










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.50 n.2 San José Jun. 2002
by
Jay M. Savage*
(Received por publication November 25, 1965)
Toads of the génus Bufo are not noted for their brilliant coloration nor marked sexual dichromism. Most members of the genus are dull creatures essentially gray or brown in ground color with dark blotches and sometimes light markings. Within the group a number of species show slight sexual dichromism, but relatively few approach the condition in such forms as the Asian Bufo raddei Strauch or Bufo canorus Camp of the high Sierra Nevada of California. Even in these latter species the colors are subdued greens, olives and dark to rusty browns.
In striking contrast to the usual Bufo situation, the new Costa Rican toad described below exhibits the most startling coloration and development of sexual dichromism recorded in the genus. The first individuals collected were a small series of males, all uniform bright orange in color. I must confess that my initial response when I saw them was one of disbelief and suspicion that someone had dipped the examples in enamel paint. The females proved to be equally astonishing, for they are olive to black with a series of large spots of brightest scarlet and without any hint of orange in their coloration. The new form is assuredly the most spectacularly colored Bufo known and is among the gaudiest of anurans. With reference to the extraordinary coloration the new toad will be called.
Bufo periglenes new species
HOLOTYPE: LA 1893; an adult male, collected May 14, 1964, by Jerry James, Norman J. Scott, Jr., and Jay M. Savage.
TYPE LOCALITY: COSTA RICA: Provincia de Alajuela: Cantón de San Carlos: Cordillera de Tilarán, 2 miles ENE of Monteverde, Provincia de Puntarenas; 1590 meters.
PARATYPES: 142 males and 31 females from the type locality, taken in April, 1964, and May 12, 14, 15, 1964.
DIAGNOSIS: Inmediately distinguished from all other recognized species in the genus by the coloration, bright uniform orange (yellow in preservative) in males and olive to black, with large scarlet spots in females. In addition, the new toad differs from other New World Bufo in the following combination of characteristics: 1) external cranial crests: low canthal, supraorbital and postorbital and knob-like supratympanic; 2) parotoid gland moderate, equal in length to upper eyelid, spinose; 3) no tympanum; 4) small ostia pharyngea present; 5) no vocal sac or slits in males; 6) no gready enlarged warts in a series along sides; 7) first finger equaI in length to second; 8) no well-developed subarticular tubercles on hands and feet, tubercles on palms and soles obsolete; 9) no tarsal fold or tubercles; 10) fingers not webbed, toes webbed at base.
In Middle America only species that usually (B. occidentalis of México) or always (B. bocourti and B. tacamensis of Guatemala and 'Southern Mexico; B. coerulescens, B. fastidiosus and B. holdridgei of Costa Rica; and B. simus and B. véraguensis of Panamá) lack a tympanum might be confused with periglenes. The new form may be distinguished from occidentalis, bocourti and tacanensis by having a well-developed knoblike supratympanic crest that separates the parotoid gland widely from the upper eyelid (versus no supratympanic crest and moderate to large paratoid glands that meet or are narrowly separated from the upper eyelid in the other three forms).
Bufo periglenes differs from veraguensis in having webbing only at the bases of the toes, a moderate parotoid gland equal in length to the upper eyelid and no parietal crest (versus toes aImost fuIly webbed, a large parotoid gland equal in length to the head length and a parietal crest in the Panamá species).
Bufo coerulescens, B. fastidiosus, B. holdridgeiand B. simus lack ostia pharyngea and have a series of enlarged warts along the sides of the body (ostia present and no enlarged warts forming a lateral series in periglenes).
Externally the new species differs from the lower Central American bufonid, Crepiáophryne epioticus, in the same characteristics that separate periglenes from the coerulescens.fastidiosus-holdridgei-simus series.
GENERAL CHARACTERISTICS: A relatively smooth toad with granular warts on upper surfaces; warts tipped with small black pointed spines. Head, viewed from above, wedge-shaped, about as long as broad. Snout pointed in males, subelliptical in females. Upper surfaces of head with a very few granular warts
in males, rugose in females. Upper eyelids warty in both sexes. Interorbital width about equal to one-fourth head width, three-fourths length of upper eyelid. From side, snout weakly acute in males, rounded in females. Nostrils lateral. Snout longer in males than in females. Canthus rostralis obtuse in cross-section, loreal region weakly concave. Orbit longer than high, equal to snout length. Eyelid membranes opaque but unpigmented. Pupil horizontally elliptical. Side of head warty.
