










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.50 n.3-4 San José Dec. 2002
Abstract
A study of the most important polliniferous plants for European and Africanized honeybees (Apis mellifera L.) was made in Quintana Roo state. Comparisons were made between the plants visited by both bee types in order to determine whether there were qualitative or quantitative differences in their choice of plant species. Also some foraging strategies of the honeybees were analysed. Pollen from pollen load samples was acetolysed and mounted on slides. Subsequently the polien grains were identified, counted and photographed. A total of 206 pollen load samples were collected at Palmas and St. Teresa during two years. The most frequent species in the ponen load samples from European and Africanized honeyhees were Cecropia peltata, Metopium brownei, Lonchocarpus sp. 2, Viguiera dentata, Eragrostis sp. 1, Bursera simaruba and Eupatorium albicaule. Both types of honey bees show a high reliance on pollen from only a few species, the first five named above comprised around 50% of all the mean percentage frequencies. Families that contributed with the largest number of polien species were Fabaceae, Asteraceac, Boraginaceae, Convolvulaecae, Euphorbiaceae, Sapindaceae, Poaceac, Myrtaceae, Sapotaceae and Tiliaceae. C. peltata, Trema micrantha, B. simaruba, Eugenia sp. 1, Thouinia canesceras, Pouteria sp. 1, Mimosa bahamensis and V. dentata, were the pollen species with the largest percentages of oceurrence in both European and Africanized bee pollen load samples, and also represent a "long-term" food resources during the year.
Keywords
Apis mellifera, Erpean honeybee, Africanized honeybee, foraging behaviour, Polliniferous plants, Yucatán Peninsula.
There is a growing interest among botanists, entomologists and ecologists to determine the food resources for bees in the tropics and also to understand the interrelationships between bees and the plants on which they forage. Some of the research in this area involves only observations of bees visiting flowers. Another type of research involves analysing the ponen that is present in bee nests or honey and pollen loads of foragers in order to determine which plants are visited.
Nectar and pollen are two of the rewards that plants offer to pollinators, so according to this, entomophilous plants can be divided into three groups: (1) nectariferous, (2) polliniferous and (3) nectariferous-polliniferous. In this work, the most importan polliniferous plants of the Yucatán Peninsula were determines, although many of the plants visited by the European and the Africanized honeybees were also anemophilous. Also foraging strategies of Apis mellifera are discussed in relation with the most important polliniferous plants.
Levin and Glowska-Konopacka (1963) and Winston (1987) found that a honeybee colony has a mean flight range of 1.7 km, with most foraging occurring within 6 km of the colony. In some cases, European and Africanized honeybees have heen observes foraging at a range of 10 km from their colonies (Visscher and Seeley 1982 , Roubik 1989 ), although no studies have been made to compare the flight distances of these two bee types in the same habitat. In this way, considering the mean flight range of the honeybee, intensive plant collections were made within a radius of 2 km from the sampled colonies in order to make a pollen reference collection and a pollen flora (Palacios et al. 1991 ), which would help to identify the pollen grains from the honey and the pollen load samples.
According to the classification of Koeppen (1936) , the type of climate that exists in the Yucatán Peninsula is Aw, which is defined as hot subhumid, with a mean annual temperature over 22 ºC and an annual precipitation between 700 and 1500 mm, and rainfall during the summer season.
Some of the most important nectariferous and polliniferous plants in the Yucatán Peninsula are from the northern and central part of this península. Sousa-Novelo et al. (1981) made a floristic list of the plant species present in the Yucatán Peninsula based on field observations. Roldan-Ramos (1984) analysed and describes the pollen grains found in honey samples taken from European honeybees and Melipona beecheii in Tixcacaltuyub, Yucatán. Villanueva-G. (1994) made comparisons of the nectar sources used by the European and the Africanized honeybees and also between the numbers of pollen and nectar flowers visited by bces from both bee types. The SAGAR (Anonymous 1998 ) describes the melliferous plants which beekeepers consider the most important. Villanueva-G. (1999) made a brief note about the most important pollen sources of European and Africanized honeybees.
