










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.50 n.3-4 San José Dec. 2002
Abstract
Donax dentifer is a dominant member of the sandy beach communities of the Colombian Pacific coast. A population of this bivalve was monthly sampled in Playa Chucheros at the Bay of Málaga, Colombian Pacific, between August 1997 and July 1998, during the "El Niño 1997/1998" phenomenon. Quantitative samples were taken in perpendicular and parallel to the coastal line transects to determine the population density and a length-frequency distribution. Qualitative samples were taken to study the reproduction and monthly length weight relationship. Sea surface temperature, dissolved oxygen, salinity and pH in the water was monthly registered in the sampling station. Additionally, a tagging-recapture experiment was performed to growth estimations. Results indicate a continuous reproduction cycle with two annual spawning seasons in December and March. The growth parameters of the non-oscillating von Bertalanffy growth funetion were K= 0.62 yr -1 and L![]() = 29.3 mm. Mortality (Z) was higher (2.65 yr-1) for small individuals (between 2 mm and 5 mm) while a lower Z value (1.71 yr-1 ) were obtained for larger ones (between 19 mm and 25 mm). An annual mean biomass value of B = 1.229 g ash-free dry weight m-2 and an annual somatic production of Psom =1.215 g ash-ftee dry weight m -2 were estimated, which correspond to a Psom / B value of 0.988. Population density increased in November and April, but any population density pattem related with "El Niño" was evident. Low body weight during NovemberDecember seems to show a relation between reproductive cycle in D. dentifer and El Niño 97/98 event. The fluctuation in dissolved oxygen show a high correlation with parameters of the reproductive cycle, which could be relatad to assimilation process and body weight cycle. Growth in D. dentifer in the Colombian Pacific coast was faster than other Donax species, as can be expected for tropical bivalves mollusks in comparison whit high latitude bivalves. The mortality pattern suggest that the population was selectively affected by El Niño 97/98, due to smaller individuals are living in the lowest areas of the beach, where they are exposed to abnormal seawater conditions for a longer time. Individual and population production was higher for medium-size animals than each for small-size ones, which were the most abundant in the population.
= 29.3 mm. Mortality (Z) was higher (2.65 yr-1) for small individuals (between 2 mm and 5 mm) while a lower Z value (1.71 yr-1 ) were obtained for larger ones (between 19 mm and 25 mm). An annual mean biomass value of B = 1.229 g ash-free dry weight m-2 and an annual somatic production of Psom =1.215 g ash-ftee dry weight m -2 were estimated, which correspond to a Psom / B value of 0.988. Population density increased in November and April, but any population density pattem related with "El Niño" was evident. Low body weight during NovemberDecember seems to show a relation between reproductive cycle in D. dentifer and El Niño 97/98 event. The fluctuation in dissolved oxygen show a high correlation with parameters of the reproductive cycle, which could be relatad to assimilation process and body weight cycle. Growth in D. dentifer in the Colombian Pacific coast was faster than other Donax species, as can be expected for tropical bivalves mollusks in comparison whit high latitude bivalves. The mortality pattern suggest that the population was selectively affected by El Niño 97/98, due to smaller individuals are living in the lowest areas of the beach, where they are exposed to abnormal seawater conditions for a longer time. Individual and population production was higher for medium-size animals than each for small-size ones, which were the most abundant in the population.
Key words
Donax dentifer, population dynamics, El Niño, reproduction, growth, mortality, production, Colombia.
La zona costera del Pacífico colombiano, incluida dentro de la franja conocida como la Ensenada de Panamá, se localiza en una región de bajas presiones atmosféricas donde convergen los vientos Alisios en el Cinturón de Convergencia Intertropical. Debido a esto, el clima se caracteriza por vientos variables y débiles y una de las tasas de pluviosidad más altas del mundo. Por su localización en una zona de colisión de placas litosféricas la franja costera presenta una topografía muy accidentada y un sistema muy activo de bandas sedimentarlas costeras (Cantera 1993 ). Uno de los ecosistemas típicos de esta región son las playas arenosas, que se encuentran generalmente en las costas abiertas o en las aberturas de los estuarios, bahías, golfos y desembocaduras de los ríos. En estas playas arenosas son comunes y dominantes las poblaciones de "almejas", bivalvos del género Donax. Al igual que una gran parte del Pacífico Oriental Suramericano, esta zona es afectada por El Niño (EN), un evento climático global durante el cual aparecen aguas superficiales más cálidas de lo normal y un incremento del nivel medio del mar. Estas condiciones repercuten en la distribución de especies planctónicas y bentónicas ( Anónimo 1998 ), sin embargo es muy poco lo que se sabe de los efectos de este evento a nivel de la estructura de las poblaciones marinas. El objetivo de este estudio fue determinar los aspectos básicos de la reproducción, crecimiento, mortalidad y producción de una especie dominante en ecosistemas de playa arenosa, como es Donax dentifer (Hanley 1843), bajo las particulares condiciones del más reciente evento EN (1997/1998).
