueron el taxon más abundante de la dieta, seguido de los peces y los moluscos.
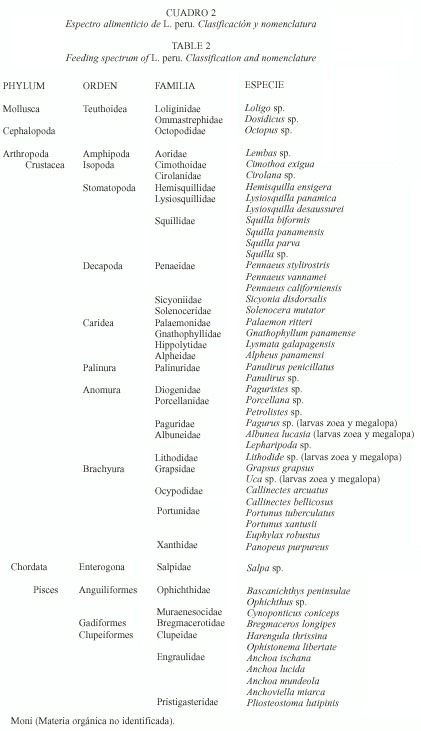
En ninguno de los casos se observaron diferencias significativas en el espectro alimentario global entre los dos años, ni entre organismos de sexo indeterminado, machos y hembras en los años considerados. Se obtuvieron 42 componentes en total para el año 1993 y 43 para el año 1994. Los componentes se agruparon en 14 grandes grupos: 1. Cefalópodos, 2. Anfípodos, 3. Copépodos, 4. Estomatópodos, 5. Penaeus vannamei, 6. Larvas de braquiuros, 7. Larvas de estomatópodos, 8. Larvas de Albuneidae, 9. Otros crustáceos, 10. Anchoa ischana, 11. Anchoa lucida, 12. Otros peces, 13. Sálpidos y 14. Moni (Materia orgánica no identificada).
Los tres componentes más importantes del año 1993 fueron: Anchoa ischana, con un índice numérico de 30% y gravimétrico de 43%, "otros crustáceos" (N = 11%, W= 15%), y las larvas de braquiuros (N = 19%, W = 2%) (Fig. 2a). Los 3 componentes más representados el año 1994 fueron: Anchoa ischana (N = 20%, W = 44%), anfípodos (N = 21%, W = 16%) y "otros peces" (N= 12%, W = 18%) (Fig. 2b).
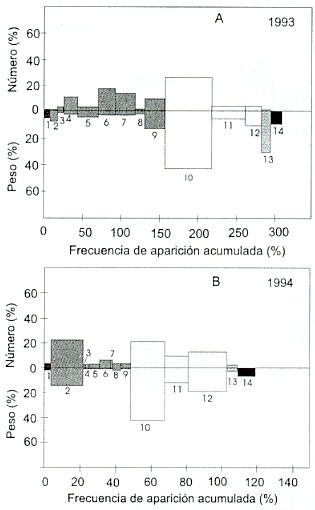
Fig. 2. Índice de Importancia Relativa global, para: A) 1993 y B) 1994.
Los números indican: 1 = Cefalópodos, 2= Anfípodos, 3 = Copépodos,
4 = Estomatópodos, 5 = Penaeus vannamei, 6 = Larvas de braquiuros,
7 = Larvas de estomatópodos, 8 = Larvas de la familia Albuneidae,
9 = Otros crustáceos, 10 = Anchoa ischana, 11 = Anchoa lucida,
12 = Otros peces, 13 = Salpas, 14 = Moni.
Fig. 2. Global Index of Relative Importance, for A) 1993, and B) 1994.
Numbers indicate: 1 = Mollusks, 2 = Amphipods, 3 = Copepods,
4 = Stomatopods, 5 = Penaeus vannamei, 6 = Brachiuran Larvae,
7 = Stomatopods Larvae, 8 = Family Albuneidae Larvae,
9 = Other crustaceans, 10 = Anchoa ischana, 11 = Anchoa lucida,
12 = Other fishes, 13 = Salps, 14 = Unidentified organic matter.
En febrero, abril, agosto y noviembre de 1993 el conjunto de crustáceos fue el más dominante; y en enero, octubre y diciembre dominaron los peces. Abril y mayo fueron los meses de mayor contenido de alimento en cuanto a crustáceos y peces, tanto en número como en peso (Fig. 3). El año 1994 se caracterizó por una presencia de crustáceos más constante, al presentarse de seis a ocho grupos de esta categoría; sin embargo, sólo en agosto dominaron en número y peso. Los peces fueron los de mayor importancia en general, a lo largo de todo el año, tanto en número como en peso (Fig. 4).
