










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.50 n.3-4 San José Dec. 2002
Abstract
In northwestern Costa Rica, cattle are being used as a "management tool" to reduce the amount of combustible material, mainly dominated by Hyparrhenia rufa, an African grass. This project is being developed within Parque Nacional Palo Verde and Reserva Biológica Lomas Barbudal, both of which fonn part of the only remaining tropical dry forests in Mesoamerica. To determine the short-term effects of cattle grazing on the natural vegetation, we compared the floristic composition within Palo Verde in an area under intermittent cattle grazing with an area that has not been grazed. There were significantly fewer plant species in the area with intermittent cattle grazing compared to the area with no grazing. Floristic composition of these two habitats was different as reflected by both Fisher's alpha values and the Shannon index of diversity, both of which were significantly higher in the ungrazed site. The ungrazed area contained more plant species and was more similar to mature forest. The structure of the vegetation was significantly different between the intermittently grazed and ungrazed sites with more small stems (1-5 cm dbh) and fewer large stems (>5 cm dbh) in the intermittently grazed habitat. These results indicate that cattle grazing has an impact on the dry forest by reducing the relative abundance and density of larger tree species and by changing the species composition and structure of the community. The current management plan implemented in Palo Verde and Lomas Barbudal is not appropriate because of the impact that cattle have on the structure of the natural vegetation and should not be considered a viable alternative in other protected areas of dry forest in the Neotropics. We suggest that alternative fire prevention measures be evaluated including hand-cutting H. rufa, the creation of more frequent and larger fire breaks, and the development of green breaks.
Keywords
Cattle, Costa Rica, grazing, Hyparrhenia rufa, management, Palo Verde National Park, tropical dry forest.
Tropical dry forests are among the most endangered ecosystems in the world with less than 0.1% of the original dry forests of Pacific Mesoamerica under protection (Murphy and Lugo 1986, Janzen 1988 ). Deforestation for the development of cattle pastures has been the principal cause of tropical dry forest conversion throughout Mexico and Central America (Quesada 1974 , Toledo 1992 , Maass 1995 , Quesada and Stoner 2003 ). The conversion of tropical dry forests lo pasture lands results in a reduction in biodiversity (Lugo 1986 , Toledo 1988 ). Although most tropical dry forests are less biologically diverse compared to tropical humid or wet forests, their threatened status and high endemism elevate their conservation value (Toledo 1988 , Ceballos 1995 ). In addition lo reduction in species diversity (Guevara et al. 1992 ), other ecological consequences of tropical forest conversion include: (1) reduction in vegetative cover, (2) disruption of the water cycle, (3) changes in soil nutrients and status, and (4) soil compaction ( Buschbacher et al. 1988 , Uhl et al. 1988 , García-Oliva et al. 1994 , Maass 1995 , Johnson and Wedin 1997 ).
lt has been emphasized recently that despite Costa Rica's renowned conservation efforts of establishing protected areas (Boza 1993 ), the agriculture, cattle, and timber industries still threaten the biodiversity of this small country (Hunter 1994 , Vandermeer and Perfecto 1995 , Sánchez Azofeifa et al. 2001 , Quesada and Stoner 2003 , Stoner and Timm 2003 ). Although the establishment of extensive pastures for cattle grazing has been one of the major threats to tropical dry forests in Costa Rica, cattle are being used as a management tool within some protected areas of this country (Stoner and Timm 2003 , Quesada and Stoner 2003 ). The management plan implemented by the Tempisque Conservation Area, Guanacaste, includes the use of cattle within the boundaries of Parque Nacional Palo Verde and neighboring Reserva Biológica Lomas Barbudal ( Vaughan et al. 1995 ). The decision to use cattle to control fires and to promote regeneration of the forest was based on a survey that concluded that cattle grazing reduce tires by controlling the expansion of "African star-grass" (Hyparrhenia rufa) and favor the fast succession of dry forest ( Barboza 1995 , 1996 ). A review of cattle grazing at Palo Verde and its impact on wildlife is provided by Stoner and Timm (2003) .
Whether or not cattle significantly reduce the fire hazard remains questionable since this grass species is not consumed by cows during the dry season when it is tall, dry, and low in nutritional value (Daubenmire 1972 ). In the tropical dry forest of Brazil it has been demonstrated that as much as 60% of cattle's diet comes from browsing on seedlings, trees and shrubs during the dry season (Thomas et al. 1983 ). In Guanacaste, Costa Rica, cattle browse on at least 16 native tree and shrub species common to this tropical dry forest (Conklin 1987 ). Regardless if cattle are consuming H. rufa, it is important to document the effect that they may have on the structure and composition of the tropical dry forest before this practice is adopted in other protected areas within the Neotropics.
The objective of our research was to compare the vegetation within an ungrazed area of tropical dry forest with an area that has experienced intermittant cattle grazing to determine the short-term effects of cattle grazing on the vegetation. We compare species diversity, floristic composition, and vegetation structure in nearby sites with no grazing and with intermittent cattle grazing within a national park.
Materials and methods
Study site and land use history: Parque Nacional Palo Verde and Reserva Biológica Lomas Barbudal are located in the Tempisque watershed, Guanacaste Province, northwestern Costa Rica. These two protected sites cover an area of approximately 20 000 hectares and encompass a variety of habitats including dry deciduous forest, regenerating dry deciduous forest of various ages, riparian and spring forests, savanna, mesic forest, mangrove forest and wetlands ( Frankie et al. 1993 ). Annual rainfall is approximately 1.5 m and average annual temperature about 27º C (Maldonado et al. 1995 ).