Materiales y métodos
La investigación se realizó en la Bahía de Málaga, Valle del Cauca, Colombia (4º 05' N y 77º 16'W), una bahía de 18 km de largo por 7 km de ancho, de profundidad variable entre 12 y 15 m que se caracteriza por la presencia de varios ríos y quebradas que desembocan en ella ( Fig. 1 ). La bahía está constituida por sedimentos consolidados del Terciario, donde son abundantes los acantilados, playas rocosas, depósitos de fango y playas arenosas. La salinidad máxima en el área llega a 27%, variando entre 18% y 23%; la temperatura superficial del agua permanece aproximadamente constante a lo largo del año, con oscilaciones entre 26º y 29 ºC (Rubio 1984 ).
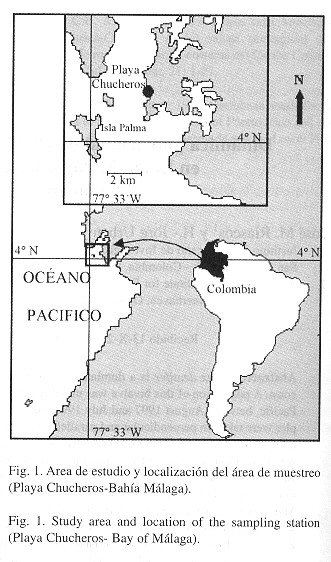
Desde julio de 1997 hasta julio de 1998 se tomaron manualmente muestras cualitativas mensuales (300 a 500 individuos) en la zona de rompiente en la playa, donde los animales quedan expuestos durante la marca alta. Para cada individuo se registró la longitud del eje anteroposterior de la concha (L) usando un calibrador digital. De estos se seleccionaron 70 animales que cubrieron todo el rango de tallas de la población (entre 5 y 32 mm) y se llevaron al laboratorio para posteriores análisis; los demás se liberaron. Adicionalmente se tomaron muestras cuantitativas mensuales en dos transectos, uno paralelo u horizontal a la línea de costa en la zona mesolitoral y otro perpendicular o vertical. En cada transecto se ubicaron 12 cuadrantes de 0.25 m2 , en los cuales se removió la arena hasta una profundidad de 20 cm y luego se pasó por un tamiz con ojo de malla de 0.5 mm, registrando la longitud y el número de individuos retenidos. Trimestralmente 300 a 400 individuos procedentes de las muestras cualitativas se midieron y marcaron usando un código adherido a la concha, liberándolos posteriormente. Especímenes testigo se incluyeron en la Colección Zoológica de Referencia de INCIVA - Museo de Ciencias Naturales "FCLV" (Cat. No. = IMCN-0222 a IMCN-00233). A partir de agosto se registró mensualmente la longitud L de los individuos recapturados, obteniendo así pares marca-recaptura. Simultáneamente con los muestreos se registraron las fluctuaciones de algunas variables ambientales de la columna de agua que inunda la zona durante marea alta: temperatura superficial (ºC), pH, oxígeno disuelto (mg/l) y salinidad (%).
En el laboratorio se tomó una primera submuestra de 30-35 individuos cubriendo el rango de tallas para estudiar los eventos reproductivos de la población. Para ello se calculó el peso seco corporal libre de ceniza "Ash-Free Dry Weight" (AFDW) y se determinó el Indice Condicional (IC), definido como la estimación mensual del peso corporal respecto a la longitud de la concha (L) (Urban 1996, Urban y Tarazona 1996). Esta determinación se basa en el cálculo del valor de las constantes a y b de la ecuación:
Donde W es el peso (g) AFDW y L es la longitud de la concha. Con base en regresiones lineales, después de una transformación logarítmica, se calculó el peso de un individuo estándar de 30 mm de longitud.
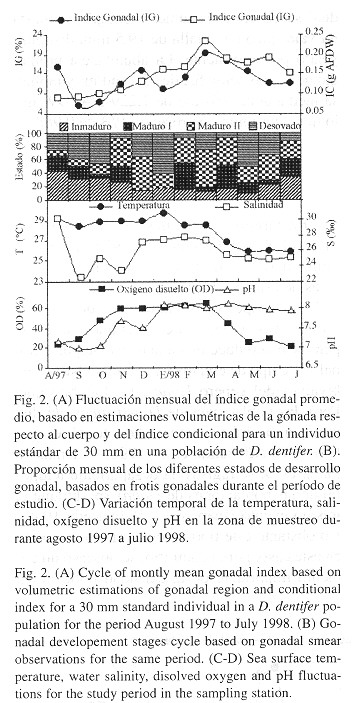
Adicionalmente se estimó el Indice Gonadal (IG) para cada individuo en la segunda submuestra de 35 animales. Este índice consiste en una cuantificación del peso o volumen relativo del tejido reproductivo respecto al peso o volumen del cuerpo. Esta medida relativa es usada para discernir, con base en cambios temporales de las gónadas, los períodos de actividad reproductiva debido a su relación con procesos de gametogénesis ( Cruz y Villalobos 1993 ). La estimación se realizó según la siguiente ecuación:
Donde Vg es una cuantificación volumétrica indirecta de la región gonadal y Vc el volumen total del cuerpo estimado por desplazamiento de agua. Para cuantificar el volumen gonadal se asumió, con base en la observación de cortes de la región gonadal, que la forma corresponde a un elipsoide, tomando las medidas necesarias para estimar el volumen de este cuerpo geométrico (Urban y Riascos 2002 ).