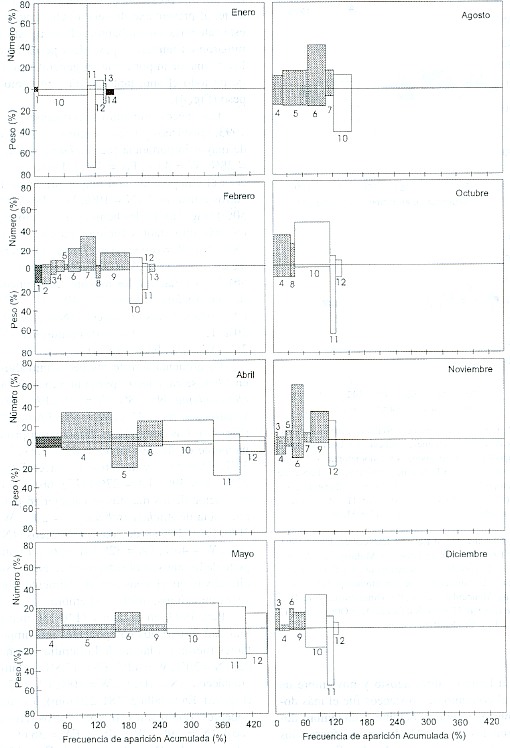
Fig. 3. Índices de Importancia Relativa, por mes, en 1993. Los números indican los mismos grupos alimentarios de la Fig. 2.
Fig. 3. Index of Relative Importance by month in 1993. Numbers indicate the same food groups as in Fig. 2.
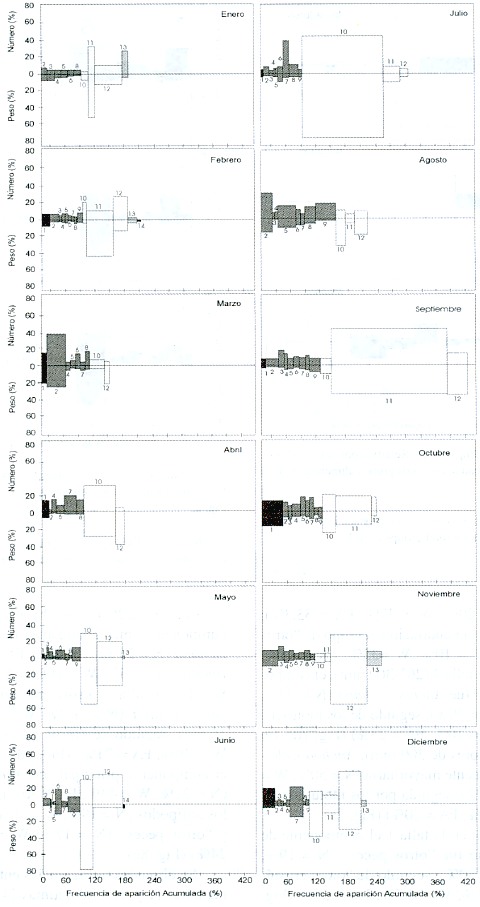
Fig. 4. Índices de Importancia Relativa, por mes, en 1994. Los números indican los mismos grupos alimentarios de la Fig. 2.
Fig. 4. Index of Relative Importance by month in 1994. Numbers indicate the same food groups as in Fig. 2.
Los peces inmaduros consumieron, en 1993, crustáceos y peces. Entre éstos, la presa de mayor importancia fue Anchoa ischana (N = 28%, W = 43%, FA = 47%). De los crustáceos el componente con mayor presencia fue "otros crustáceos" (N = 10%, W = 10%, FA =50%) (Fig. 5a). En las hembras los componentes más importantes fueron: Anchoa ischana (N = 38%, W=44%, FA = 27%) y "otros crustáceos" (N = 11%, W = 10%, FA = 12%) (Fig.5b). En los machos, los mayores componentes fueron: Anchoa ischana (N = 40%, W = 22%, FA = 30%), "otros crustáceos" (N = 10%, W = 20%, FA = 39%) y "larvas de braquiuros" (N =25%, W = 2%, FA = 14%) (Fig. 5c).
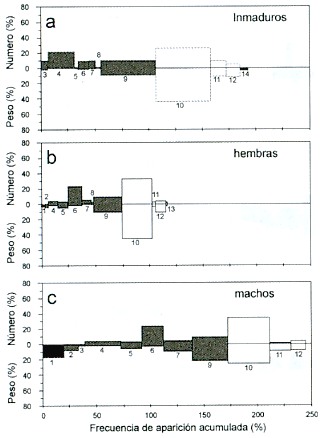
Fig. 5. Índices de Importancia Relativa, por sexo, en 1993.
Los números indican los mismos grupos alimentarios de la Fig. 2.
Fig. 5. Index of Relative Importance by sex in 1993. Numbers
indicate the same food groups as in Fig. 2.