The region of Palo Verde (more area than is presently park lands) was the Stewart family cattle ranch from 1923 until 1979 and has contained cattle under different densities throughout this time (D.A. Stewart, pers. com.). Approximately 18 000 cattle were rotated within an area of 1 00 000 ha including parts of the present-day national park (i.e. Laguna Palo Verde) from 1923-1949. These records also indicate a decline to an average of 7 000 head of livestock per year on 23 000 hectares from 1963-1979. During this period, the cattle were regularly rotated from uplands outside of the Palo Verde area during the rainy season to the Laguna Palo Verde and adjacent pastures during the dry season (D.A. Stewart, pers. com.). In 1975, 18 000 ha were expropriated from the Stewart family by the Costa Rica government as part of a settlement program (Mozo 1995 ). In 1977 part of the lands expropriated were designated as a national wildlife refuge (Executive Decree No. 6942-A, April 18, 1977) and in 1979 all cattle were removed from the newly created protected arca (D.A. Stewart, pers. com.). In 1990 the wildlife refuge and the adjacent arca were fused into a single park, Parque Nacional Palo Verde. In 1991, cattle were introduced to some areas of Palo Verde including the wetlands and flat lowlands. This was a result of the Tempisque Conservation Arca deciding to utilize cattle as a "management tool" to control fires and to manage the wetlands ( McCoy 1994 , Mozo 1995 ).
Our study sites were chosen based on aerial photographs of Parque Nacional Palo Verde from 1956, 1970, and 1986 (Instituto Geográfico Nacional, Costa Rica), and interpreted Landsat TM satellite images from 1992 (Maldonado et al. 1995 ). These photographs and images indicate that the lowlands and the hills between the ridges in the arca between Cerro La Carreta and Cerro El Jocote were covered with forest in 1970 and 1986. At the end of the 1980's severas fires went through this region of the park eliminating much of the forest cover originally present in the 1970 and 1986 aerial photographs ( Vaughan et al. 1995 ). In 1991, with the implementation of the management program in Palo Verde, cattle were introduced into the park and allowed to graze within a 200 ha fenced-in arca called La Carreta. Cattle were maintained within this arca from May until January and the density varied between 1.0 and 1.4 head per ha; occasionally cattle were removed when there was not sufficient food for them to eat within the arca (Barboza 1995 ).
Vegetation surveys were conducted in two sites (treatments) that were located near each other: (a) intermittently grazed; and (b) ungrazed. The arca of La Carreta where cattle were regularly introduced between 1990-1996 and allowed to graze was considered vegetation influenced by intermittent cattle grazing. This arca contained Hyparrenia rufa interspersed with shrubs and small trees. The arca between Cerro La Carreta and Cerro El Jocote where cattle were never observed nor were there signs such as tracks, manure, or trampled or grazed seedlings, was considered ungrazed.
H. ruffa was not observed in this arca. Interviews with park personnel and one of the former owners of the Palo Verde ranch (D.A. Stewart) indicate that cattle were not found in this arca either when it was a ranch, or more recently when the cattle program was reinitiated within Palo Verde. Since the aerial photographs showed both our grazed and ungrazed habitats to consist of forest in 1970 and 1986 and fires have passed through this entire arca in subsequent years, we assume that any differences in the vegetation between the two treatments are due to the effects of cattle grazing.
Vegetative sampling protocol: Data were collected in July, 1996. The method of variable arca sampling was used to compare floristic diversity, composition, and vegetation structure (Stern 1998 ) between the intermittently grazed and ungrazed sites. This vegetation rapid sampling method was developed for and used by the Conservation International Rapid Assessment Program (web site: http://www.conservation.org ). Sampled stems were divided into the following three size classes (diameter at breast height, 1.3 m; dbh): (a) 1.0-2.5 cm, (b) >2.5-5 cm, and (c) >5 cm. Six transects two meters wide were conducted within each type of habitat and all stems greater than 1.0 cm dbh were identified to species. A relatively narrow transect width of 2 m was selected to minimize oversampling spatially-clumped stems often of a single species. Each transect was continued until 50 stems were recorded for each of the three size classes, for a total of 150 stems (each stem was considered an individual). In two transects, both at the intermittently grazed site, 50 stems of the large size elass (>5.0 cm) were not found in which case the transect was discontinued after a distance of 1 km; these two cases both included > 40 stems. To compare size classes between the two sites, we recognized category (c) as >5-10 cm and later extrapolated a fourth category (d) of >10 cm based on field data. Plant species determinations were made in the field by local experts and with the aid of Janzen and Leisner (1980) and Gentry (1993) .
Data analysis: A chi-square test was used to determine differences between the intermittently grazed and ungrazed sites in terms of: (a) number of plant species, (b) number of stems in the small (1.0-2.5 cm dbh) and large (>5 cm dbh) size class of the fifteen most common species, and (c) number of stems in the small/medium (here lumped, 1.0-5.0 cm dbh) and large (>5 cm dbh) size class of the 20 species recognized by Hartshorn (1983) as representative of mature dry forest at Palo Verde.
Fisher's alpha was the algorithm used lo compare plant species diversity between the intermittently grazed and ungrazed samples; this test, based on species presence and not relative abundance, is appropriate and reliable for comparative purposes when sample sizes are relatively small (Condit et al. 1998 ). A Mann-Whitney U test was used to compare differences in Fisher's alpha between the six transects of the intermittently grazed and ungrazed sites. Fisher's alpha was also calculated for larger (>10 cm dbh) trees only, and for Hartshorn's (1983) tree data from Palo Verde mature forest. In 1970, Hartshorn (1983) established a 4 ha permanent plot in Palo Verde and all trees (>10 cm dbh) were identified. According to Hartshorn this forest had experienced little human intervention and cattle grazing had been minimal. The data obtained by Hartshorn (1983) represents the only plant species database of original mature forest available for this tropical dry forest. Finally, a Shannon index of diversity using the 6 replicates of each treatment was calculated lo compare diversity between the grazed and ungrazed sites and the Shannon t-test was used lo test for differences between these habitats ( Magurran 1988 ).
