










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.50 n.2 San José Jun. 2002
insect orders
by
Alvaro Wille*
(Received for publication May 9, 1960)
The phylogeny and the interrelationships of the insect orders always remain a matter of great interest to the general entomologist. The number of papers that have been published on this subject is very large. Very unfortunately, however, most of these papers are too brief, containing only a few facts to support the views which the author has adopted, or they deal only with one order or a group of more or less interrelated orders. The purpose of this paper is to offer a general account of the origin of the insects and the known relationships between the insect orders, giving as many facts as possible to show these interrelationships.
As might be expected, current interpretations of the phylogeny of, and connections between the insect orders are not definitive, and many of our present views may have to be changed in the future, as knowledge progresses. Although this type of studies always reveals many unsolved problems, the large amount of data accumulated in the last few decades allows us to speculate on these matters, and in doing so, we may put some order and understanding into these chaotic accumulations of facts. This may explain, perhaps, the large number of papers on this subject, and will always justify further additions.
Since this is not a detailed revision of all the known views available in the literature, I have adopted those which the facts best seem to support. However, in those cases in which the data can be equally interpreted in different ways, the various interpretations are discussed.
Acknowledgments
The present work was started as a seminar, given a few years ago at the University of Kansas and the final form was completed at the University of Costa Rica, Facultad de Agronomía.
I wish to thank Dr. C. D. Michener, Dr. G. Byers, and Dr. R. Matsuda, for reading the manuscript and offering some of the University of Kansas, suggestions.
Thanks are due also to Dr. H. V. Daly, of Louisiana State University for checking many of the bibliographical citations and to Dr. R. L. Rodríguez of the University of Costa Rica, for his special interest in helping to draw the phylogenetic tree and proofreading the manuscript.
Finally, I wish to thank my wife, Mrs. María Eugenia B. de WiIle, for typing and proofreading much of this paper.
The Origin of the Insects *
The insects arose about 350 million years ago, probably in the lower Devonian or earlier. Our knowledge of the morphology of arthropods seems to indicate that the closest relatives of insects are the myriapods, of which the Symphyla have the greater number of characters in common with the primitive insects. This does not mean that the insects arose directly from the Symphyla or other myriapod classes, but rather that the insects and myriapods were derived from a common stock or from myriapod-like arthropods. It is also possible that insects originated by neoteny from myriapod-like arthropods, at least there are some evidences for this. Evolution by neoteny is probably much more common in the animal kingdom than most biologists have supposed. There are two main types of neoteny, one the which the rate of development of the body remains more of less constant but the reproductive system is accelerated, and the other in which the rate of development of the body is retarded but there is no acceleration of the rate of development of the reproductive organs. The first type of neoteny leads to specialization, usually resulting in a simplification often associated with parasitism, and is of little importance for phylogeny. This happens, as DE BEER (1) has pointed out, because "if the reproductive system is accelerated, the structure of the body will be less fully developed when the animal is sexually mature than was the body of the ancestor. On the other hand, if there is no acceleration 'of the rate of development of the reproductive system, but only a slower rate of succession of ontogenetic stages of the body, the latter will be no less ' well made' for retaining the larval plan." We know that evolution by neoteny probably took place in a number of cases. Of these we can mention the chordates from the echinoderms, and the appendicularians from the ascidians. It is possible that even man evolved neotenically from former anthropoid-like primates (for details see DE BEER, 1). Neotenic evolution may explain the lack of the so-called missing links in some cases, and affords a means of emancipation, from the restraints of extreme specialization.
The evidence that the insects may have evolved from myriapod-like arthropods by neoteny, is based on the fact that the first instar of many myriapods, such as Strongylosoma and Iulus in the DipIopoda, show a great number of similarities with the insects, namely, a head of six united segments, a thorax composed of three segments, each with a pair of legs, and an abdomen of about five segments without appendages, or with only very vestigial legs. If a form like this becomes neotenic, undergoing a retardation in the development of the legs behind the first three pairs, * and the larval number of body segments retained into the adult stage, it wouId be very insect-like and could give rise to the Apterygota. It is interesting to point out that among the Apterygota, there are many species which have leg-like appendages on the abdominal segments. Furthermore, there is a large group, the Collembola, in which the abdomen is composed of six segments only.
Evidence that the myriapods are most closely related to the ancestors of the insects is indicated by the similarity of several characters found in common in thc myriapods and insects. These characters are the following:
1) Absence of the dorsal muscle (levator) of the pretarsus in all myriapods and insects.
2) With one pair of antennae. Moreover, antennae composed of a variable number of true segments each being supplied by one or more muscles, is a common feature in all the myriapods and in some primitive insects (Diplura and Collembola).
3) Retention of at least two segmental organs in the head, sometimes only partially retained, is common in some myriapods and primitive insects, in which they function as salivary and nephrocytic organs.
4) The problematic organ of Tömösvary (postantennal organ) is common to Diplopoda, Symphyla and Collembola.
5) Eversible sacs at the bases of the appendages are common to all Symphyla, some Diplopoda, Pauropoda, Diplura, and several lepismid genera.
6) The similarity of the Malpighian and tracheal tubes in both groups.
7) The similarity in structure and development of the heart and aorta. Segmental blood vessels occur in Diplopoda, Chilopoda, and even in the orthopteroid insects.
8) A massive fat body in the haemocoele is common in both groups.
9) Ecdysis takes place through a transverse split at the hind border of the head in myriapods, Protura and Collembola.