All cranial crests relatively low, marked by smooth warts; best developed in females. Canthal, supraorbital and postorbital crest form a smooth continuous ridge from nostril to swollen enlarged supratympanic. Supratympanic from eyelid to parotoid gland. No suggestion of crests on side of head.
Parotoid glands, elongate but inconspicuous; width about three-fifths of length; warty. Dorsum covered with heterogeneous mixture of large and small warts; fewer large warts in males than in females; warts largest on sides and upper surfaces of limbs.
Anterior limbs relatively long, longer in males than in females, more than half of standard length. Third finger longest. In males upper surface of first and second finger and posterior margin of first covered by dark horny material. Undersides of palm and fingers covered by very low obscure rounded tubercles. Thenar and palmar tubercles rounded, large; palmar largest; thenar in males partially covered by dark horny material continuous with similar material on upper surface of hand. Hindlimbs relatively long, longer in males than in females. Toes webbed at base, with lateral fleshy fringes; only about one-half of each proximal phalanx webbed. Underside of foot with poorly developed, obscure subarticular and plantar tubercles. Metatarsal tubercles rounded, inner elongate, outer smallest.
Throat granular. Ventral surfaces covered with small warts, somewhat smaller, but otherwise indistinguishable from dorsal warts.
Choanae ovoid, large, near level of anterior margin of orbit; separated by a distance twice that separating nostrils. No teeth or odontoids. Ostia pharyngea very small slits. Tongue pyriform, not notched.
COLORATION: In life. Males (Fig. 1): a uniform bright yellow-orange, orange or orange-red; venter somewhat lighter than dorsum and obscurely mottled with darker and lighter tones of orange and in some individuals a few faint dark or flesh-colored spots. Supraorbital crests, nuptial pads on fingers and wart spicules dark brown to black. Females (Fig. 2): dorsal ground color of head, body, and limbs dirty greenish yellow, yellow-olive, smoky black or dark black. In lighter colored individuals a number of dusky black markings, blotches or large spots; in some examples dark markings form irregular lines on paravertebral region to give impression of striped pattern with light vertebral region. All examples with bright scarlet spots on upper surfaces. Red spots outlined by yellow to yellow-olive, in some individuals a yellowish ring outlined by black. Throat dirty yellow to flesh. Venter and undersides of limbs greenish yellow to flesh, frequently with black mottling and irregular pink to red blotches. Undersides of hindimbs similar to venter or same color as upper surface of limbs.
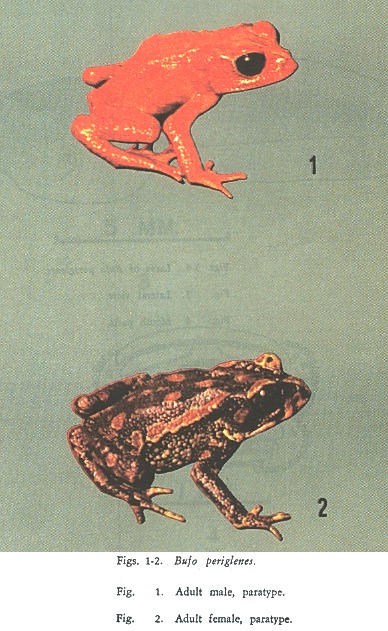
Iris in both sexes black with a few golden flecks.
A recently transformed toadlet deep brown with pale bluish-white spots; venter mottled with black and pale bluish-white.
In preservative males uniform faded yellow. Females similar to life colors although generally duller and colors somewhat faded.
MEASUREMENTS: Standard length (snout to vent) is in millimeters all other measurements are given as percentages of standard length. In the style of notation used, the first figure indicates the lower range limit, the second figure the mean and the last figure the maximum. Data are based on 10 individuals of each sex. A recently transformed example, collected at the type locality, is 18 mm in standard length.
Males: standard length, 41. - 44.9 - 48; head length, 32 - 34.3 . 37; head width, 30 - 33.0 - 35; orbit, 10 - 11.2 - 12; snout length (eye to tip of snout), 14 - 14.6 - 16; interorbital width, 7.5 - 9.1 - 10; length of upper eyelid, 9.1 - 10; length of arm, 55 - 65.8 - 89; leg length (anus to tip of longest toe), 132 - 138.3 - 148; length of tibia, 33 - 40.0 - 44; length of foot (including tarsus), 59 - 65.5 - 74; length of 4th toe (from base of 3rd), 26 - 28.1 - 31.