Results
The number of plant species was significantly different between sampling sites (X2 = 5.6, p < 0.025, df = 1). The ungrazed habitat had 84 plant species compared lo 56 species in the intermittently grazed area (Table 1 ). A Mann-Whitney U test comparing Fisher's alpha values from the 12 transects indicated that plant species diversity of these two sites was significantly different (W = 24, p < 0.01). Alpha values were significantly lower in the six transects where cattle grazed intermittently compared lo the alpha values calculated for the six transects with no grazing (7.56 versus 12.59 median ranks, respectively). Similarly, a total Fisher's alpha value calculated for each treatment revealed a two times greater diversity in the ungrazed habitat compared to the intermittently grazed area (Table 1 ). The Shannon diversity index also was significantly greater in the ungrazed sites than the intermittently grazed sites, again representing a higher species diversity in areas without grazing (t 3.14, p < 0.005, df = 743; Table 1 ).

When the fifteen most common species at each site were compared and divided into size classes, floristic differences between the two habitats were even greater (Fig. 1 ). Nine of the fifteen most common species in ungrazed habitat were not found in the fifteen most common species in the grazed area. Furthermore, the size class structure between the two habitats was significantly different (X 2 = 5.7, p < 0.016, df =1), with greater representation of small stems (1.0-2.5 cm) in the intermittently grazed area and a greater representation of large stems (>10 cm) in the ungrazed area ( Fig. 1 ).
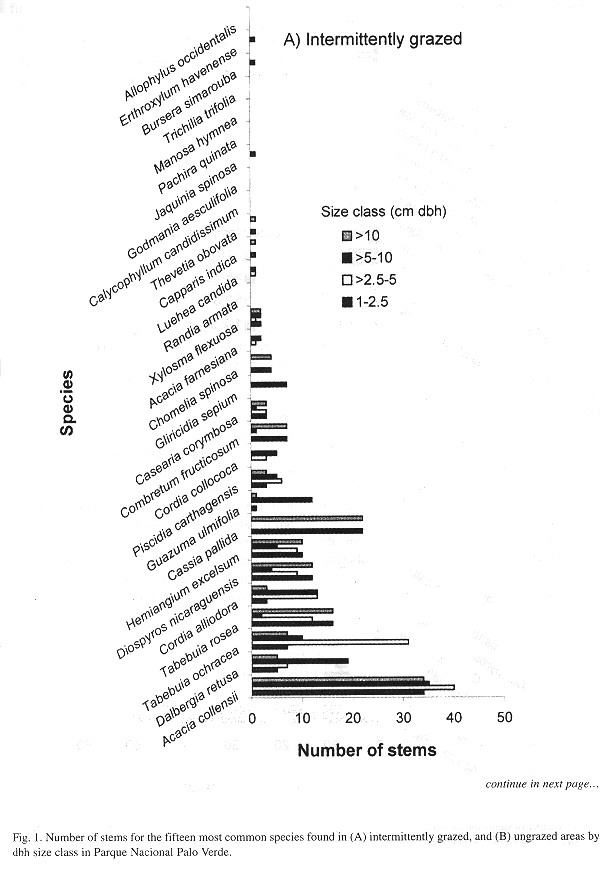

Further evidence of the impact that grazing has on the floristic composition and structure of the Palo Verde tropical dry forest was demonstrated by comparing Fisher's alpha values for large trees only from the grazed and ungrazed sites with Hartshorn's (1983) permanent tree plot data in Palo Verde mature forest ( Table 2 ). The ungrazed area is more similar to the forest in the Hartshorn plot than to the nearby intermittently grazed site. lt should be noted that the Hartshorn plot is located approximately 20 km from our sampling area and data were collected in 1970.

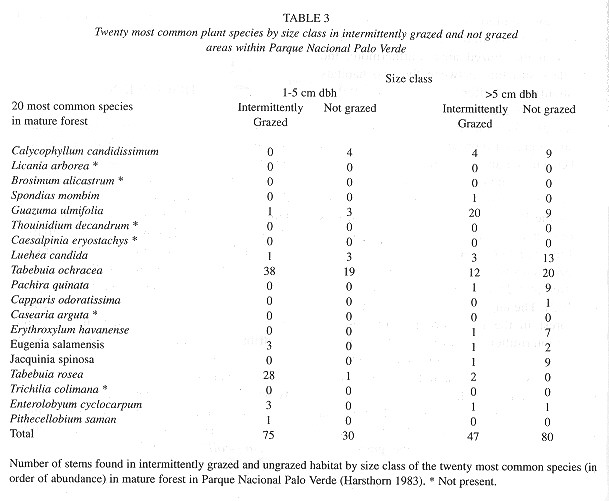
Finally, there was a significant difference in the number of small stems (1.0-5 cm) and large stems and trees (> 5 cm) in the grazed and ungrazed treatments when comparing only the 20 species most commonly found in Hartshorn's mature forest (X2 = 27.3, p < 0.0001, df = 1; Table 3 ). While the intermittently grazed area had a greater abundance of small stems (of the 20 species considered) than the ungrazed area (75 versus 30 stems, respectively), this ratio changed as the cohort aged (large stems, 30 versus 80 stems, respectively) as many small stems were possibly eaten by cattle, trampled, or killed by other factors.
Discussion
Impact of cattle grazing on vegetation structure and composition: We demonstrate that the floristic structure and composition of the intermittently grazed sites sampled were different from the sites sampled with no cattle grazing. The analysis of floristic composition showed significantly lower species diversity at the sites where intermittent cattle grazing occurred than at the sites with no grazing ( Table 1 ). Although the habitat with intermittent grazing was less diverse at the species level, 14 species were unique to this area thus contributing to the overall diversity of the landscape; however, the area with no grazing contributed more to the overall species diversity with 42 species unique to this habitat.