Usando los mismos animales se tomó una muestra del tejido gonadal, haciendo frotis que se observaron microscópicamente para determinar el sexo y Estado de Madurez de los gametos. Para esto se adoptó un método similar al usado por Guillou et al. (1990) , en el cual tanto hembras como machos pueden clasificarse en 4 estadios: Inmaduros, Maduro I, Maduro II y Desovado. Para evaluar la influencia de los factores ambientales sobre la reproducción, se realizó un análisis de correlación de Spearman entre las variables ambientales, los índices (IC, IG) y la proporción de estadios Maduro I y II.
El análisis del crecimiento individual se realizó mediante el modelo de crecimiento no estacionalizado de von Bertalanffy (1938) :
Donde Lt es la longitud L en la edad t [años]; L
Donde, L1 es la longitud en el momento del marcaje y L 2 es la longitud en el momento de recaptura y t2 - t1 es el tiempo transcurrido entre el marcaje y la recaptura. Nótese que con este modelo el parámetro t o no puede estimarse. Sin embargo este puede asumirse como cero utilizando edades relativas [años-t0 ] . Es conocido que los métodos basados en datos de incrementos de tallas, como el método de Fabens, pueden llevar a hacer determinaciones erróneas del parámetro L
Como método de comparación de los parámetros estimados con otros reportados en la literatura y con base en la relación estadística que existe entre K y L![]() , planteada por Pauly (1979) se calculó el parámetro
, planteada por Pauly (1979) se calculó el parámetro ![]() a partir de la ecuación:
a partir de la ecuación:
La mortalidad total (Z) de la población se calculó a partir de los datos de longitud-frecuencia acumulados, los parámetros de crecimiento y la curva de captura linealizada convertida a tallas (Pauly 1983 ). Previamente los datos de longitud se convirtieron a edades usando la formula inversa de von Bertalanffy:
La mortalidad Z se calculó entonces mediante un análisis de regresión:
Donde a y b son las constantes de la recta, es el número de individuos en el intervalo de longitud i; ![]() ti es el tiempo requerido para crecer de un intervalo de longitud a otro y ti es la edad en el intervalo de longitud i.
ti es el tiempo requerido para crecer de un intervalo de longitud a otro y ti es la edad en el intervalo de longitud i.
La producción somática (Psom) e individual (Pind) de la población se calculó con base en el método de Crisp (1984) como una tasa de incremento específico en peso, a partir de la densidad media mensual de las muestras cuantitativas, los datos de longitud-frecuencia, los parámetros K y L![]() y la relación de longitud-peso (AFDW) durante el año:
y la relación de longitud-peso (AFDW) durante el año:
Donde Ni es el número promedio de individuos (N m-2) del intervalo de longitud i, W i es el peso corporal promedio del intervalo i (g AFDW) y Gi es la tasa de incremento de peso (años-1):
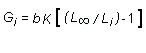
La biomasa media anual (B) se estimó con base en la densidad y la relación anual entre longitud y peso:
Resultados
Reproducción: De un total de 391 individuos analizados, el 55.5% fueron machos, 33% hembras y el 11.5% se catalogaron como inmaduros. En la Fig. 3A se observa la variación del Indice Condicional e Indice Gonadal. Partiendo de un valor mínimo en agosto/97 (0.091g AFDW) el peso corporal aumenta progresivamente hasta marzo/98 (0.233g AFDW). Después de ese punto el peso vuelve a disminuir hasta julio. El índice gonadal presenta un patrón similar, pero con un pico también en diciembre, lo que indica un patrón bianual de desove. La evaluación del estado de madurez gonadal (Fig. 3b ) muestra que los estados Maduro I y II están presentes todos los meses, es decir que la población está reproductivamente activa todo el año, pero prevalecen en noviembre y febrero-abril, indicando nuevamente un patrón bianual.
Los parámetros ambientales registrados (Fig. 2C y 2D ) indican que la temperatura fue alta, llegando a 30.3ºC en enero, posteriormente decreció en forma gradual hasta 26ºC en mayo. La salinidad alcanzó su valor máximo en agosto (30.5% o ) el cual es un valor inusualmente alto en la zona. El oxígeno disuelto y el pH mostraron valores bajos al inicio del estudio y aumentaron entre febrero y abril. El análisis de correlación de Spearman ( Cuadro 1 ) mostró un grado de asociación alto entre el oxígeno disuelto y el índice condicional y en menor medida la salinidad y el índice gonadal.