La alimentación de los peces inmaduros, en 1994, se caracterizó por la presencia de Anchoa ischana (N = 18%, W= 35%, FA= 28%) y anfípodos (N = 21%, W = 5%, FA = 32%) (Fig. 6a). Las hembras se alimentaron principalmente de Anchoa ischana (N = 25%, W = 37%, FA = 29%) y de "otros crustáceos" (N = 22%, W = 39%, FA = 27%) (Fig. 6b). La alimentación de los machos se caracterizó por la presencia de Anchoa ischana (N = 20%, W =20%, FA = 26%) y de "otros crustáceos" (N= 20%, W = 40%, FA = 32%) (Fig. 6c). En ninguno de los años se observaron diferencias significativas en el consumo de alimento entre peces inmaduros, machos o hembras.
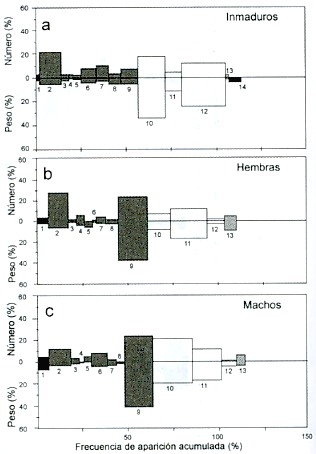
Fig. 6. Índices de Importancia Relativa, por sexo, en 1994. Los números indican
los mismos grupos alimentarios de la Fig. 2.
Fig. 6. Index of Relative Importance by sex in 1994. Numbers
indicate the same food groups as in Fig. 2.
Para 1993, en la talla 1 (140-180 mm de longitud furcal), los componentes más importantes fueron las larvas de la familia Albuneidae (N = 20%, W= 42%, FA= 17.8%) y "otros crustáceos" (N = 10.5%, W= 10%, FA = 27%) (Fig. 7a). En la talla 2 (181-220 mm), los componentes dominantes fueron "otros crustáceos" (N = 19%, W = 48%, FA = 36.6%) y estomatópodos (N= 9%, W = 8%, FA = 15%) (Fig. 7b). En la talla 3 (221-260 mm), el espectro alimenticio estuvo dominado por Anchoa ischana (N = 20%, W = 37%, FA = 33.3%); también tuvo gran aportación el grupo de "otros crustáceos" (N = 10%, W =35%, FA = 43%) (Fig. 7c). En la talla 4 (261-300 mm), el componente principal fue Anchoa ischana (N = 55%, W= 39%, FA = 40%), seguido de los moluscos (N = 10%, W = 10%, FA = 23%) (Fig. 7d). En la talla 5 (mayores de 300 mm), Anchoa ischana fue ampliamente mayoritaria (N = 50%, W= 37%, FA = 70%), seguida por los moluscos (N = 9%, W = 33%, FA = 20%) (Fig. 7e).
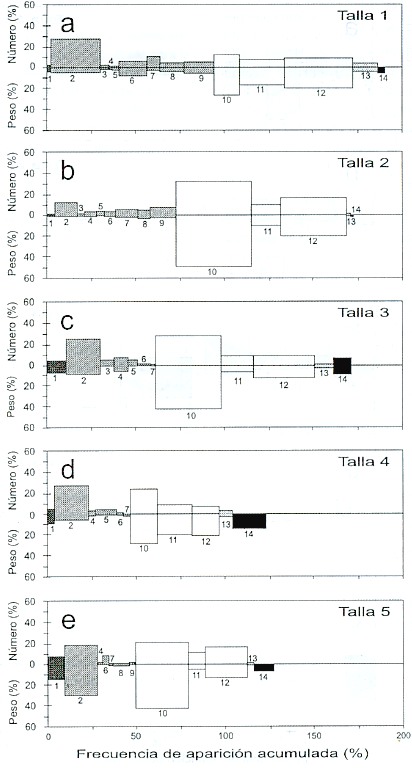
Fig. 7. Índices de Importancia Relativa, por clase de talla, en 1993. a) talla 1= 140 - 180 mm LF, b) talla 2 = 181 - 220 mm,
c) talla 3 = 221 - 260 mm, d) talla 4 = 261 - 300 mm, e) talla 5 = mayores de 300 mm. Los números indican los mismos
grupos alimentarios de la Fig. 2.
Fig. 7. Index of Relative Importance by length class in 1993. a) length 1= 140 - 180 mm FL, b) length 2 = 181 - 220 mm,
c) length 3 = 221 - 260 mm, d) length 4 = 261 - 300 mm, e) length 5 = larger than 300 mm. Numbers indicate the same
food groups as in Fig. 2.