Species that were abundant in both intermittently grazed and ungrazed areas, included Acacia collinsii, Dalbergia retusa, and Tabebuia ochraceae. Some of the most abundant species in the intermittently grazed area included severas early successional species such as Tabebuia rosea, Cassia pallida, and Piscidia carthagenensis. In contrast, severas of the most common species in the ungrazed area included species characteristic of mature tropical dry forest ( Hartshorn, 1983 ) such as Luehea candida, Calycophyllum candidissimum, Jacquinia spinosa, and Pachira quinata. The significant difference in size class distributions between intermittently grazed and ungrazed areas emphasizes the impact that cattle have on the structure of tropical dry forest ( Fig. 1 ). There were significantly more small stems in the intermittently grazed area and significantly fewer large stems. The low abundance of even the smallest size class of species that are characteristic of mature dry forest in the intermittently grazed area suggests that cattle grazing have a detrimental effect on the regeneration of tropical dry forest, and that the impact of cattle may direct a change in the specíes composition of the forest as the small class matures.
Of the twenty most common tree species in the Hartshorn plot from mature forest within Palo Verde, six species were not found in any of our transects. Twelve of the 20 most common species from the Hartshorn plot were found in our transects from both intermittently grazed and ungrazed sites. The ungrazed sites contained 12 common mature forest species in greater abundance's and from larger size classes than in transects from the intermittently grazed sites. The species diversity (Fisher's Alpha values) of the Hartshom plot was intermediate between the diversity of our ungrazed and intermittently grazed sites. A possible explanado for the highest diversity in the ungrazed area is that this habitat represents a mosaic of both pioneer and mature forest species because this area is regenerating from fires.
Since aereal photographs indicate that both our intermittently grazed and ungrazed areas consistes of forest in 1970 and 1986, and fires probably affected both of these areas equally since they are adjacent lo each other, the subsequent changes in vegetation are most likely due to the effect of cattle grazing. Alternatively, severas other factors undoubtedly have contributed to the differences observed in these two habitats. The presence of H. rufa in the intermittently grazed area may prevent the establishment of other plants since grasses often inhibit the establishement of woody and herbaceous species in open areas ( Guariguata et al. 1995 , Zahawi and Augspurger 1999 ). Furthermore, grazing promotes the horizontal expansion of H. rufa, allowing this aggressive grass to outeompete other woody colonizers ( Parsons 1972 , McNaughton 1985 , Milchunas et al. 1988 ). Finally, abiotic factors encountered by seeds in the more open areas in the intermittently grazed habitat likely decrease the survival of small seedlings in this area (Nepstad et al. 1990 , Gerhardt 1993 , Nespstad et al. 1996 ).
Many studies in western North America have demonstrated that grazing by domestic livestock in native ecosystems results in changes in plant species composition, disruption of ecosystem function, alteration of ecosystem structure, and changes in faunal diversity (for a review see Tables 1-4 in Fleischner 1994 ). In general, these studies show a decrease in plant species diversity accompanied by a change in floristic composition and structure within areas grazed by domestic livestock (Cottam and Evans 1945 , Gardner 1950 , Rummell 1951 , Blydenstein et al. 1957 , Winegar 1977 , Webb and Stielstra 1979 , Reynolds and Trost 1980 , Szaro and Pase 1983 , Bock et al. 1984 , Brady et al. 1989 , Schulz and Leiniger 1990 ). Furthermore, numerous researchers have documented the negative impacts of cattle grazing on native vertebrate populations (Berry 1978 , Duff 1979 , Reynolds and Trost 1980 , Jones 1981 , 1988 , Crouch 1982 , Mosconi and Hutto 1982 , Szaro et al. 1985 , Taylor 1986 , Taylor and Littlefield 1986 , Campbell 1988 , Medin and Clary 1989 , Bowen and Kruse 1993 , Stoner and Timm 2003 ).
Studies evaluating the impact of domestic grazing on tropical ecosystems also have documented changes in vegetation structure and composition. In the Western Ghats, Kamataka, India, dry forests expanded into former pasture/savanna habitats when protected from human impacts of fire and grazing (Puyravaud et al. 1994 ). In the eastern Amazonian rainforest of Pará, Brazil, the success of forest regeneration in pasture lands depended on the intensity of pasture use. Furthermore, the composition and structure of recovered mature forest was not always similar to the mature forest prior to land clearing ( Buschbacher et al. 1988 , Uhl et al. 1988 ). A study in Curagao, Netherlands Antilles, documented that vegetation composition and structure was significantly different between rock vegetation with livestock grazing compared to rock vegetation with no livestock ( Debrot and Freitas 1993 ).
Implications for management: The management objectives of the Tempisque Conservation Arca for the cattle project in Parque Nacional Palo Verde and Reserva Biológica Lomas Barbudal include the following: (1) reduce the risk of forest fires by managing the amount of combustible material, (2) minimize the impact of fire on biodiversity, (3) facilitate ecological restoration by using controlled grazing, and (4) generate a socio-economic benefit for participating cattle owners ( Mozo 1995 ). A parameter that was not considered in the management objectives of the Tempisque Conservation Arca is the impact that cattle have on the regeneration of tropical dry forest.
The management plan that is currently being implemented, claims that the Palo Verde area has been exposed to disturbance caused by cattle grazing during the last 400 years (Vaughan et al. 1995 ), however, no data is presented to confirm this statement. McCoy (1994) and Vaughan et al. (1995) argue that this disturbance has maintained a high diversity of plant species in dry forest communities and interpret this phenomenon as a result of the intermediate disturbance hypothesis proposed by Connell (1978) . Therefore, according to the management plan, cattle are essential for the regeneration of tropical dry forest communities and must be maintained in these areas perpetually (Vaughan et al. 1995 ).