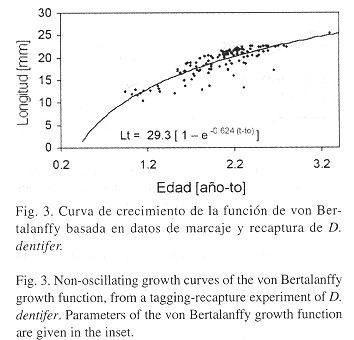
Crecimiento: Un total de 1380 individuos fueron marcados, de los cuales se recapturaron 146 (10.5%). La estimación de los parámetros de crecimiento se muestra en el Cuadro 2 . Puede observarse que hay una diferencia sustancial entre el valor de L max (30.5 mm) y L
La Fig. 3 muestra la curva de crecimiento ajustada con los valores definitivos de K y L![]() . Para graficar los puntos, la edad de L 1 (datos de marcaje y recaptura) se estimó con la fórmula inversa de von Bertalanffy.
. Para graficar los puntos, la edad de L 1 (datos de marcaje y recaptura) se estimó con la fórmula inversa de von Bertalanffy.
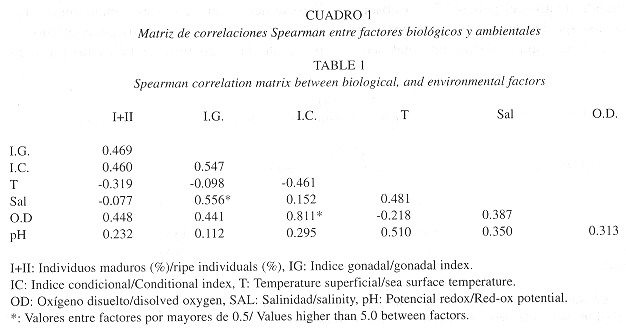
Mortalidad: Debido a que la población estudiada no está sometida a presión por pesca, se asumió que la mortalidad (Z) es igual a la mortalidad natural (M) de la población. La Fig. 4 muestra dos tendencias en la mortalidad de la población. Para los individuos más pequeños de la población, entre los 0.5 y 16.5 mm la mortalidad es más alta (2.605) y para los animales más grandes de 21.5 mm, la mortalidad es menor (1.717).
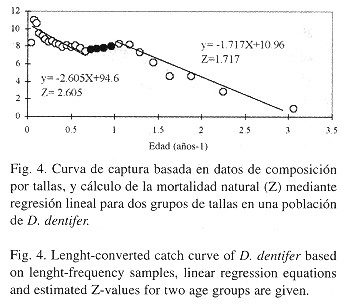
Producción: La producción somática anual de la población alcanzó un valor de 1.2 g AFDW/m2, en el que la mayor parte es sustentada por los individuos de tamaño moderadamente grande (19-23 mm) ( Fig. 5 ). Obsérvese que aunque estos individuos no son los más abundantes en la población, son los que más aportan en la producción somática total. Por otro lado, aunque los individuos pequeños son definitivamente los más abundantes, es muy poco lo que aportan en peso a la producción poblacional. En cuanto a la producción individual se observa un aumento progresivo hasta un valor pico en la talla de 19.5 mm, disminuyendo posteriormente. La abundancia promedio de la población fue de 65.1 ind/m2 y biomasa media anual (B) fue de 1.229 g AFDW/ m2, lo que determinó una relación P/B de 0.988.

Discusión
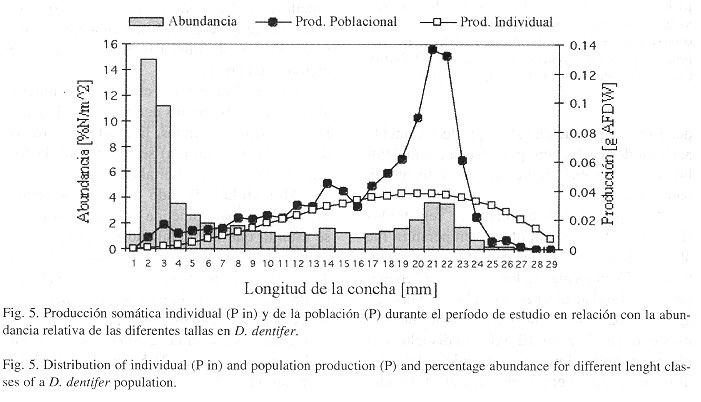
En términos generales los efectos de un evento EN fuerte (como el de 97/98) sobre la fauna marina se traducen en mortalidades masivas de especies bentónicas y emigración forzada de especies pelágicas y demersales (Arntz et al. 1988 ). En este sentido podría esperarse una reducción en la densidad poblacional. Este tipo de efectos ya se han registrado en bivalvos del genero Donax. En Lambayeque, Perú, las poblaciones de D. marincovichi disminuyeron drásticamente durante el evento EN 97/98 ( Chaname et al. 1998 ). Además en Paita, Perú, se registró la desaparición temporal de las poblaciones de Donax sp. durante el mismo evento ( Inapaqué y Balmaceda 1998 ). No obstante este tipo de efecto no se presentó en este caso, por el contrario, se observó un aumento importante de la densidad poblacional durante noviembre y abril (Fig. 6 ), a causa del proceso de reclutamiento de la población ( Riascos y Urban 1999 ). Por otro lado no se encontró una relación evidente entre los cambios de temperatura y la densidad poblacional. Un tipo de comportamiento similar en cuanto a densidad poblacional encontraron Arntz et al. (1987) para de D. peruvianus en Santa María del Mar, Perú, (12 'S) durante el evento EN 1982/83. En este caso la densidad poblacional permaneció casi constante y unos pocos meses después del evento se registró un incremento importante.