Females; standard length, 47 - 51.4 - 54; head length, 34 - 34.9 - 37; head width, 33 - 35.2 - 37; orbit, 9.5 - 11.4 - 13; snout length, 11 - 11.6 - 16; interorbital width, 8.7 - 9.9 - 11; length of upper eyelid, 12 - 12.9 - 15; parotoid length, 11 - 13.4 - 17; parotoid width, 6.7 - 7.8 - 8.3; length of arm, 50 - 57.3 - 63; leg length, 111 - 123.7 - 134; length of tibia, 34 - 37.3 - 40; length of foot, 52 - 57.9 - 62; length of 4th toe, 22 - 26.2 - 30.
Sexual dimorphism is indicated for the following measurements: snout length, paratoid width, arm and hidlimb lengths. In all these instances the measurements in males are proportionately larger.
LARVAE: A series of eggs collected from an amplexing pair from the type locality were raised. The eggs were laid on May 14, 1964, and the first larvae transformed on June 21, 1964. Five advanced larvae at stages 35-36 (LIMBAUGH and VOLPE, 6) form the basis for this description (Fig. 3).
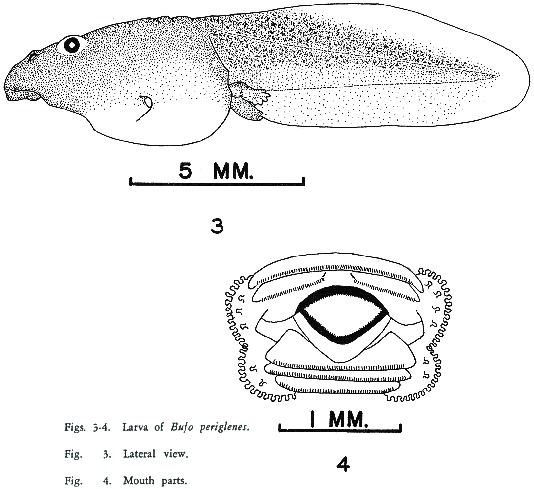
Body relatively short and shallow, somewhat depressed; greatest depth about one-half of standard length. Eyes and nostrils dorsal; eyes directed laterally, nostrils dorsolaterally. Anus median, spiracle sinistral. Tail fins moderate, rounded at tip.
Mouth (Fig. 4) on underside of head, near tip. Labial disk small; labia infolded laterally; a single row of papillae on lateral margins of disk, none on upper (anterior) or lower (posterior) margins. Denticles in 2/3 row; rows on lower lip slightly shorter than others; uppermost row not complete, with a gap over mouth. Beaks small, upper weak.
Coloration a uniform deep dark brown, with no distinctive pattern. Ventral areas lighter than dorsum.
Standrad length 6-7.08 mm; smallest transformed toadlets 7.0 mm.
Ecological Notes
In 1963 Norman J. Scott, Jr., and I visited the Costa Rica Quaker colony at Monteverde from October 16-22. The existence of Bufo periglenes was called to our attention by Mr. Jerry James, who reported that he had seen individuals of a bright solid orange toad on a trail near the continental divide that forms the boundary between Puntarenas and Alajuela provinces. He further noted that he had seen the toads only near the beginning of the rainy season in April and May. He had just returned from the area a few days before but said that none of the toads were active. Some months later Mr. James brought us several recently preserved specimens of the orange toad at the Universidad de Costa Rica. After seeing these examples, Scott and I, accompanied by Drs. Charles A. McLaughlin and Fred S. Truxal of the Los Angeles County Museum, left as soon as possible for Monteverde.
On May 14, 1964, James, Scott and I visited the area where the orange toads had been seen, about 2 miles ENE of Monteverde, Provincia de Puntarenas. Monteverde is located on a mesa-like ridge on the Pacific slope of the Cordillera de Tilarán at 1410 m. The bioclimate at this locality has produced a Subtropical Wet Forest, following the system of HOLDRIDGE (5), and excellent stands of undisturbed forest are present near the Quaker colony. At slightly higher elevations, around 1500 m, the forests receive more rain than at Monteverde and appear typical of Subtropical Rainforest, probably as the result of Atlantic rains that pass over the relatively low continental divide at 1600 m.