In contrast to the management plan, the earliest records of cattle in the Palo Verde region come from 1923 when the Stewart family began a cattle ranch in this area (D.A. Stewart, pers. com.). Furthermore, it is difficult to interpret the disturbance of cattle at Palo Verde in the context of Connell's intermediate disturbance hypothesis as suggested by the management plan. This hypothesis suggests that highest diversity will be observed at intermediate levels of frequency and intensity of natural disturbances (Connell 1978 ). Cattle are not native animals of tropical dry forests of Mesoamerica and they do not represent a natural disturbance. Their presence within an area creates a constant disturbance in terms of both frequency and intensity that is determined by human intervention. The intensity of land use (i.e. disturbance by cows) depends largely upon the density of animals in a given area and the availability of forage. These two variables are controlled by human interests. For example, the density of cattle in the Palo Verde region when it was managed as a cattle ranch from 1923-1979 varied from 0.18-0.30 head of cattle per hectare. Since the implementation of the cattle program in Palo Verde and Lomas Barbudal the density has increased to 0.7-1.0 head of cattle per hectare (Mozo 1995 ), for a total of approximately 4000 cattle in 1998. In contrast to the cattle rotation schedule utilized by the Stewart family when Palo Verde was a ranch, the current management plan assigns specific sites to the participating cattle owners that result in areas with high animal density and no rotation. This practice may cause an incalculable damage to the structure and dynamics of this tropical dry forest.
An analysis of change in land-use of the area of Palo Verde and Lomas Barbudal was obtained using interpretad images of aerial photographs from 1956 and compared to satellite images interpretad in 1993 (Maldonado et al. 1995 ). This analysis shows that the area covered with pastures increased from 5% in 1956 to 12% in 1993 within Palo Verde and Lomas Barbudal. In contrast, Parque Nacional Guanacaste, another tropical dry forest also located in northwestem Costa Rica, has experienced an overall decrease in pasture land from 1979 to 1985. In only 6 years, 28% of the pasture land became successional deciduous or evergreen forest (Kramer 1997 ). Restoration of pastures to forest in Parque Nacional Guanacaste depends upon the distance to the seed sources, the seed dispersal mode of the plant species, the proximity to the forest patches, and the number and species of animals involved (Janzen 1986 ). Cattle, which were formerly present in this park, have been eliminated from this area and are not considered to be a viable management tool for controlling tires.
Our study shows that cattle grazing changes the floristic composition and decreases the structural complexity of the vegetation in tropical dry forest. Although the ultimase long-term impact of cattle grazing on tropical dry forest ecosystems is still unknown, there is no doubt that cattle are a risk factor to the maintenance of biological diversity of dry tropical forest vegetation. The long-term ecological cost of preventing fires with the use of cattle will be much greater than the short-term economic cost of investing in alternative fire manamenent programs. The effectiveness of alternative fire prevention measures should be evaluated, such as: (1) hand-cutting H. rufa with machetes, (2) the creation of more frequent and larger fire breaks, and (3) the development of "green" fire breaks consisting of fast-growing fire-resistant native plant species. The economic cost of cach of these alternatives should be evaluated in the context of available resources and infrastructure. Concurrently, community-based environmental education programs that promote safe burning practices should be continued and expanded. Finally, future studies should determine the effect of fire on the regeneration of this ecosystem to better understand the interaction between grazing and fire. The strategy of utilizing cattle to "manage" natural ecosystems in the rangelands of western North America has been criticized previously as a large uncontrolled experiment (Bock et al. 1993, Noss 1994). it now appears that the Tempisque Conservation Area in Guanacaste, Costa Rica, is similarly initiating an uncontrolled experiment within one of the few remnants of tropical dry forests in Central America.
Acknowledgments
We thank the student participants of OTS 96-9 (Tropical Plant Systematics) for help with field work. The help of N. Zamora, U. Chavarría, and R. Espinoza for field assistance and plant identifications is also gratefully acknowledged. We thank J. Stanford, R. M. Timm, and an anonymous reviewer for helpful comments on an earlier draft of this manuscript.
Resumen
En el noroeste de Costa Rica se utiliza ganado como una "herramienta de manejo" para reducir la cantidad de material combustible, principalmente Hyparrhenia rufa, un zacate que proviene de Africa. Este proyecto se desarrolla dentro del Parque Nacional Palo Verde y la Reserva Biológica Lomas Barbudal, ambos forman parte de los últimos remanentes de bosque tropical seco de Mesoamérica. Con el fin de determinar los efectos a corto plazo del pastoreo de ganado vacuno sobre la vegetación natural, realizamos un muestreo dentro de Palo Verde para comparar la composición florística de un área de pastoreo intermitente con un área sin pastoreo. Encontramos significativamente menos especies de plantas en el área con pastoreo intermitente comparado con el área sin pastoreo. La composición florística de estos dos hábitats fue significativamente diferente basada en una comparación del alfa de Fisher y el índice de Shannon de diversidad; ambos fueron significativamente más altos en el área sin pastoreo. El área no pastoreado tuvo mayor número de especies y mayor semejanza con bosque maduro. La estructura de la vegetación fue significativamente diferente entre las dos áreas pastoreadas y no pastoreadas, con mayor número de individuos de diámetros pequeños (1-2.5 cm) y menor número de individuos de diámetros grandes (>5 cm) en el área con pastoreo intermitente. Estos resultados sugieren que el ganado reduce la abundancia relativa y la densidad de las especies de árboles y cambia la composición florística y estructura del bosque tropical seco. El plan de manejo actual de Palo Verde y Lomas Barbudal no es apropriado debido al impacto que el ganado tiene sobre la estructura de la vegetación natural y no debe ser considerado como una alternativa viable para otras áreas protegidas del neotropico. Sugirimos que se evalúen métodos alternativos para prevenir incendios, como el corte de jaragua (Hyparrhenia rufa) con machetes, la creación de rompefuegos en mayor cantidad y magnitud, y rompefuegos verdes.