Aunque la población presenta una actividad reproductiva contínua durante el año, tanto la proporción de estados Maduro I y Maduro II como el Indice Gonadal indican dos picos anuales de actividad reproductiva (noviembre-diciembre y marzo-abril). Sin embargo el Indice Condicional sugiere un patrón unimodal con un pico en marzo. Este hecho podría estar relacionado con un efecto de EN 97/98 sobre el peso corporal. Se ha observado que bajo este tipo de condiciones de estrés, la energía disponible se desvía hacia el metabolismo y por lo tanto disminuye la energía disponible para el crecimiento y la reproducción ( Urban y Tarazona 1996 ). Consecuentemente, un patrón bimodal en el ciclo del IC estaría siendo enmascarado como consecuencia de EN 97/98. Efectos similares al propuesto fueron observados por Birket y Cook (1987) sobre los patrones de desove en D. serra en la costa de Sur Africa durante el evento El Niño 82/83. Las condiciones oceanográficas preponderantes en el área de estudio son fuertemente afectadas por la pluviosidad y el aporte hídrico de los ríos que desembocan en la bahía. En condiciones normales la zona de estudio presenta dos picos lluviosos, uno entre abril y mayo y otro más importante entre septiembre y noviembre (Montealegre 1988 , Cantera 1991 ). El evento EN 97/98 en el área de estudio se caracterizó por la reducción del régimen normal de lluvias y, por consiguiente, por la disminución del caudal normal de los ríos que desembocan en la zona (Anónimo 1998 ) De esta manera, durante la etapa crítica del evento, el pico lluvioso de noviembre-diciembre se vió drásticainente disminuido. Esta situación se reflejó en el comportamiento de las variables ambientales evaluadas (temperatura, salinidad, oxígeno disuelto y pH) que en general muestran un aumento gradual desde agosto/97 hasta enero-febrero/98 y un descenso posterior (Fig. 2C, 2D ) que coincide con el inicio del debilitamiento del fenómeno. Aunque se ha propuesto a la temperatura como uno de los factores claves para el crecimiento y los procesos reproductivos (Vakily 1992 ) no se observó una relación evidente en este caso ( Cuadro 1 ). El análisis de correlación de Spearman mostró una fuerte correlación (R=0. 811) entre el oxígeno disuelto y el Indice Condicional, lo que resalta la relación que existe entre el oxígeno y el proceso de asmilación y por lo tanto en el aumento de peso. Por otro lado la correlación (R=0.556) entre la salinidad y el índice condicional probablemente esté reflejando la sincronía entre la época lluviosa (disminución de salinidad) y los eventos de desove.
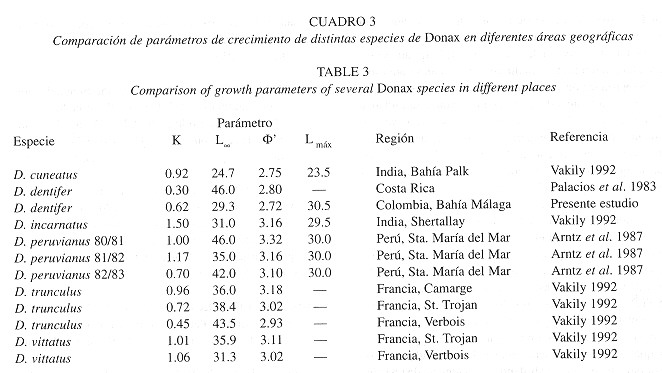
Para analizar el crecimiento de D. dentifer, en el Cuadro 3 se muestran estimaciones de K, L![]() ,
, ![]() ' y Lmax de diferentes especies del género reportadas en la literatura. Con base en esto se puede afirmar que el crecimiento de D. dentifer en el Pacífico colombiano es lento (K=0.624), tratándose de un animal de talla media/grande dentro del género. Se ha observado que las especies de distribución tropical o subtropical presentan un crecimiento más acelerado que sus contrapartes de altas latitudes (Urban y Tesch 1996 ), por lo cual se esperaría un valor más alto del parámetro K. Por otra parte se puede observar que el valor de K es muy similar al encontrado por Arntz et al. (1987) para D. peruvianus durante 1982/1983, justamente bajo la influencia de EN 82/83, catalogado como uno de los mas fuertes del presente siglo. Por estas razones es posible sugerir que los valores de los parámetros estimados en este estudio reflejan un efecto negativo sobre el crecimiento de esta especie. Por otro lado el valor de
' y Lmax de diferentes especies del género reportadas en la literatura. Con base en esto se puede afirmar que el crecimiento de D. dentifer en el Pacífico colombiano es lento (K=0.624), tratándose de un animal de talla media/grande dentro del género. Se ha observado que las especies de distribución tropical o subtropical presentan un crecimiento más acelerado que sus contrapartes de altas latitudes (Urban y Tesch 1996 ), por lo cual se esperaría un valor más alto del parámetro K. Por otra parte se puede observar que el valor de K es muy similar al encontrado por Arntz et al. (1987) para D. peruvianus durante 1982/1983, justamente bajo la influencia de EN 82/83, catalogado como uno de los mas fuertes del presente siglo. Por estas razones es posible sugerir que los valores de los parámetros estimados en este estudio reflejan un efecto negativo sobre el crecimiento de esta especie. Por otro lado el valor de ![]() (medida de la relación entre L