There were two objectives in this study: (a) to identify the polliniferous plants most commonly used by honeybees (Apis mellifera L.) in the eastern Yucatán Peninsula, and (b) to determine some foraging strategies of European and Africanized honeybees as well as evaluate their diets.
Materials and methods
The research was made with pollen load samples taken from the European and the Africanized honeybees (Apis mellifera Linnaeus 1578) during two years, and an attempt has been made to determine through pollen analysis, the preferences of these honeyhees types for pollen of different flower species. The study took place in the eastern region of the Yucatán Peninsula, southern Mexico. There is an important beekeeping activity in this area. It hegan in September 1990, 3 years after the arrival of the feral Africanized honeybee in the study area. Two sites were chosen for the work. The first one, Palmas (site 1), is located 33 km south of the city of Felipe Carrillo Puerto (19º 09' N, 88º 09' W) and the second one, St. Teresa (site 2), is in the research station of the Biosphere Reserve of Sian Ka'an (19º 41' N, 87º 48'W).
Fieldwork
The types of vegetation that are present in both sites are: "selva baja subcaducifolia" (low sub-deciduous forest), "selva mediana subcaducifolia" (medium sub-evergreen forest) and secondary vegetation in different successional stages. These vegetation types are very common in most of the Yucatán Peninsula. The arcas surrounding Palmas are more disturbed than those in the immediate vicinity of St. Teresa. This is due to the fact that the first site is near a road and there is more agricultural activity. In St. Teresa there are only a few small apiaries, but in the environs of Palmas there is intense beekeeping activity.
In May 1987, the Africanized honeybee was detected in the south of the Yucatán Peninsula, in Quintana Roo state (Barrios Delgado et al. 1990 ). At both study sites, Palmas and St. Teresa, 15 hives contaming colonies headed by mated European honeybee queens, and 15 hives containing Africanized colonies derived from Africanized wild colonies (identified morphometrically by the method of Daly and Balling 1978 ) were selected. These Africanized colonies were collected from the surroundings of the study arcas. Pollen load samples were obtained from these hives for determination and quantification of polliniferous plants used by these two bee types. European and Africanized apiaries were separated 80 m from each other to prevent drifting. Both European and Africanized bee colonies were never fed artificially neither with pollen nor sugar-water rife.
From each study site, three European and three Africanized honeybee colonies were randomly selected for pollen load sampling. Modified Ontario Agricultural College (O.A.C.) pollen traps (Smith and Adie 1963 , Waller 1980 ) were placed at the base of the hives to sample the pollen loads that the honeybees carry on their hind legs. Pollen loads were collected for 48 hours each month during a period of two years (June 1989 to May 1991). A total of 206 pollen load samples were obtained in both sites (104 from European honeybees and 102 from Africanized honeybees).
Palynological Analysis from Pollen Load Samples
The collected pollen load samples were dried at 45º C for 24-48 hours until reaching constant weight and the final weight recorded. A subsample of 10 to 20 g of pollen was taken from each sample and soaked in 20 ml of distilled water and stirred magnetically for 1 hour. The pollen grains from the sample were further desegregated using a sonicator cell disrupter (O'Rourke and Buchmann method, 1991 ). The pollen was sonicated for 5 minutes at 24 kHz using a probe 'ultrasonic disintegrator' (M.S.E. SONIPREP) adjusted to medium power setting. In this way, each sample composition could be analysed in terms of (a) pollen percentage frequency, which was calculated by obtaining the pollen percentage by taxon from each sample, (b) mean percentage frequency was calculated by obtaining the mean of the percentage of each pollen species in all the samples either from European or Africanized honeybees, and c) the occurrence refers to the percentage of each pollen species in the total number of samples either from European or Africanized honeybees, considers only the presence or absence of a pollen species in a pollen load sample (see Villanueva G. 1999 ).