Para 1994, en la talla 1 el componente de mayor dominio fue "otros peces" (N = 19%, W=18%, FA =50%), seguido de "otros crustáceos" (N = 27%, W = 4.5%, FA = 30% (Fig.8a). En la talla 2, los componentes más importantes fueron: Anchoa ischana (N = 31%, W=52%, FA =40%) y "otros peces" (N = 16%, W = 23%, FA = 22%) (Fig. 8b). En la talla, 3 los componentes más abundantes fueron: Anchoa ischana (N = 25%, W= 42%, FA= 30%) y anfípodos (N = 25%, W= 10%, FA = 20%) (Fig.8c). En la talla 4, estuvieron presentes: Anchoa ischana (N = 18%, W = 25%, FA = 19.6%), seguido de "otros peces" (N = 17%, W = 20%, FA = 21%) (Fig. 8d). En la talla 5, el componente principal fue Anchoa ischana (N = 21%, W = 43%, FA = 30%), seguido por los anfípodos (N = 19%, W= 30%, FA= 18%) y "otros peces" (N = 17%, W = 12%, FA =24%) (Fig. 8e).
Se observaron diferencias entre las primeras tallas (1 y 2) y las últimas (4 y 5), ya que las tallas menores consumen un mayor número de grupos de crustáceos y en las mayores disminuye el consumo de estos componentes.
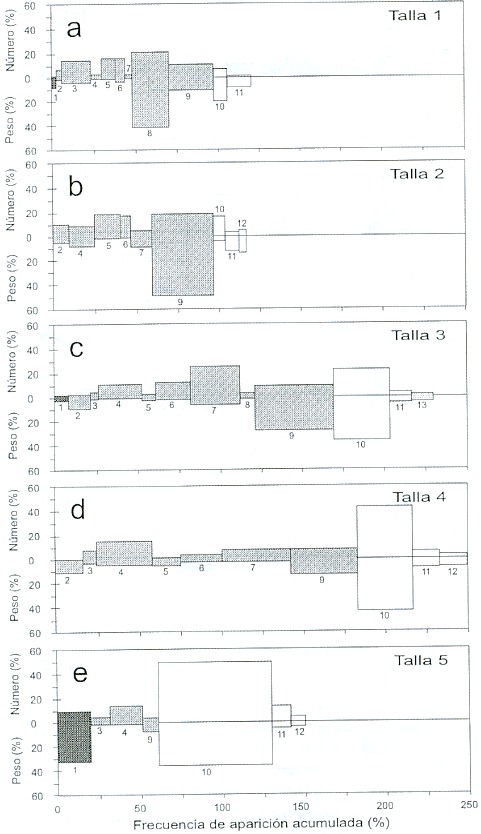
Fig. 8. Índices de Importancia Relativa, por clase de talla, en 1994. a) talla 1= 140 - 180 mm LF, b) talla 2 = 181 - 220 mm,
c) talla 3 = 221 - 260 mm, d) talla 4 = 261 - 300 mm, e) talla 5 = mayores de 300 mm. Los números indican los mismos
grupos alimentarios de la Fig. 2.
Fig. 8. Index of Relative Importance by length class in 1994. a) length 1= 140 - 180 mm FL, b) length 2 = 181 - 220 mm,
c) length 3 = 221 - 260 mm, d) length 4 = 261 - 300 mm, e) length 5 = larger than 300 mm. Numbers indicate the same
food groups as in Fig. 2.
Discusión
El predominio de las hembras encontrado en este trabajo contrasta con lo reportado para la misma especie en otras zonas del Pacífico mexicano (Cruz-Romero et al. 1988, Rocha-Olivares 1991). Sin embargo, esto se puede deber a una mayor supervivencia de las hembras, pero también podría deberse a diferencias en la distribución y hábitat de ambos sexos, como ocurre con L. griseus (Claro 1983c, Stark 1971).
Allen (1985) determinó que los pargos son depredadores activos y que se alimentan principalmente durante la noche de una gran variedad de organismos, pero fundamentalmente de peces pequeños.
Podemos señalar que el espectro alimenticio de L. peru en las aguas del litoral de Guerrero es bastante amplio, similar a otras especies de Lutjánidos (Barroso 1965, Stark 1971, Claro 1983a, b, c, Rojas-Herrera 1996, Sierra y Popova 1997). El espectro se integró de 51 componentes diferentes, pertenecientes a 3 táxones mayores: moluscos, crustáceos (importantes en número) y peces (importantes en peso).
En la dieta de cada uno de los años se obtuvieron 42 componentes en 1993 y 43 en 1994. No existe una marcada diferencia de componentes entre ambos años, a pesar de que 1993 estuvo enmarcado en un evento "El Niño". El componente más importante en los contenidos estomacales de ambos años fue Anchoa ischana; pero en 1994 también fueron importantes Anchoa lucida, "otros peces" y los anfípodos.
Los peces constituyentes de la dieta de L. peru pertenecen a las familias Engraulidae, Clupeidae, Bregmacerotidae, Ophichthidae y Muraenosocidae, siendo los engráulidos los mejor representados. Los peces de las familias Engraulidae y Bregmacerotidae también son los más abundantes en los espectros tróficos de L. guttatus y Caranx caballus, capturados en la misma región de estudio y en la misma época del año (Rojas-Herrera 1996), por lo que dichas presas pueden considerarse muy abundantes en la zona.