(medida de la relación entre L![]() y K) para este estudio, es el más bajo de los reportados en la literatura, lo que indica que la estimación es atípica y estaría confirmando la suposición de un efecto de las condiciones ambientales sobre el crecimiento. Es probable que el crecimiento de esta especie en condiciones normales sea más alto que el estimado en este estudio. Hay que señalar que el valor de K estimado por Palacios et al. (1983) difiere de nuestros resultados pese a tratarse de la misma especie. Creemos que el bajo valor de
y K) para este estudio, es el más bajo de los reportados en la literatura, lo que indica que la estimación es atípica y estaría confirmando la suposición de un efecto de las condiciones ambientales sobre el crecimiento. Es probable que el crecimiento de esta especie en condiciones normales sea más alto que el estimado en este estudio. Hay que señalar que el valor de K estimado por Palacios et al. (1983) difiere de nuestros resultados pese a tratarse de la misma especie. Creemos que el bajo valor de ![]() ' (2.80) indica que la estimación no es confiable, ya que en este caso los parámetros de crecimiento se calcularon a partir de datos de frecuencias de longitudes. Sparre y Venema (1995) observaron que los métodos para estimar el crecimiento, basados únicamente en datos de longitud frecuencia, no son siempre confiables en el caso de especies tropicales. Esto se debe a que en estas poblaciones la reproducción y el reclutamiento son continuos, y por tanto la estructura de tallas no presenta modas definidas, susceptibles de ser analizadas como cohortes separadas.
' (2.80) indica que la estimación no es confiable, ya que en este caso los parámetros de crecimiento se calcularon a partir de datos de frecuencias de longitudes. Sparre y Venema (1995) observaron que los métodos para estimar el crecimiento, basados únicamente en datos de longitud frecuencia, no son siempre confiables en el caso de especies tropicales. Esto se debe a que en estas poblaciones la reproducción y el reclutamiento son continuos, y por tanto la estructura de tallas no presenta modas definidas, susceptibles de ser analizadas como cohortes separadas.
Al observar la abundancia relativa de las distintas tallas en la población, es claro que existen dos grupos de tallas que dominan numéricamente la población, uno que va de 2 a 5 mm y otro de 19 a 23 mm aproximadamente ( Fig. 5 ). Con base en esta observación se realizaron estimativos de mortalidad para cada grupo dominante mostrando la existencia de una mortalidad alta en los individuos pequeños, y menor en los individuos grandes ( Fig. 4 ). Las condiciones ambientales extremas ocasionadas por el evento EN 97/98 probablemente influyen en el patrón de mortalidad encontrado ya que los individuos más pequeños son más vulnerables. Esto es debido a que la población muestra un patrón de zonación por tallas en el que los individuos pequeños se encuentran en las partes más bajas de la playa y están sometidas durante un tiempo más largo a las condiciones extremas del agua marina ( Riascos y Urban 1999 ).
Pese a que los individuos de tallas medias (13 a 24 mm) no dominan numéricamente en la población, la producción poblacional e individual es mayor que la de los individuos pequeños, numéricamente dominantes (Fig. 5 ).
Infortunadamente no existe información de referencia, que permita dilucidar posibles efectos ambientales sobre la población. Solamente a partir de estudios posteriores será posible hacer determinaciones definitivas sobre la manera como el evento EN 97/98 afectó esta especie.
Agradecimientos
Este trabajo forma parte del plan institucional de investigaciones auspiciado por el Instituto Vallecaucano de Investigaciones Científicas, INCIVA. Los autores agradecen a Germán Parra Valencia por su permanente apoyo para la realización de este proyecto y a Norma Ibón Rallón por su apoyo en las actividades de campo y laboratorio.