Micrographs of the Pollen Grains
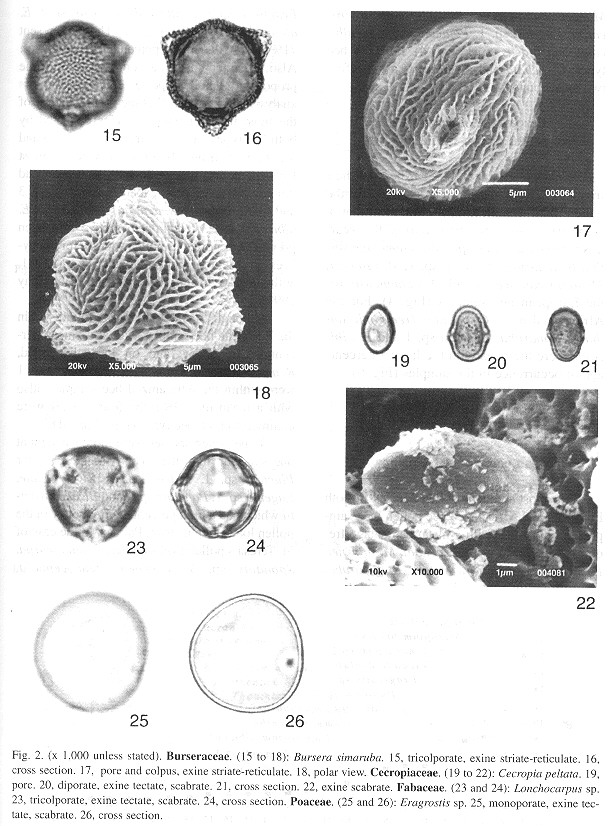
Photographs of the most abundant pollen grains in the pollen loads were taken. This was done with an optical light microscopic (Olympus BH-2) and a scanning electron microscope (SEM). A standard magnification of 1000 X has been used for most of the light micrographs, and a scale of 1mm:1mm has been maintained, so a measurement on the grain illustration in mm is the actual true size in microns (pm). This permits an easy measurement of the size of a pollen grain and a direct comparison between grains. Magnifications of all the photographs taken with the optic light microscope and with the SEM are indicated in the plate explanations (Figs. 1 and 2 ).
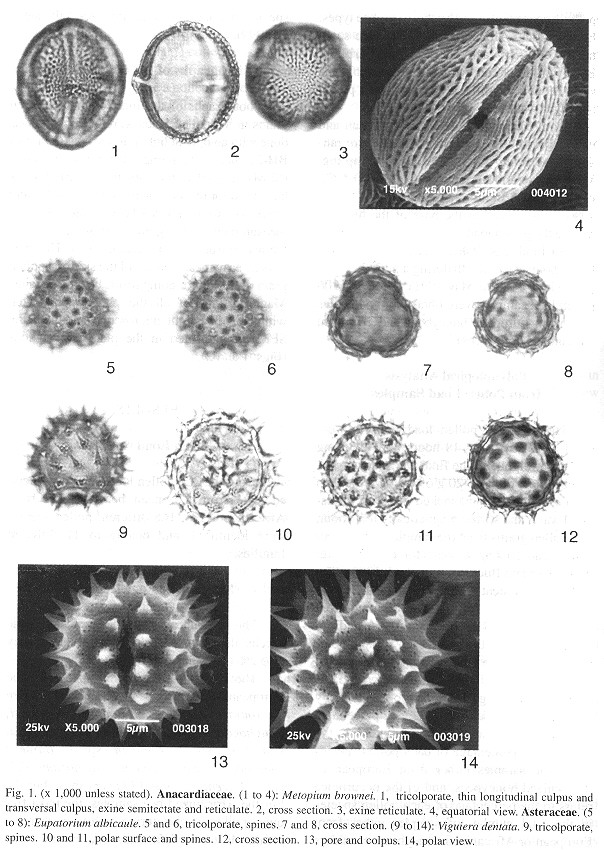
Pollen Load Analysis
From the 206 pollen load samples collected (104 from European bee and 102 from Africanized bee), 168 different pollen species were identified, and belong to 41 different families.