El grupo "otros crustáceos" fue el más importante entre los crustáceos en 1993. Incluyó a las familias Palaemonidae, Gnathophyllinae, Alpheidae, Hippolytidae, Callianasidae, Palinuridae, Lithodidae, Porcelanidae, Majidae, Xanthidae, Grapsidae y Portunidae, la mayoría de ellos crustáceos bentónicos. En 1994, los anfípodos constituyeron el grupo de crustáceos más importante, mientras que el resto de los crustáceos tuvo una representación mucho menor. La preferencia de los pargos, adultos y juveniles, por los crustáceos ha sido documentada en varias especies de Lutjanus, tales como: L. synagris, L. analis, L. aya, L. argentiventris, L. guttatus (Barroso 1965, Claro 1981a, b, Suárez y Rubio 1983, Rojas 1997b).
Los moluscos, las salpas (Ascidiacea) y la materia orgánica no identificada (Moni) tuvieron una representación muy pequeña ambos años.
Lutjanus peru de las aguas del litoral de Guerrero posee un espectro alimenticio mucho más amplio que el obtenido por Díaz-Uribe (1994) en las bahías de La Paz y la Ventana B.C.S. En éstas, L. peru se alimenta principalmente de organismos planctónicos, casi exclusivamente de colonias de urocordados (doliólidos y salpas) y, en menor proporción, de moluscos, crustáceos y peces.
Parrish (1987) menciona que L. peru presenta un carácter generalista y carnívoro-oportunista y basa su alimentación en invertebrados bentónicos. Otras especies de la familia Lutjanidae, como L. kasmira (Oda y Parrish 1981), Ocyurus chrysurus, L. analis, L. griseus (Pedroso y Anderes 1983, Sierra y Popova 1997), L. guttatus (Rojas 1997a) y L. colorado (Rojas 1997b), también basan su alimentación en crustáceos y peces.
En el análisis mensual se observaron ligeras variaciones de la dieta a lo largo de los dos años, encontrándose los mismos grupos de organismos con diferencias en número y peso. (Sierra y Popova 1989) mencionan que la intensidad de la dinámica estacional en relación con la alimentación en las especies estudiadas mantiene un carácter cíclico regular. El ritmo anual de la intensidad de alimentación de los peces está relacionado con los cambios ambientales y las alteraciones que éstos provocan sobre la base alimentaria. En la región tropical, las fluctuaciones del ciclo vegetativo no impiden que los peces se alimenten durante todo el año.
En el estado de Guerrero existen por lo general cinco meses de lluvias (junio-noviembre), con precipitaciones que llegan a alcanzar los 800 mm y siete meses de "secas" (diciembre-mayo). Por lo que consideramos que las variaciones de los componentes alimentarios a lo largo del año se deben a los cambios ambientales. Este tipo de comportamiento ha sido referido por otros auto res, para especies tropicales (Guevara et al. 1994, Rooker 1995, Sierra y Popova 1997).
Sin embargo, diversos autores mencionan no haber encontrado variaciones estacionales en la composición de los espectros alimentarios, y algunas fluctuaciones en la proporción de los diferentes componentes que constituyen las dietas de las especies por ellos estudiadas (Claro 1981a, b, 1983a, b, c, González-Sansón y Rodríguez-Viñas 1983, Sierra et al. 1994, Rojas 1997b). Esto puede ser debido a que en las regiones tropicales existe una estabilidad relativa de las condiciones climáticas durante el ciclo anual, lo que permite cierta estabilidad de la base alimentaria de los peces. A pesar de ello, no descartan la influencia que ejercen los cambios del régimen hidrometeorológico sobre las comunidades de organismos que habitan en una región (Sierra y Popova 1989). Por otro lado, Randall (1967) plantea la posibilidad de variaciones en la composición del espectro alimentario de los peces tropicales, con relación a las diferencias locales en la abundancia y disponibilidad de los organismos que constituyen la base alimentaria de los depredadores.
La alimentación en machos y hembras de L. peru es similar cualitativamente, y ligeramente diferente desde el punto de vista cuantitativo, ya que se alimentan de los mismos componentes sin que exista diferencia significativa en la dieta (prueba de órdenes con signo de Wilcoxon). Lo anterior coincide con lo encontrado por Guevara et al. (1994) en L. jocu, L. analis y L. cyanopterus, y Pedroso y Anderes (1983) en L. griseus, en el archipiélago de Los Canarreos, Cuba.