References
Barboza, G. 1995. Pastoreo controlado para control de incendios y restauración de bosque tropical seco, en el Parque Nacional Palo Verde, Guanacaste, Costa Rica. Informe de avance de resultados de monitoreo del proyecto, 12 de junio de 1995. Organización para Estudios Tropicales, San José, Costa Rica. [ Links ]
Barboza, G. 1996. Pastoreo controlado evita incendios y ayuda a ganaderos. Organización para Estudios Tropicales, OET al Día 1: 4.
Berry, K.H. 1978. Livestock grazing and the desert tortoise. Trans. North Amer. Wildlife Natur. Resources Conf. 43: 505-519. [ Links ]
Blydenstein, J., C.R. Hungerford, G.I. Day & R.R. Humphrey. 1957. Effect of domestic livestock exclusion on vegetation in the Sonoran Desert. Ecol. 38: 239-242. [ Links ]
Bock, C.E., J.H. Bock, W.R. Kenney & VM. Hawthome. 1984. Responses of birds, rodents, and vegetation to livestock exclosure in a semidesert grassland site. J. Range Manage. 37: 239-242. [ Links ]
Bock, C.E., J.H. Bock & H.M. Smith. 1993. Proposal for a system of federal livestock exclosures on public rangelands in the western United States. Conserv. Biol. 7: 731-733. [ Links ]
Bowen, B.S. & A.D. Kruse. 1993. Effects of grazing on nesting by upland sandpipers in southcentral North Dakota. J. Wildlife Manage. 57: 291-301. [ Links ]
Boza, M.A. 1993. Conservation in action: past, present, and future of the national park system of Costa Rica. Conserv. Biol. 7: 239-247. [ Links ]
Brady, W.W., M.R. Stromberg, E. F. Aldon, D.D. Bonham & S.H. Henry. 1989. Response of a semidesert grassland to 16 years of rest from grazing. J. Range Manage. 42: 284-288. [ Links ]
Buschbacher, R., C. Uhl, E.A.S. Serráo. 1988. Abandoned pastures in eastern Amazonia. 11. Nutrient stocks in the soil and vegetation. J. Ecol. 76: 682-699. [ Links ]
Campbell, F.T. 1988. The desert tortoise. pp. 567-581. In J. Chandler (ed.). Audubon Wildlife Report 1988-1989. Acadeniic, San Diego, Califomia.
Ceballos, G. 1995. Vertebrate diversity, ecology, and conservation in neotropical dry forests pp. 195-220. In S.H. Bullock, H.A. Mooney & E. Medina (eds.). Seasonally dry tropical forests. Cambridge University, Cainbridge, United Kingdom.
Condit, R., R.B. Foster, S.P. Hubt>ell, R. Sukumar, E.G. Leigh, N. Manokaran, S. Loo de Lao, J.V LaFrankie & P.S. Ashton. 1998. Assessing forest diversity on small plots: calibration using species-individual curves from 50-ha plots, pp. 247-268. In F. Dallmeier & J.A. Comiskey (eds.). Forest biodiversity research, monitoring and modeling: conceptual background and Old World case studies, UNESCO and Parthenon Publishing, Paris, France. [ Links ]
Conklin, N.L. 1987. The potential nutritional value to cattle of some tropical browse species from Guanacaste, Costa Rica. Ph.D. thesis, Cornell University, New York. [ Links ]
Connell, J.H. 1978. Diversity in tropical rainforests and coral reefs. Science 199: 1302-1310. [ Links ]
Cottam, W.P. & F.R. Evans. 1945. A comparative study of the vegetation of grazed and ungrazed canyons of the Wasatch Range, Utah. Ecol. 26: 171-181. [ Links ]
Crouch, G.L. 1982. Wildlife on ungrazed and grazed bottom-lands on the South Platte River, northeastem Colorado, pp. 186-198. In L. Nelson, J.M. Peek & P.D. Dalke (eds.). Proceedings of the wildlife-livestock relationships syniposium. Forest, Wildlife, and Range Experiment Station, University of Idaho, Moscow, Idaho. [ Links ]
Daubenmire, R. 1972. Some ecologic consequences of converting forest to savanna in northwestern Costa Rica. Trop. Ecol. 13: 31-51. [ Links ]
Debrot, A.O. & J.A. de Freitas. 1993. A cornparison of ungrazed and livestock-grazed rock vegetations in Curagao. Biotropica 25: 270-280. [ Links ]
Duff, D.A. 1979. Riparian habitat recovery on Big Creek, Rich County, Utah, pp. 91-92. In O.B. Cope (ed.). Proceedings of the Forum-grazing and riparian/stream ecosystems. Trout unlimited, Denver, Colorado. [ Links ]
Fleischner, T.L. 1994. Ecological costs of livestock grazing in western North America. Conserv. Biol. 8: 629-644. [ Links ]
Frankie, G.W., L. Newstrom, S.B. Vinson & J.F. Barthell. 1993. Nesting-habit preferences of selected Centris bee species in Costa Rican dry forest. Biotropica 25:322-333. [ Links ]
García-Oliva, F., I. Cesar, P. Morales & J.M. Maass. 1994. Forest-to-pasture conversion influecnes on soil organic carbon dynamics in a tropical deciduous forest. Oecologia 99: 392-396. [ Links ]
Gardner, J.L. 1950. Effeets of thirty years of protection from grazing in desert grassland. Ecol. 31: 44-50.