Mean percentage frequency
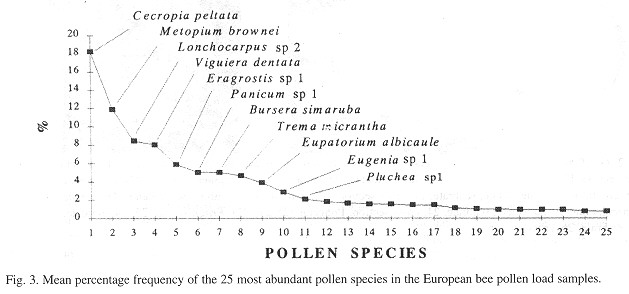
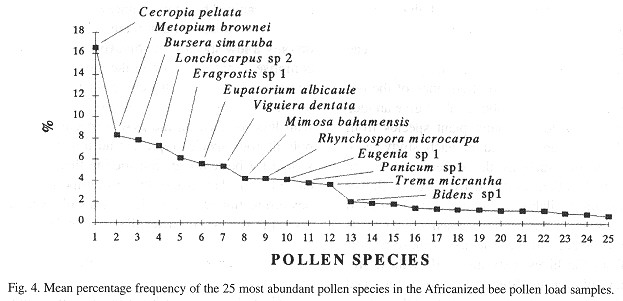
This account mostly concentrates on those species that have a mean percentage frequency of > 2% ( Figs. 3 and 4 ).
The most frequent pollen species in the European bee samples of years 1 and 2 were Cecropia peltata, Metopium brownei, Lonchocarpus sp. 2, Viguiera dentata, Eragrostis sp. 1, Panicum sp. 1, Bursera simaruba, Trema micrantha, Eupatorium albicaule, Eugenia sp. 1 and Pluchea sp. 1 (Fig. 3 ). For the Africanized bee samples, the most frequent pollen species during both years were C peltata, M. brownei, B. simaruba, Lonchocarpus sp. 2, Eragrostis sp. 1, E. albicaule, V dentata, Mimosa bahamensis, Rhynchospora microcarpa, Eugenia sp. 1, Panicum sp. 1, E. albicaule and Bidens sp. 1 (Fig. 4 ). For both bee types, C. peltata and M. brownei are the first two most abundant.
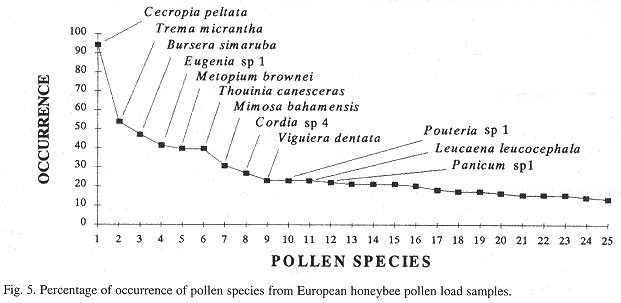
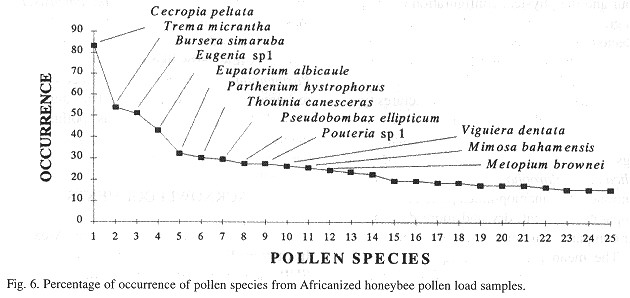
Percentage of occurrence of pollen species in the samples
Some pollen species (regardless of their frequency) were present in at least 30% of the pollen load samples, and also represent a "long-term" food resource during the year; this is the case with C. peltata, Trema micrantha, B. simaruba, Eugenia sp. 1 , M. brownei, Thouinia cabeceras and M. bahamensis for the European bee samples (Fig. 4 ). For the Africanized bees, C. peltata, Trema micrantha, B. simaruba, Eugenia sp. 1 and E. albicaule were the ones with the largest percentage of occurrence in the samples (Fig. 6 ).