El espectro alimenticio del huachinango varía durante el crecimiento, pero se encuentran siempre los mismos grupos. Al parecer durante la etapa adulta (tallas 4 y 5) se manifiesta una especialización, ya que ingiere fundamentalmente peces, particularmente engráulidos. Todavía en las tallas 3 y 4 los crustáceos ocupan una parte importante de la dieta, aunque los índices numéricos y gravimétricos son generalmente bajos. Estos resultados concuerdan con los obtenidos por Díaz-Uribe (1994), quien concluye que los adultos de L. peru presentan una dieta más especializada que la de los juveniles, con poco sobrelapamiento entre ambas. Las pocas diferencias obtenidas en estos trabajos pueden deberse a la abundancia o disponibilidad del alimento del área donde fueron recolectados los organismos; ya que el trabajo citado se localiza en aguas templado-cálidas y el presente corresponde a aguas netamente cálidas.
Stark (1971) encontró, en los estómagos de juveniles de L. griseus de Florida, crustáceos bentónicos y camarones penéidos, principalmente. En aguas de Costa Rica, Sierra y Popova (1997) detectaron que los juveniles de cinco especies de pargos, al alcanzar el primer año de vida dirigen su alimentación hacia aquellos grupos de organismos que constituyen su base alimentaria cuando son adultos. Los organismos de L. guttatus menores de 20 cm de longitud total consumen principalmente camarones; después de los 20 cm el espectro se diversifica hacia el consumo de peces, moluscos, estomatópodos, poliquetos, equinodermos y crustáceos, hasta los 50 cm aproximadamente, después de los 52 cm su dieta está compuesta por estomatópodos (Rojas 1997a).
En L. colorado se encontró que los cambios de hábitos alimentarios, que se suceden entre los juveniles y adultos, parecen estar asociados con la disminución espacial en la capacidad depredadora cuando abandonan la zona estuarina e invaden las aguas profundas, donde la preferencia de los adultos por el consumo de crustáceos podría estar relacionada con la abundancia del recurso. Por lo tanto, la reducción del consumo de peces se compensa habitando zonas donde la abundancia natural de su presa es alta (Rojas 1997b).
En la dieta de L. apodus, en el suroeste de Puerto Rico, Rooker (1995) encontró que los juveniles mayores de 70 mm de longitud basan su alimentación casi exclusivamente de crustáceos y sipuncúlidos; a los 100 mm de longitud cambia su alimentación, consumiendo cangrejos, camarones y peces. Dicho autor menciona que este cambio en la alimentación se debe básicamente a cambios en la morfología mandibular.
En síntesis, los resultados obtenidos sobre la ecología alimentaria del huachinango, del litoral del estado de Guerrero, permiten sugerir que existe un patrón diferencial en la utilización de los recursos presa (tipo, posiblemente tamaño y proporción), entre sus distintas clases de talla. Esto puede ser una estrategia ecológica para la optimización en el uso de recursos tróficos, para reducir o evitar el nivel de competencia intraespecífca (Schoener 1974, Werner 1979).
Agradecimientos
Queremos hacer patente nuestro agradecimiento a los pescadores artesanales de Punta Maldonado por facilitar la labor de muestreo. A Azucena Carballido por la identificación de crustáceos. AFelipe Galván y Francisco Gutiérrez por la identificación de restos de peces.
Resumen
En este artículo se describen los hábitos alimentarios del huachinango del Pacífico Lutjanus peru en la costa sur del estado de Guerrero, México. Estos datos son importantes para el manejo de las pesquerías locales. A partir de capturas comerciales, se recolectaron 385 estómagos llenos (206 de hembras y 179 de machos) en total, mensualmente, durante 1993 y 1994. El contenido estomacal se identificó hasta el nivel de especie cuando fue posible. Los elementos alimentarios se agruparon taxonómicamente, y se cuantificaron, se pesaron y se obtuvo la frecuencia de aparición. Se calculó el índice de importancia relativa (IIR) del total de la muestra, por meses, por sexos y tallas. Se obtuvieron 42 elementos alimentarios en total, en 1993, y 43 en 1994. En 1993 La dieta estuvo constituida por peces, crustáceos y moluscos. El agrupamiento de las especies, géneros o familias presa produjo 14 grupos alimentarios, de la siguiente forma: Moluscos, Anfípodos, Copépodos, Estomatópodos, Penaeus vannamei, Larvas de braquiuros, Larvas de estomatópodos, Larvas de la familia Albuneidae, Otros crustáceos, Anchoa ischana, Anchoa lucida, Otros peces, Salpas, Moni. Anchoa ischana dominó el espectro trófico ambos años. Pero, los Anfípodos, Anchoa lucida y Otros peces tuvieron índices importantes en 1994. Existió una gran variabilidad en la composición de la dieta de cada mes. Sin embargo, los crustáceos en conjunto dominaron la dieta de febrero a agosto de 1993. En 1994, los crustáceos sólo fueron dominantes en marzo y agosto. No se encontraron diferencias en la dieta de peces inmaduros, machos y hembras. La dieta del huachinango del Pacífico varía con el crecimiento, aunque siempre se encuentran los mismos grupos de alimento. En la fase adulta, en clases de longitud mayores a 261 mm LF, existe un cierto grado de especialización, ya que consumen más peces, principalmente engráulidos.