Gentry A.H. 1993. A field guide to the families and genera of woody plants of northwest South America (Colombia, Ecuador, Peru) with supplementary notes on herbaceous taxa (R. Vásquez, illustrator). The University of Chicago Press, Chicago, Illinois. [ Links ]
Gerhardt, K. 1993. Tree seedling development in tropical dry abandoned pasture and secondary forest in Costa Rica. J. Veg. Sci. 4: 45-102. [ Links ]
Guariguata, M.R., R. Rhenigans & E Montagnini. 1995. Early woody invasion under tree plantations in Costa Rica: implicaions for forest restoration. Rest. Ecol. 3: 252-260. [ Links ]
Guevara, S., J. Meave, P. Moreno-Cassasola & J. Laborde. 1992. Floristic composition and structure of vegetation under isolated trees in neotropical pastures. J. Veg. Sci. 3: 655-664. [ Links ]
Hartshorn, G.S. 1983. Plants: Introduction, p. 11 8-157. In D. H. Janzen (ed.). Costa Rican natural history. University of Chicago, Chicago, Illinois. [ Links ]
Hunter, J.R. 1994. Is Costa Rica truly conservation-minded? Conserv. Biol. 8: 592-595. [ Links ]
Janzen, D.H. 1986. Guanacaste National Park: tropical ecological and cultural restoration. Editorial Universidad Estatal a Distancia, San José, Costa Rica. [ Links ]
Janzen, D.H. 1988. Tropical dry forests: The most endangered major tropical ecosystem, pp. 130-137. In E.O. Wilson (ed.). Biodiversity, National Academy Press, Washington, D.C. [ Links ]
Janzen, D.H. & R. Liesner. 1980. Annotated check-list of plants of lowland Guanacaste Province, Costa Rica, exclusive of grasses and non-vascular cryptogains. Brenesia 18: 15-90. [ Links ]
Johnson, N.C. & D.A. Wedin. 1997. Soil carbon, nutrients, and mycorrhizae during conversion of dry tropical forest to grassland. Ecol. Applications 7: 171-182. [ Links ]
Jones, K.B. 1981. Effects of grazing on lizard abundance and diversity in western Arizona. Southwestern Naturalist 26: 107-115. [ Links ]
Jones, K.B. 1988. Comparison of herpetofaunas of a natural and altered riparian ecoysystem, pp. 222-227. In R.C. Szaro, K.E. Severson & D.R. Patton (eds.). Management of amphibians, reptiles, and small mammals in North America: General Technical Report RM- 166. U.S. Forest Service, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colorado. [ Links ]
Kramer, E. 1997. Measuring landscape changes in remnant tropical dry forest, pp. 386-399. In W.F. Laurance & R.O. Bierregaard, Jr. (eds.). Tropical forest remnants: ecology, management and conservation of fragmented communities. The University of Chicago Press, Chicago, Illinois. [ Links ]
Lugo, A.E. 1986. Estimating reductions in die diversity of tropical forest species, pp. 58-70. In E.O. Wilson (ed.). Biodiversity. National Academy, Washington D.C. [ Links ]
Maass, J.M. 1995. Conversion of tropical dry forest to pasture and agriculture, pp. 399-422. In S.H. Bullock, H.A. Mooney & E. Medina (eds.). Seasonally dry tropical forests. Cambridge University, Cambridge, United Kingdom. [ Links ]
Magurran, A.E. 1988. Ecological diversity and its measurement. Princeton University, Princeton, New Jersey. [ Links ]
Maldonado, T., J. Bravo, G. Castro, Q. Jiménez, O. Sahorío & L. Paniagua. 1995. Evaluación ecológica rápida región del Tempisque Guanacaste, Costa Rica. Fundación Neotrópica, San José, Costa Rica. [ Links ]
McCoy, M.B. 1994. Seasonal, freshwater marshes in the tropics: a case study in which cattle grazing is not detrimental, Essay 12C, pp. 352-353. In G.K. Meffe & C.R. Carroll (eds.). Principles of conservation biology. Sinauer Associates, Sunderland, Massachusetts. [ Links ]
McNaughton, S.J. 1985. Ecology of a grazing ecosystem: The Serengeti. Ecol. Monogr. 55: 259-294. [ Links ]
Milchunas, D.G., O.E. Sala & W.K. Lauenroth. 1988. A generalizad model of the effects of grazing by large herbivores on grassland community structure. Am. Nat. 132: 87-106. [ Links ]
Medin, D.E., and W.R Clary. 1989. Small mainmal populations in a grazed and ungrazed riparian habitat in Nevada. Research Paper INT-413. U.S. Forest Service, Intermountain Research Station, Odgen, Utah. [ Links ]
Mosconi, S.L. & R.L. Hutto. 1982. The effect of grazing on the land birds of a westem Montana riparian. pp. 221-233. In L. Nelson, J.M. Peek & P.D. Dalke (eds.). Proceedings of the wildlife-livestock relationships symposium. Forest, Wildlife, and Range Experiment Station, University of Idaho, Moscow, Idaho. [ Links ]
Mozo, E.T. 1995. Pastoreo con ganado vacuno, una alternativa del ACT para prevención de incendios forestales, recuperación de humedales y restaruración del bosque tropical seco. Convenio MIRENEM-Opción Colombia, Universidad Sergio Arboleda, Bagaces, Costa Rica. [ Links ]
Murphy, RG. & A.E. Lugo. 1986. Ecology of tropical dry forest. An. Rev. Ecol. Sys. 17: 67-88. [ Links ]
Nepstad, D.C., C. Uhl & E.A.S. Serráo. 1990. Surmounting barriers to forest regeneration in abandoned highly degraded pastures: a case study form the Paragominas, Pará, Brazil, pp. 215-229. In A.B. Anderson (ed.). Altematives to deforestation: steps toward sustainable use of the Amazon rain forest. Columbia University, New York, New York. [ Links ]
Nepstad, D.C., C. Uhl, C.A. Pereira & J.M.C. Da Silva. 1996. A comparative study of tree establishment in abandoned pasture and mature forest of eastern Amazonia. Oikos 76: 25-39. [ Links ]
Noss, R.F. 1994. Cows and conservation biology. Conserv. Biol. 8: 613-616. [ Links ]
Parsons, J.J. 1972. Spread of African pasture grasses to the American tropics. J. Range Manage. 25: 12-17. [ Links ]
Puyravaud, J.R, J.R Pascal & C. Bufour. 1994. Ecotone structure as an indicator of changing forest-savanna boundaries. J. Biogeogr. 21: 581-593. [ Links ]
Quesada, M. & K.E. Stoner. 2003. Threats to the conservation of tropical dry forest in Costa Rica. In G.W. Frankie, A. Mata & S.B. Vinson (eds.). Biodiversity Conservation in Costa Rica: Learning the leasons in the seasonal dry forest. University of California Press, Berkeley, California. [ Links ]
Quesada, R. 1974. La forja de una nación, Tomo II. Tesis de Licenciatura en derecho. Universidad de Costa Rica, San José, Costa Rica.