To reach the type locality of the new Bufo, we walked along a muddy meandering trail through the rainforest zone. After a hike of an hour and a half we came to another trail along the continental divide at an elevation of 1590-1600 m. Vegetation in the arca is characteristic of Lower Montane Rainforest, which develops in regions with a mean anual biotemperature between 12º and 18º C and an annual rainfall over 4000 mm. Because of the constant prevailing winds from the Caribbean that blow across the divide from the northeast, the trees on the ridge are somewhat stunted and the forest is almost always shrouded in clouds or fog and receives heavy rains almost daily. Under the forest canopy of broad-leaved evergreen trees the limbs and trunks are dark in color and covered with a rich, development of epiphytes. The soil is black and covered with fallen leaves in various stages of decomposition. The general impression is of a deep, dark, dank, dripping woods, rather cool in temperature. During the day the enclosed forest has an eerie atmosphere created by the black soil, trunks, and tangled roots and the billowing clouds of fog pass over head through the trees. At night the interior of the forest is pitch black.
Several toads were collected along the edge of the trail bordering the forest. When we stepped into the forest, everywhere we looked we saw bright blotches of orange standing out in marked contrast to the black forest-floor. Each orange blotch was a male Bufo periglenes. Within a radius of 5 meters at least 200 toads were visible. The females were much less common and less obvious than the males. A number of amplexing pairs were captured as the female carried the male about the forest. When approached the toads attempted to escape by crawling into holes among the roots of the trees or by hopping rapidly away. Several examples were observed above the ground (up to 0.5 m) on tree trunks or roots. All specimens collected were apparently actively foraging for food or involved in courtship. Collections were made between 3:00 and 5:00 P.M.
The species is apparently voiceless and no sound was heard at the type locality that could be attributed to these toads. Males do not emit a release call when grasped by other males or when artificially stimulated, but give a typical release vibration, (BOGERT 1). Several amplexing pair were congregated around a small water-filled depression, about 0.3 - 0.2 m wide and 5 cm deep.
We returned to the type locality that night (8:00-10:00 P. M.) in a driving rain. Fewer frogs were active than during the day but several were observed feeding on insects. Eggs had been deposited in the previously described depres. sion and 10 or 12 amplexing pairs were around the margins or in the water. One female subsequently laid 220 heavily pigmented small eggs in two long strings. Counts on ripe eggs for several females indicate that about 200 eggs are usually laid. A few additional specimens were obtained on the morning of May 15, 1964, between 8:00 and 9:00 A.M. Numerous examples were again active in the woods.
Other amphibians taken in association with the new species include: diurnal, Eleutherodactylus podiciferus; nocturnal, Atelopus varius, Hyla angustilineata, Hyla pseudopuma, Eleutherodactylus cruentus, Eleutherodactylus diastema, Eleutherodactylus palmatus, and Eleutherodactylus platyrhynchus.
The species is known only from a single locality in the Lower Montane Rainforest near the upper limits of the Subtropical Rainforest zone. These toads are apparently active diurnally and to some extent at night. The males are voiceless and the life history pattern is substantially similar to that of other Bufo.
The role of sexual dichromism in B. periglenes seems to be correlated with the breeding habits of the species. The absence of the auditory apparatus and voice in the new form indicates that location and recognition of potential mates may depend upon visual cues. In most anurans females locate males by the latter's call, but in this diurnal toad the bright orange coloration of the male probably serves the same function. Discrimination between males and females by the female is provided, in this instance, not by auditory but visual cues. The striking differences in coloration in the two sexes may thus serve an extremely important function in mate recognition. These statements are equally applicable whether or not the toads possess color vision, since differences in light intensities in the color pattern would provide an effective means of discrimination. Since males of all anurans have very poor abilities at sexual discrimination, the warning vibration in periglenes prevents amplexus between males. The extreme development of bright coloration in this toad suggests that the selective advantage of sexual recognition by the females far outweighs any disadvantage. Probably most potential predators on this ,form, principally small mammals, lack color vision and the aposematic value of the coloration lies at the intraspecific rather than inter-specific level. It may also be possible that the bright color functions to some extent as a warning device since most Bufo are distasteful or poisonous to some predators.
Relationships
The new species is referred to the genus Bufo on the basis of the following characteristics: tensor fasciae latae muscle not elongate; adductor longus muscle present; adductores mandibulae externus superficialis and posteriori subexternus present; eight procoelous presacral vertebrae; cervical and second vertebrae not fused; sacrum and coccyx not fused; sacral diapophyses expanded; coccyx biconcave and not expanded; quadratojugal meets maxillary; sphenethmoid ossified, single; palatines contact both maxillary and sphenethmoid, no omosternum; no teeth; phalangeal formula, hand 2-2-3-3, foot 2-2-3-4-3; a well-developed parotoid gland; eggs small, numerous and pigmented; larvae with median anus, sinistral spiracle, labia laterally infolded, labial papillae along lateral margins only, and denticles in 2/3 rows.