Discussion
Mean percentage frequency
The most frequent pollen species in both European and Africanized bee pollen load samples in both sites (with a mean percentage frequency of > 2%) were C. peltata, M. brownei., Lonchocarpus sp. 2, V dentata, B. simaruba, Eragrostis sp. 1, E. albicaule, Panicum sp. 1, E. albicaule and Eugenia sp. 1. These represent 71% of all the mean percentage frequencies. Also, these ten species account for 89% of the proportion of pollen species considered in this analysis which means that the great majority of the most frequent pollen species collected by both bee types were the same ( Figs. 1 , 2 , 3 and 4 ). C. peltata and M. brownei were the most frequent pollen species in both European and Afficanized bees pollen load samples ( Figs. 3 and 4 ). M. brownei, Y dentata, B. simaruba, E. albicaule and M. bahamensis have also bcen previously reported as pollen sources for honeyhees in Mexico (Sousa-Novelo et al. 1981 , Villanueva-G. 1984 , Chemás and Rico-Gray 1991 ).
Although Pluchea sp. 1 was found within the European bee samples with a > 2 mean percentage frequency, and on the other hand, R. microcarpa, M. bahamensis and Bidens sp. 1 were within the Africanized bee samples (also with a mean of > 2%), the four species were common to both bee types (Figs 3 and 4 ).
Some pollen species were more frequent at one site than the other. This was the case for Pluchea sp. 1 , Bidens sp. 1, E. albicaule, Eugenia sp. 1, Panicum sp. 1 and Buxus bartlettii which had a high percentage frequency in the pollen load samples from Palmas. In the case of St. Teresa's pollen load samples, R. microcarpa, Spondias sp. 1, Leucaena leucocephala, Heliocarpus donnell-smithii, Fabaceac 2, Hibiscus sp. 2 and Conocarpus erecta were the more frequent. H. donnell-smithii and Fabaceae 2 were only present in St. Teresa.
The differences in the frequency of the pollen species mentioned above might give an idea of the abundance of many plant species from which they were derived in each study area. Other factors such as the differences in the amount of pollen produced by different flower species, the availability of pollen and nectar from flower species during days when pollen loads were sampled, and the distance of those resources from the hives, may also reflect these frequency differences. Some of these factors, and also the ones already mentioned about the colour, odour and morphology of the flowers, are the same as those probably determined the preference of honeybees for nectar and pollen resources.
Some families contributed with a large number of pollen species: Fabaceae, Asteraceae, Boraginaceae, Convolvulaceae, Euphorbiaceae, Sapindaceae, Poaceae, Myrtaceae, Sapotaceae and Tiliaceae. As with honey samples (Villanueva 1994 ), Fabaceae and Asteraceae families are very important in the bee diet, both families were well represented in the pollen load samples, with 12% and 11% of the total number of pollen species respectively.
According to Stanley and Linskens (1974) and Jay (1986) workers choose which pollen to collect not by their nutritive value, age, moisture, content, or colour, but on the basis of the odour and the physical configuration of the pollen grains, although many of the pollen species collected by honeybee foragers (some with high percentage frequency and high percentage occurrence) had a psilated exine (with a smooth surface), without spines or other structures typical of entomophilous plants. This was the case with the Cyperaceae and Poaceae pollen grains (Figs. 3 and 4 ), also M. bahamensis, Mimosa pudica and Chlorophora tinctoria. Shuel (1992) mentioned that anemophilous pollen tends to be comparatively light, dry and more drab in colour compared with entomophilous pollen.
The mean percentage frequency distribution of the pollen load sampled (from European and Africanized bees) was analysed (Figs. 3 and 4 ). lt can be observed that there is high reliance on only a few species, the first five for example comprise almost 50 % of all the mean percentage frequencies, and as the curves become linear, the dependency of the bees is more distributed, with less reliance on the first 5 species. These polliniferous species represent an important resource for beekeeping, considering that the Yucatán Peninsula is one of the most important honey production regions of the world with over 10 hives per square kilometer ( Paxton 1992 ).
Percentage of Occurrence
C. peltata, E. albicaule, B. simaruba,Eugenia sp. 1, T canesceras, Pouteria sp. 1, M. bahamensis and V dentata, were the pollen species with the largest percentage of occurrence in both European and Africanized bee pollen load samples, but none of the 168 pollen species identified was found to be present in all the 206 pollen load samples from either European or Africanized bees ( Figs. 5 and 6 ).