Reynolds, T.D. & C.H. Trost. 1980. The response of native vertebrate populations to crested wheatgrass plantings and grazing by sheep. J. Range Manage. 33: 122-125. [ Links ]
Rummel, R.S. 1951. Some effects of livestock grazing on ponderoso pine forest and range in central Washington. Ecol. 32: 100-103. [ Links ]
Sánchez-Azofeifa, G.A., R.C. Harris & D.L. Skole. Deforestation in Costa Rica: A quantitative analysis using remote sensing imagery. Biotropica 33: 378-384. [ Links ]
Schulz, T.T. & W.C. Leininger. 1990. Differences in riparian vegetation structure between grazed areas and exclosures. J. Range Manage. 43: 295-299. [ Links ]
Stern, M.J. 1998. Field comparisons of two rapid vegetation assessment techniques with permanent plot inventory data in Amazonian Peru, pp. 273-288. In E Dallmeier & J.A. Comiskey (eds.). Forest biodiversity research, monitoring and modeling: conceptual background and Old World case studies. UNESCO and Parthenon, Paris, France. [ Links ]
Stoner, K.E. & R.M. Timm. 2003. Tropical dry forest mammals: Conservation priorities in a changing landscape. In G.W. Frankie, A. Mata & S. B. Vinson (eds.). Biodiversity Conservation in Costa Rica: Learning the leasons in the seasonal dry forest. University of California, Berkeley, California. [ Links ]
Szaro, R.C. & C.P. Pase. 1983. Short-term changes in a cottonwood-ash-willow association on a grazed and an ungrazed portion of Little Ash Creek in Central Arizona. J. Range Manage. 36: 382-384. [ Links ]
Szaro, R.C., S.C. Belfit, J.K. Aitkin & J.N. Rinne. 1985. Impact of grazing on a riparian garter snake, pp. 359363. In R.R. Johnson, C.D. Ziebell, D.R. Patton, P.F. Folliott & EH. Hamre (eds.). Riparian ecosystems and their management: reconciling conflicting uses, General Technical Report RM- 120, U. S. Forest Service, Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colorado. [ Links ]
Taylor, D.M. 1986. Effects of cattie grazing on passerine birds nesting in riparian habitat. J. Range Manage. 39: 254-258. [ Links ]
Taylor, D.M. & C.D. Littlefield. 1986. Willow flycatcher and yellow warbler response to cattle grazing. American Birds 40: 1169-1173. [ Links ]
Thomas, D., R.P. de Andrade, W. Couto, C.P. Moore & C.M.C. de Rocha. 1983. Pasture development in the tropical savanna region of Brazil. World Rev. An. Prod. 19: 37-44. [ Links ]
Toledo, V.M. 1988. La diversidad biológica de México. Ciencia y Desarrollo 8: 7-16. [ Links ]
Toledo, V.M. 1992. Bio-economic cost, pp. 63-71. In T. Downing, S. Hecht & H. Pearson (eds.). Development or destruction? The Conversion of tropical lorest to pasture in Latin America. Westview, New York. [ Links ]
Uhl, C., R.B. Buschbacher & E.A.S. Serráo. 1988. Abandoned pastures in castern Amazonia. I. Patterns of plant succession. J. Ecol. 76: 663-681. [ Links ]
Vandermeer, J. & I. Perfecto. 1995. Breakfast of biodiversity: The truth about rain forest destruction. The Institute for Food and Development Policy, Oakland, California. [ Links ]
Vaughan, C., M. McCoy, J. Fallas, H. Chaves, G. Barboza, G. Wong, M. Carbonell, J. Rau & M. Carranza. 1995, Plan de manejo y desarrollo del Parque Nacional Palo Verde y Reserva Biológica Lomas Barbudal. Contrato SENARA-BID-MIRENEM-UNA. Universidad Nacional, Heredia, Costa Rica. [ Links ]
Webb, R.H. & S.S. Stielstra. 1979. Sheep grazing effects on Mojave Desert vegetation and soils. Environmental Manage. 3: 517-529. [ Links ]
Winegar, H.H. 1977. Camp Creek channel fencing-plant, wildlife, soil and water responsos. Rangeman's J. 4: 10-12 [ Links ]
Zahawi, R.A. & C.K. Augspurger. 1999. Early plant succession in abandoned pastures in Ecuador. Biotropica 31: 540-552. [ Links ]
1 . Institute of Economic Botany, T'he New York Botanical Garden, Bronx, New York, 10458-5126, U.S.A.
2 . Instituto de Ecología, Departamento de Ecología de los Recursos Naturales, Universidad Nacional Autónoma de México, Apartado Postal 27-3 (Xangari) Morelia, Michoacán México 58089. Fax: 52 (443) 322-2712. mquesada@oikos.unam.mx ; kstoner@oikos.unam.mx