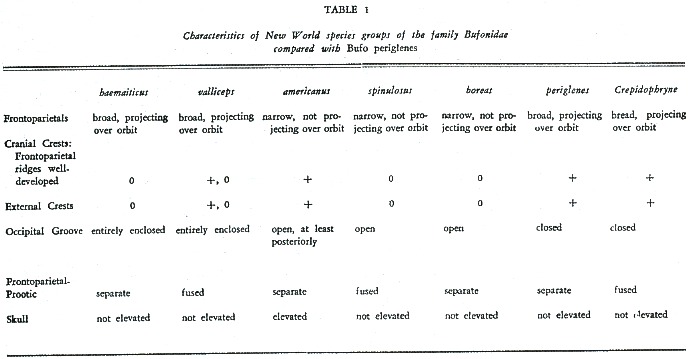
TIHEN (8) characterized five major New World groups within the genus Bufo. The new species differs from all of these groups in one or more diagnostic features (Table 1). Bufo periglenes (Figs. 5-7) has the frontoparietal broad and projecting over the orbit posteriorly, with the lateral margins upturned into smooth bony ridges, without dermal ornamentation and the skin over ridges forming external cranial crests; the groove for the occipital artery is a completely enclosed canal; the frontoparietals and prootics are separate; and the skull is not elevated. On the basis of these characters periglenes appears to be most closely allied with Bufo haematiticus Cope among American species. The new species differs most obviously from B. haematiticus in having the lateral margin of the frontoparietals upturned into a high smooth bony ridge, without dermal ornamentation and the skin over these ridges thickened and keratinized to form external crests. In haematiticus the lateral margin of the frontoparietals is slightly upturned and no external crests are present. In addition, direct comparison of periglenes and haematitius indicates other major differences (features of the latter species in parentheses): tympanic portion of squamosal enlarged projecting ventrally as a rectangular plate, for half length of squamosal shaft covered with extensive derma! ornamentation (tympanic portion of squamosal moderate, not projecting ventrally as a rectangular plate, not covered with extensive dermal ornamentation); supratympanic ridge enlarged, knob-like, covered by dermal ornamentation (no obvious supratympanic ridge); no columella (columella present); tibiofibula robust (tibiofibula slender and enlongate). In other features of the skeleton the two species are very similar. Externally they differ in numerous characteristics, the most notable being: cranial crests (none); moderate, elongate parotoid, about size of upper eyelid (Iarge parotoid, equaI in size to side of head); no tympanum (tympanum present); no vocal sacs or slits in males (a single vocal sac and one sinistraI vocal slit); and no tarsaI fold (an inner tarsal fold). Although the two species show considerable affinity in interna! structures they differ so markedly that any relationship must be a distant one. Nevertheless it seems appropriate to place periglenes tentatively in the haematiticus group while admitting the differences between the two included species.