Referencias
Allen, G.R. 1985. Snappers of the world an annotated and illustrated catalogue of lutjanid species known to date. FAO Fisheries Synopsis 125(6): 1-208. [ Links ]
Barroso, L.M. 1965. Regime alimentar do pargo (Lutjanus aya Blonch, 1975) nordeste Brasileiro. Bol. Est. Pesca 5(3): 1-16. [ Links ]
Claro, R. 1981a. Ecología y ciclo de vida del pargo criollo, Lutjanus analis (Cuvier), en la plataforma Cubana. Inf. Cient. Téc. Biol. Pesq. Acad. Cienc. Cuba 186:1-83. [ Links ]
Claro, R. 1981b. Ecología y ciclo de vida de la biajaiba Lutjanus synagris (Linnaeus) en la plataforma Cubana. Inf. Cien. Tec. II, Biol. Pesq. Acad. Cienc. Cuba 177: 1-53. [ Links ]
Claro, R. 1983a. Ecología y ciclo de vida de la rabirubia Ocyurus chrysurus (Bloch) en la plataforma cubana. Identificación, distribución, hábitat, reproducción y alimentación. Inf. Cien. Tec. Acad. Cienc. Cuba 15:1-33. [ Links ]
Claro, R. 1983b. Ecología y ciclo de vida de la biajaiba, Lutjanus synagris, (Linnaeus), en la plataforma cubana. V. Dinámica estacional de algunos indicadores morfofisiológicos. Rep. Invest. Inst. Oceanol. Acad. Cienc. Cuba 16:1-24. [ Links ]
Claro, R. 1983c. Ecología y ciclo de vida del caballerote Lutjanus griseus (Linnaeus), en la plataforma Cubana. Identidad, distribución y hábitat, alimentación y reproducción. Rep. Invest. Oceanol. Acad. Cienc. Cuba 7:1-30. [ Links ]
Cruz-Romero, M. Espino-Bar, J. Mimbela-López, A. García- Boa, L. Obregón-Alcaraz & E. Girón-Botello. 1991. Biología reproductiva de tres especies de Lutjánidos en Colima. Reporte Técnico C.R.I.P. Manzanillo, Colima, INP México. 118 p. [ Links ]
Díaz-Uribe, J.G. 1994. Análisis trofodinámico del huachinango, Lutjanus peru, en las bahías de la Paz y la Ventana., B.C.S., México. Tesis de maestría en ciencias, CICESE, Ensenada, B.C., México. 57 p. [ Links ]
Elorduy-Garay, J.F. & J. Caraveo-Patiño. 1994. Hábitos alimentarios de la pierna, Caulolatilus princeps Jenyns 1842 (Pisces: Branchiostegidae), en la Bahía de La Paz, B.C.S., México. Ciencias Marinas 20(2): 199-218. [ Links ]
González-Sansón, G. & L. Rodríguez-Viñas. 1983. Alimentación natural de Eugerres brasilianus (Cuvier) y Gerres cinereus (Walbaum) (Pisces: Gerreidae) en las lagunas costeras de Tunas de Zaza, Cuba. Rev. Invest. Mar., Ser. 8, 4(1): 91-143. [ Links ]
Guevara, C.E., M.A. Bosch, M.R. Suárez & R.R. Lalana. 1994. Alimentación natural de tres especies de pargos (Pisces: Lutjanidae) en el Archipiélago de los Canarreos, Cuba. Rev. Invest. Mar. 15(1): 63-72. [ Links ]
Oda, K.D. & J.D. Parrish. 1981. Ecology of commercial snappers and groupers introduced to hawaiian reefs. Proceedings of the Fourth International Coral Reef Symposium. Manila, Vol. 1: 59-67. [ Links ]
Parrish, J.D. 1987. The trophic biology of snapper and grouper, pp. 561-603. In J.J. Polovina & S. Ralston (eds.). Tropical Snappers and Groupers: Biology and Fisheries Management. Westview, Boulder, Colorado. [ Links ]
Pedroso, B. & B.L. Anderes. 1983. Resultados preliminares de los hábitos alimentarios del caballerote (Lutjanus griseus Linnaeus, 1758) en pesqueros artificiales. Rev. Cub. Inv. Pesq. 8(2): 75-84. [ Links ]
Pinkas, L., M.S. Oliphant & I.L. Iverson. 1971. Food habits of albacores, bluefin tuna and bonito in California water. Calif. Dept. Fish and Game Fish Bull. 152:1-105. [ Links ]
Polovina, J.J. & S. Ralston (eds.). 1987. Tropical Snappers and Groupers: Biology and Fisheries Management. Westview, Boulder, Colorado. 603 p. [ Links ]
Randall, J.E. 1967. Food habits of reef fishes of the West Indies. Stud, Trop. Oceanogr. Miami 5: 665-847. [ Links ]