Resumen
Donax dentifer es un bivalvo dominante en comunidades de playa arenosa en la costa del Pacífico colombiano. Se muestreó mensualmente una población de este bivalvo en Bahía Málaga, Pacífico colombiano entre agosto de 1997 y julio de 1998 cuando ocurrió el evento "El Niño" 1997/1998. Se tomaron muestras cuantitativas mensuales usando transectos lineales perpendiculares y paralelos a la línea de costa para determinar la densidad poblacional y realizar una distribución de frecuencias de longitud. Se tomaron también muestras cualitativas mensuales para estudiar la reproducción y establecer relaciones peso-longitud. Se registró la temperatura, oxígeno disuelto, salinidad y pH de la columna de agua en el sitio de muestreo. Adicionalmente se realizó un experimento de marcaje-recaptura para estimar el crecimiento de la población. Los resultados indican que la población tiene un ciclo de reproducción continuo, con dos períodos de desove anuales en diciembre y marzo. Los parámetros de crecimiento de la función de crecimiento de von Bertalanffy fueron K=0.624 año-1 y L![]() =29.3 mm. La mortalidad total (Z) fue de Z=2.605 año -1 para los individuos pequeños (entre 2 mm y 5 mm) y menor (Z=1.714 año -1) para los individuos grandes (entre 19 mm y 25 mm). La biomasa promedio anual de la población fue =1.22 g de peso seco libre de cenizas m-2 y la producción somática anual Psom/= 1.215 g de peso seco libre de cenizas m -2, lo que corresponde a una tasa de Psom/= 0.988. Aunque se observó un incremento en la densidad poblacional durante noviembre y abril, no hay evidencia de una clara relación entre la densidad poblacional y la ocurrencia del fenómeno "El Niño". Los cambios en la concentración de oxígeno disuelto mostraron una alta correlación con los parámetros del ciclo reproductivo, posiblemente como consecuencia de la influencia de este parámetro en procesos de asimilación y el ciclo de peso corporal. El crecimiento de D. dentifer fue más lento que en otras especies del género, como cabe esperarse para bivalvos tropicales en comparación con los de altas latitudes. El patrón de mortalidad observado sugiere un efecto selectivo de El Niño 97/98 sobre la población, debido a que los individuos pequeños ocupan las partes bajas de la zona intermareal, donde el grado de exposición al agua marina es alto. La producción poblacional e individual fue mayor para los animales de tallas medias en comparación con los animales pequeños aunque estos últimos fueron los más abundantes en la población.
=29.3 mm. La mortalidad total (Z) fue de Z=2.605 año -1 para los individuos pequeños (entre 2 mm y 5 mm) y menor (Z=1.714 año -1) para los individuos grandes (entre 19 mm y 25 mm). La biomasa promedio anual de la población fue =1.22 g de peso seco libre de cenizas m-2 y la producción somática anual Psom/= 1.215 g de peso seco libre de cenizas m -2, lo que corresponde a una tasa de Psom/= 0.988. Aunque se observó un incremento en la densidad poblacional durante noviembre y abril, no hay evidencia de una clara relación entre la densidad poblacional y la ocurrencia del fenómeno "El Niño". Los cambios en la concentración de oxígeno disuelto mostraron una alta correlación con los parámetros del ciclo reproductivo, posiblemente como consecuencia de la influencia de este parámetro en procesos de asimilación y el ciclo de peso corporal. El crecimiento de D. dentifer fue más lento que en otras especies del género, como cabe esperarse para bivalvos tropicales en comparación con los de altas latitudes. El patrón de mortalidad observado sugiere un efecto selectivo de El Niño 97/98 sobre la población, debido a que los individuos pequeños ocupan las partes bajas de la zona intermareal, donde el grado de exposición al agua marina es alto. La producción poblacional e individual fue mayor para los animales de tallas medias en comparación con los animales pequeños aunque estos últimos fueron los más abundantes en la población.
Referencias
Anónimo, 1998. El Niño y su efecto en el clima de Colombia. Innovacción y Ciencia VII(I): 56-62. [ Links ]
Arntz, W.E., E. Valdivia & J. Zeballos. 1988. Input of El Niño 1982-83 on the comercially exploited invertebrates of the Peruvian shore. Meeresforsch. 32: 3-22. [ Links ]
Arntz, W.E., T. Brey, J. Tarazona, & J. Robles. 1987. Changes in the structure of Shallow sandy beach community in Peru during an El Niño event. In A.I.L. Payne, J.A. Gulland & H.K. Brink. (eds.). The Benguela and Comparable Ecosystems. S. Afr. J. Mar. Sci. 5: 645-658. [ Links ]
Bertalanffy, L. von. 1938. A quantitative theory of organic growth (inquiries on growth laws II) Hum. Biol. 10(2): 181-213. [ Links ]
Birket, D.A. & P. Cook. 1987. Effect of the Benguela temperature anomaly, 1982-1983 on the breeding cycle of Donax serra Röding. In A.I.L. Payne, J.A. Gulland & H.K. Brink. (eds.). The Benguela and Comparable Ecosystems. S. Afr. J. Mar. Sci. 5: 191-196. [ Links ]
Cantera, J.R. 1991. Etude structurale des mangroves et des peuplements litoraux, des deux babies do Pacifique columbien (Málaga et Buenaventura). Rapport avec les conditions du melieu et les perturbations antropiques. These d' Etat Sciences. Université d'Aix-Marseille III. Marseille France, p. 429. [ Links ]
Cantera, J.R. 1993. Oceanografía. pp. 11-23. In R Leyva (ed.). Colombia: Pacífico. Tomo I Fondo para la protección del medio ambiente FEN. Santafé de Bogotá [ Links ].