Some woody species (regardless of their high frequency or their large volume in the pollen load samples) are also important in the honeybee diet because they represent a constant source of pollen during most of the year. This is the case with trees like C. peltata, B. simaruba, M. brownei, T canesceras and Psidium sartorianum; shrubs like B. bartlettii and T micrantha, and shrubs or trees like Cordia sp. 4, Pouteria sp. 1 and Eugenia sp. 1 and M. bahamensis. All these species also had a high percentage of occurrence in the pollen load samples (Figs. 5 and 6 ). A few herbs like Eupatorium sp. 2 and Parthenium hysterophorus representad constant resources of pollen for the honeybees during the year, but their contribution as polliniferous plants was very small.
Acknowledgements
I want to acknowledge the Mexican Institutions El Colegio de la Frontera Sur (ECOSUR) and Consejo Nacional de Ciencia y Tecnología (CONACYT) for their great support in the development of the present work, and also I wish to thank the School of Pure and Applied Biology of the University of Wales College of Cardiff for the facilities I received during the development of this study. I would like to express my sincere thanks to Robert S. Pickard, John Free, Nicola Bradbear, Neil Kidd, Robert J. Paxton and David Roubik, for their useful comments on the manuscript. I would like to thank Wilberto Colli Ucán, who assisted me in the fieldwork. I am grateful to Rodolfo Palacios and Beatriz-Ludlow-Wiechers for their help in the identification of some of my pollen grains.
Resumen
Un estudio de las plantas poliníferas más importantes para las abejas europeas y africanizadas (Apis melifera L.) se realizó en el estado de Quintana Roo. Se hicieron comparaciones entre plantas visitadas por ambos tipos de abejas, con el objetivo de determinar si hay diferencias cualitativas o cuantitativas en la elección de la especie de planta. Adicionalmente, se analizaron algunas estrategias de forrajeo de las abejas. Muestras de cargas de pólen se acetolizaron y se montaron en láminas. Posteriormente, los granos de pólen se identificaron, se contaron y se fotografiaron. Se recolectaron un total de 206 muestras de cargas de pólen en Las Palmas y Sta. Teresa durante dos años. Las especies más frecuentes en las muestras de cargas de pólen fueron Cecropia peltata, Metopium brownei, Lonchocarpus sp. 2, Viguiera dentata, 1, Eragrostis sp. 1, Bursera simaruba y Eupatorium albicaule. Ambos tipos de abejas mostraron una alta dependencia hacia sólo unas pocas especies de plantas, las primetas cinco de las mencionadas anteriormente constituyen aproximadamente un 50% de todas las frecuencias de porcentaje promedio. Las familias que contribuyeron con un mayor número de especies de pólen fueron Fabaceae, Asteraceae, Boraginaceae, Convolvulaceae, Euphorbiaceae, Sapindaceae, Poaceae, Myrtaceae, Sapotaceae y Tiliaceae. En tanto que C. peltata, Trema micrantha, B. simaruba, Eugenia sp. 1, Thouinia canesceras, Pouteria sp. 1, Mimosa bahamensis y V. dentata, fueron las especies de pólen con los más altos porcentajes de presencia en las cargas de pólen de las abejas y representan además, una fuente de recursos alimenticios de larga duración durante todo el año.
References
Anonymous. 1998. Flora nectarífera y polinífera de la Península de Yucatán. COTECOCA-SAGAR, Mexico, D. E 128 p.