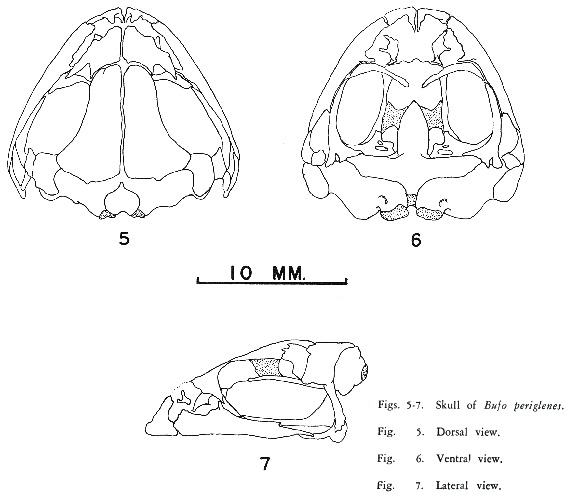
Superficially Bufo periglenes resembles the peculiar Costa Rican endemic genus Crepidophryne. This genus, erroneously called. Crepiditis Cope, 1875 (3), by SAVAGE and KLUGE (7), a name preoceupied by Crepiditis Candeze, 1859 (2), in Coleoptera, was given the available substitute name of Crepidophryne COPE (4). The single known species Crepidophryne epioticus (Cope, 1875) differs from periglenes by having a markedly expanded coccyx, the frontoparietal and prootic fused, a phalangeal formula of 1-2-3-2 for the hand and 1-2-2- 4-2 for the foot; and in lacking adductor longus and adductor mandibulae posteriori subexternus muscles. Crepidopbryne and the new toad are similar to one another in external cranial crest arrangement, the knoblike supratympanic crest, the absence of a tympanum and the obsolescent tubercles on hands and feet. In Crepidophryne the bony cranial crests of the frontoparietal are greatly enlarged and heavily ornamented and the entire auditory apparatus has been lost, while in B. periglenes the frontoparietal crests are without dermal ornamentation and a reduced Eustachian tube and small ostia pharyngea are present. The skeleton of Crepidophryne is much more reminiscent of the valliceps group than of B. periglenes or B. haematiticus and the resemblances between periglenes and C. epioticus are probably convergent. The relationship of Crepidopbryne to Bufo and the status of other Costa Rican toads without a tympanum (B. simus O. Schmidt, B. fastidiosus Cope, B. coerulescens Cope and B. holdridgei Taylor) will be covered in detail in a paper I have preparation.
The name of the new toad is from the Greek periglnzes, meaning very bright.
Acknowledgments
Thanks are extended to Mr. Jerry James of Monteverde, the original discoverer of this toad, and Mr. Norman J. Scott, Jr. of the Universidad de Costa Rica for field assistance. Scott also raised the tadpoles of the new form and provided the color plates (Figs. 1-2). The drawings of the tadpoles and skull were prepared by Anthony J. Gaudin.
The holotype and a series of paratypes will be deposited in the herpetological collections of the Los Angeles County Museum (LA).
Summary
A new species of toad, Bufo periglenes, is described from the Lower Montane Rainforest zone of the Cordillera de Tilarán on the divide between Puntarenas and AlajueIa provinces, Costa Rica. The new form exhibits a combination of extremely bright coloration and marked sexual dichromism. Males are solid orange, females greenish to black with scarlet spots. The species lacks a tympanum and columella and is voiceless. Tadpoles are described. Relationships and the role of sexual dichromism are discussed.
Resumen
Se describe una nueva especie de sapo, Bufo periglenes, del bosque montano bajo pluvial de la Cordillera de Tilarán en la divisoria entre las provincias de Puntarenas y Alajuela, Costa Rica. La nueva especie presenta una evidente combinación de colores brillantes y marcado dicromatismo sexual. Los machos son de un color anaranjado intenso uniforme, las hembras verdusco a negro con manchas rojo escarlata. Esta especie carece de tímpano, columela y voz. Se describen los renacuajos. Se discute el parentesco y el papel del dicromatismo sexual.
Literature Cited
1. BOGERT, C. M. 1960. The influence of sound on the behavior of amphibians and reptiles. Am. Inst. Biol. Sci., Publ. 7: 137-320. [ Links ]
2. CANDEZE, E. 1859. Monographie des Elatérides. Tome II. Mem. Soc. Sci. Liège, 14: 1-543. [ Links ]
3. COPE, E. D. 1875 On the Batrachia and Reptilia of Costa Rica. J. Acad Nat. Sci. Philadelphia, ser. 2, 8(4): 93-154. [ Links ]
4. COPE, E. D. 1889. The Batrachia of North America. Bull. U.S. Nat. Mus., 34: 1-525, pis. 1.85. [ Links ]
5. HOLDRIDGE, L. R. 1964. Life zone ecology. San José, Costa Rica. 117 pp., pis. 1-50. [ Links ]
6. UMBAUGH, B. A. 4t E. P. VOLPE 1957. Barly development of the gulf coast toad, Bufo valliceps Weigmann. Ame,. Mus. Nat. Hist. Nov., 1842: 1-32. [ Links ]
7. SAVAGE, J. M. 4t A. G. KLUGE 1961. Rediscovery of the strange Costa Rica toad, Crepidius epioticus Cope. Rev. Biol. Trop., 9(1): 39-51. [ Links ]
8. TIHEN, J. A. 1962. Osteological observations on New World Bufo. Am. Midi. Nat. 67 (1): 151-183. [ Links ]
* Department of Biological Sciences and AlIan Hancock Foundation, University of Southern California, Los Angeles, California, U.S.A.