Chaname, J., E. Tejada, V. Alvitres & J. Fupuy. 1998. Estructura del macrozoobentos de la playa arenosa de la caleta San José (Lamayeque - Perú) junio 1995julio 1998. Memorias IV Congreso Latinoamericano de Ecología y 11 Congreso Peruano de Ecología. Universidad San Agustín. Arequipa-Perú. pp. 7-12. [ Links ]
Crisp, D.J. 1984 Energy Flow measurements. pp. 284-372. In N.A. Holme & A.D. McIntyre (eds.). Methods for the study of marine benthos. Blackwell, Londres. [ Links ]
Cruz, R.A. & C.R. Villalobos. 1993. Shell length at sexual maturity and spawning cycle of Mytella guyanensis (Bivalvia: Mytilidae) from Costa Rica. Rev. Biol. Trop. 41(1): 89-92. [ Links ]
Fabens, A.J. 1965. Properties and fitting of the von Bertalanffy growth curve. Growth 29: 265-285. [ Links ]
Guillou, J., G. Bachelet, M. Desprez, J.M. Ducrotoy, I. Madani, H. Rybarczyk, P.G. Sauriau, B. Sylvand, B. Elkaim, & M. Glemarec. 1990. Les modalités de la reproduction de la coque (Cerastodenna edule) sur le littoral francais de la Manche et de l'Atlantique. Aquat. Living. Resour. 3: 29-41. [ Links ]
Inapaqué, T.J. & J. Balmaceda. 1998 Impacto biológico del evento "El Niño" sobre el macrozoobentos del intermareal arenoso "La Esmeralda" Colán - Paita. Agosto 1997 - Agosto 1998. Memorias IV Congreso Latinoamericano de Ecología y II Congreso Peruano de Ecología. Universidad San Agustín. Arequipa Perú. E- 8. [ Links ]
Montealegre, M. 1988. Condiciones meteorológicas predominantes en la región del Pacífico Sud-oriental adyacente a la Costa Suramericana durante el período 1987-1988. Memorias VI Seminario Nacional de Ciencias del Mar. Universidad Jorge Tadeo Lozano. Bogotá - Colombia. [ Links ]
Palacios, J., R. Cruz & O. Pacheco. 1983. Estructura poblacional y cuantificación de Donax dentifer Hanley, 1843 (Pelecypoda: Donacidae) en Playa Garza, Puntarenas, Costa Rica. Rev. Biol. Trop. 31: 163-166. [ Links ]
Pauly, D. 1979. Theory and management of tropical multiespecies stocks: a review whith emphasis on the Southeast Asian demersal fisheries. ICLARM Stud. Rev., (1): 35 pp. [ Links ]
Pauly, D. 1983. Lengt-Converted Catch Curves. A powerful tool for fisheries research in the tropics. Part l. Fishbyte 1(2): 9-13. [ Links ]
Riascos, J.M. & H.J. Urban. 1999. Morfología y aspectos ecológicos de una población de "chipi-chipi" (Donax dentifer Hanley, 1843 BIVALVIA:DONACIDAE) en Bahía Málaga, Pacífico Colombiano. Cespedesia 23(73-74): 63-77 [ Links ]
Rubio, E. 1984. Estudio taxonómico preliminar de la Ictiofauna de la bahía de Málaga. An. Inst. Inv. Mar. Plunta Betín 14: 157-173. [ Links ]
Sparre, R & S.C. Venema. 1995. Introducción a la evaluación de recursos pesquemos tropicales. Parte 1. Manual. FAO Documento Técnico de Pesca. No. 306.1 Rev. 1., 440 p. [ Links ]
Urban, H.J. & J.M. Riascos. 2002. Estimating Gonado-Somatic indices in bivalves with fused gonads. j. Shellfish Res. 21(1): 249-254. [ Links ]
Urban, H.J & C. Tesch. 1996. Aspects of the population dynamics of six bivalve species from Southern Chile. Results of the "Victor Hensen" Cruise to the Magellan Strait and the Beagle Chanel in October/November 1994. Arch. Fish. Mar. Res. 44(3): 243-256. [ Links ]
Urban, H-J. & J. Tarazona. 1996. Effects of El Niño/Southern Oscillation on the populations dynamics of a Gari solida population (Bivalvia: Psammobiidae) from Bahía Independencia, Perú. Mar. Biol. (125): 725-734. [ Links ]
Urban, H-J. 1996. Population dinamics of the bivalves Venus atiqua, Tagelus dombeii, and Ensis macha from Chile at 36ºS. J. Sellfish Res. (15): 3, 719-727. [ Links ]
Vakily, J.M. 1992. Determination and cornparison of bivalve growth, with emphasis on Thailand and other tropical arcas. ICLARM Tech. Rep. 36, 125 p. [ Links ]
1 . Instituto Vallecaucano de Investigaciones Científicas (INCIVA), Museo de Ciencias Naturales. Diagonal 28 # 30-11 San Fernando, Cali - Colombia. Fax: ++57-2-5583466; pesamoso@hotmail.com