Barrios-Delgado, C., E. Pémz-Dominguez & L.M. Sánchez-Navarro. 1990. Efectos y repercusiones de la abeja africana en la Península de Yucatán. Memorias del IV Seminario Americano de Apicultura. Secreta, ría de Agricultura y Recursos Hidráulicos and Unión Nacional de Apicultores, Mexico. pp. 15-33. [ Links ]
Chemás, A. & Y Rico-Gray. 1991. Apiculture and management of associated vegetation by the Maya of Tixcacaltuyub, Yucatán, Mexico. Agrofo. Syst. 13: 13-25. [ Links ]
Daly, H. V. & S.S. Balling. 1978. Identification of Africanized honeybees in the Western Hemisphere by discriminant analysis. J Kansas Entomol. Soc. 5: 857-869. [ Links ]
Jay, S.C. 1986. Spatial management of honey bees on crops. Annu. Rev. Entomol. 31: 49-65. [ Links ]
Koeppen, W. 1936. Das gepographische System der Klimate In Handbuch der Klimatologie. Band I, Teil C; Berlin, Germany. 44 p. [ Links ]
Levin, M.D. & S. Glowska-Konopacka. 1963. Responses of foraging honeybees in alfalfa to increasing competition from other colonies. J. Apicult. Res. 2: 33-42. [ Links ]
Maheshwari, R 1950. An introduction to the embryology of Angiosperms. McGraw-Hill; New York, USA. 453 p. [ Links ]
O'Rourke, M.K. & S.L. Buchmann. 1991. Standardised analytical techniques for bee-collected pollen. Entomol. Soc. Amer. 20: 507-513. [ Links ]
Palacios-Chávez, R.B., R. Ludlow-Wiechers y G. Villanueva. 1991. Flora palinológica de la Reserva de la Biósfera de Sian Ka'an, Quintana Roo, México. Centro de Investigaciones de Quintana Roo; Quintana Roo, Mexico. 321 p. [ Links ]
Paxton, R.J. 1992. Africanized honeybees: new ideas in honeybee biology. p. 32-41. In J.B. Free (ed.). Keeping bees. Central Association of Beekeepers' Publications, Cardiff, United Kingdom. [ Links ]
Roldán-Ramos, L.A. 1984. Flora melífera de la zona de Tixcacaltuyub, Yucatán. Bch Thesis. Facultad de Ciencias, Universidad Nacional Autónoma de México, Mexico, D. F. 210 p. [ Links ]
Roubik, D.W. 1989. Ecology and natural history of tropical bees. Cambridge University, Cambridge, UK. 514 p. [ Links ]
Shuel, R.W. 1992. The production of nectar and pollen. pp. 401-436. In J. M. Graham (ed.). The hive and the honeybee. Dadant & Sons, Hamilton, III., USA. [ Links ]
Smith, M.V & A. Adie. 1963. A new design in pollen traps. Can. Bee J. 74: 4-8. [ Links ]
Sousa-Novelo, N.,V.M. Suárez-Molina & A. Barrera-Vázquez. 1981. Plantas melíferas y poliníferas de Yucatán. Fondo Editorial de Yucatán; Mérida, Yuc., Mexico. 60 p. [ Links ]
Stanley, R.G. & H.F. Linskens. 1974. Pollen: biology, biochemistry, management. Springer-Verlag; Berlin, Heidelberg and New York. 307 p. [ Links ]
Villanueva-G., R. 1984. Plantas de importancia apícola en el ejido de Plan del Río, Veracruz, México. Biofica. 9: 279-340.
Villanueva-G., R. 1994. Nectar sources of European and Africanized honeybees (Apis mellifera L.) in the Yucatán Peninsula, Mexico. J. Apicult. Res. 33: 44-58. [ Links ]
Villanueva-G., R. 1999. Pollen sources of European and Africanized honeybees in the eastern Yucatán Peninsula, Mexico. J. Apicult. Res. 38: 105-1 1 1. [ Links ]
Visscher, P.K. & T.D. Seeley. 1982. Foraging strategy of honey bee colonies in a temperate deciduous forest. Ecology. 63: 1790-1801. [ Links ]
Walter, G.D. 1980. A modification of the O.A.C. pollen trap. Amer. Bee J. 120: 119-121. [ Links ]
Winston, M. 1987. The biology of the honey bee. Harvard University, Cambridge, Mass., USA. 281 p. [ Links ]
* El Colegio de la Frontera Sur (ECOSUR), Apdo. Postal 424, C.P. 77049, Chetumal, Quintana Roo, Mexico. Fax: (983) 2 04 47. e-mail: rogel@ecosur-qroo.mx