Rocha-Olivares, A. 1991. Edad y crecimiento del huachinango del Pacífico Lutjanus peru (Nichols y Murphy, 1922) (Perciformes: Lutjanidae) en la Bahía de La Paz y zonas adyacentes, B.C.S., México. Tesis de licenciatura, U.A.B.C.S., La Paz, B.C.S., 121 p. [ Links ]
Rojas-Herrera, A.A. 1990. Análisis de las capturas en las pesquerías del estado de Guerrero, México. Mem. VII Congreso Nacional de Oceanografía, Mazatlán, Sin. Mex. [ Links ]
Rojas-Herrera, A.A. 1996. Análisis de la alimentación del flamenco Lutjanus guttatus (Steindachner, 1869) de la costa de Guerrero, México. Mem. primer encuentro regional sobre investigación y desarrollo costero: Guerrero, Oaxaca y Chiapas. Puerto Ángel, Oaxaca. Mex. [ Links ] [ Links ]
Rojas, M.J.R. 1997b. Dieta del "pargo colorado" Lutjanus colorado (Pisces: Lutjanidae) en el Golfo de Nicoya, Costa Rica. Rev. Biol. Trop. 45(3): 1173-1183. [ Links ]
Rooker, J.R. 1995. Feeding ecology of the schoomaster snapper Lutjanus apodus (Walbaum), from South-western Puerto Rico. Bull. Mar. Sci. 56(3): 881-894. [ Links ]
Santamaría-Miranda, A. & A.A. Rojas-Herrera. 1994a Análisis de la variación morfofisiológica del huachinango, Lutjanus peru (Nichols y Murphy, 1922) de la costa chica del estado de Guerrero, México., periodo 1993-1994. Mem. IV Congreso Nacional de Ictiología, Morelia Mich. 1994: 104. [ Links ]
Santamaría-Miranda, A. & A.A. Rojas-Herrera. 1994b. Análisis de la variación morfofisiológica del flamenco, Lutjanus guttatus (Steindachner, 1869) (Pisces:Lutjanidae), en el litoral de Acapulco, Guerrero, México, periodo 1993-1994. Mem. XII Congreso Nacional de Zoología, Morelia, Mich. 1995: 183. [ Links ]
Schoener, T.W. 1974. Resource partitioning in ecological communities. Science, 185: 27-39. [ Links ]
Sierra, L.M. & O.A. Popova. 1989. Dinámica del ritmo de la alimentación de varias especies de peces neríticos del Golfo de Batabanó en diferentes años. Rep. Invest. Inst. Oceanol. Acad. Cienc. Cuba (2): 1-19. [ Links ]
Sierra, L.M., R. Claro & O.A. Popova. 1994. Alimentación y relaciones tróficas. En: R. Claro (ed.). Ecología de los peces marinos de Cuba. Instituto de Oceanología Academia de Ciencias de Cuba y Centro de Investigaciones de Quintana Roo (CIQRO), México. 525 p. [ Links ]
Sierra, L.M. & O.A. Popova. 1997. Relaciones tróficas de los juveniles de cinco especies de pargo (Pisces: Lutjanidae) en Cuba. Rev. Biol. Trop. 44(3)/45(1): 499-506. [ Links ]
Sokal, R.R. & F.J. Rohlf. 1979. Biometría. Principios y métodos estadísticos en la investigación biológica. H. Blume, España. 832 p. [ Links ]
Stark, W.A. 1971. Biology of gray snapper, Lutjanus griseus (Linnaeus) in the Florida Keys. Stud. Oceanog. 10: 11-150. [ Links ]
Suárez, A. & F.F. Rubio. 1983. Los hábitos alimentarios de juveniles de Lutjanus guttatus (Pisces: Lutjanidae) en la bahía de Málaga, Pacífico de Colombia. Res.Mem. Congr. Centroamericano y del Caribe en Ciencias del Mar. Universidad del Valle, Colombia 1: 1-152. [ Links ]
Werner, E.E. 1979. Niche partitioning by food size in fish communities, pp. 311-322. In R.H. Stroud & H. Clepper (eds.). Predator prey systems in fisheries management. Sport Fishing Inst., Washington. [ Links ]
1 CIIDIR Sinaloa, Carretera a Las Glorias, km. 1, Apdo. postal 280, Guasave, Sin., Méx. 81100. Tel.: + 687 2 96 26. Fax: + 2 96 25. Becario EDI-IPN; asantama@redipn.ipn.mx .
2 Depto. Pesquerías y Biología Marina, CICIMAR - IPN, Ave. IPN s/n, col. Playa Palo de Sta. Rita, 23090 La Paz, BCS, Méx. Becario COFAA-IPN, EDI-IPN; jelorduy@redipn.ipn.mx
3 Escuela Superior de Ecología Marina Gran Vía Tropical 20, Fracc. Las Playas, Acapulco, Gro., Méx. Tel.: +74 83 27 80