As has been already mentioned, among the myriapods, the Symphyla seem the closest to the insects, especially to the primitive forms. This is suggested by the following characters, common to SymphyIa and insects:
1) As shaped epicraneal suture or ecdysial suture is common to many Symphyla and insects. However, it should be pointed out, that since the arms (i)f the epicraneal suture vary in the different groups of insects, being merely the lines along which the cuticle splits in molting, and since other sutures have been confused with it, its morphological value is not as great as was once thought.
2) The structure of the postmandibular appendages of the head is essentially the same in Symphyla and Insecta. The first postmandibular appendages are paired maxillae, each is composed of stipes and cardo, the stipes having a pair of apical lobes, the galea and lacinia. This type of maxilla is found only in Symphyla and Insecta. The second postmandibular appendages are united, forming a labium.
3) A distinetly three-lobed hypopharynx, consisting of a median lingua and a pair of anterolateral superlinguae, is found in the Symphyla and some primitive insects, such as Diplura, Collembola, Machilidae, and the larvae of Ephemeroptera (according to George Byers, in a personal communication, some cranefly larvae have also a three-lobed hypopharynx).
4) The head apodemes of the Symphyla are, according to SNODGRASS- (35), "suggestive of the anterior arms of the tentorium of Thysanura and pterygote insects, inasmuch as they give attachment to the same muscles that are attached to the insect tentorium."
5) An abdomen with styles and eversible sacs is a common feature of the Symphyla and Diplura.
6) The terminal cerci of Symphyla seem to correspond to those of insects. Furthermore, the cerci of some Symphyla (Scolopendrella) and Diplura (Anajapyx) are provided with similar spinning glands.
7) A premandibular segmentaI .organ is evident in the late embryo of Symphyla, and in some species it survives in the adult. Evidence of the premandibular organ in the insects was first found in the orthopteran embryos and was called "sub-oesophageal body." It is also found in Plecoptera, Isoptera, MaHophoga, Coleopteta and Lepidoptera.
8) The peculiar 'dorsal organ' of the embryo of Symphyla is al so found in Collembola and Campodea and even in higher insects (see JOHANNSEN and BUTT, 20).
9) Embryologically, TIEG, 37, 38) has shown that the Symphyla have fourteen trunk segments, as in insects.
10) To these we may add that, according to MANTON (23), "the full range of insectan gaits (i.e. relative duration of forward and backward strokes and phase differences between successive legs) is seen in the Symphyla, bur nor in the adult of any other group of myriapods."
The theory that the Symphyla stand closest to the ancestor of the insects is fairly well supported by the above data. Apparently, however, it presents a few weaknesses. According to SNODGRASS (35), those who believe in the Symphyla as the ancestral relatives of the insects "give too little attention to the discrepancy in the mandibular structure." In spite of this TIEGS and MANTON (39) made the following statement, "the mandibles, though segmented, could be the forerunner of the thysanuroid mandible." Another major objection is the progoneate condition of the Symphyla, that is to say, the reproductive organs of these myriapods open upon the third somite behind the head. To this, TIEGS and MANTON (39) replied: "A suggestion that comes to mind is that some myriapods have found it expedient to evolve a new genital opening (one might point, for analogy, to the accessory male genitalia in Odonata) when the terminal segments became involved in anamorphosis and many Diplopoda even show sexual precocity."
Coming back again to the idea of evolution by neoteny, it should be recalled that insects could not have been derived from adult Symphyla, for the structure of the latter is too specialized. In the same way, neoteny could not have occurred with the type of development which the Symphyla have at the present time, and give origin to the insects. This is due to the fact that the young in Symphyla hatch with six or seven pairs of legs. Therefore, if we assume that the insects originated from the myriapods by neoteny, and. at the same time believe in the Symphyla as the ancestral relatives of the insects, we must have to presume that the present development of the Symphyla is secondary, and that formerly the young hatched with three pairs of legs, just as is the case with Diplopoda and Pauropoda. Attention is called to the fact that the first moult of the hexapod larva of Diplopoda gives origin to a young with usually seven pairs of legs, which would correspond to the present stage of the young in Symphyla. This may suggest, perhaps, that the Symphyla have repressed the first stages only.
In summary, our present knowledge seems to indicate that the myriapods, especially the Symphyla, stand closest to the ancestors of the insects. Furthermore, the peculiar hexapod larvae of many myriapods may also suggest that the Insects could have originated by neoteny from myriapod-like arthropods.
Major Trends in the Evolution of Insects
Three main radiations seem to have taken place during the evolution of the insects. The second and third of these radiations were typically explosive evolution. In insects the first radiation is represented by the Apterygota, found from the Devonian to the Recent. This was the first step in the evolution of insects and therefore the one that had and still preserves the most primitive characters shared only with the myriapods. The second step in the evolution of insects was represented by the Paleoptera, presumably the first insects which acquired wings, but could not flex them on the back of the abdomen. This group is represented in the geological strata of the lower Pennsylvanian to Recent. The acquisition of wings made them a dominant and successful group. The Paleoptera seem to have undergone an explosive evolution which somewhat paralleled the Neoptera. Thus we find forms which were similar to Hemiptera (Protohemiptera*), others were like the Hymenoptera (Protohymenoptera), and still other forms acquired independently the capacity to flex the wings
(Megasecoptera), etc. The Paleoptera were very abundant during the Pennsylvanian, much more so than the Neoptera of that time; but after the Permian they diminished in number, displaced by the better adapted Neoptera and today this primitive group is represented only by two orders, the Ephemeroptera and Odonata. The third and last radiation of insects was the Neoptera, presumably derived from the Paleoptera. They were characterized by the capacity to flex the wings. This characteristic was obviously more advantageous than the inability to fold them. It permitted the insect to occupy more ecological niches which otherwise could not be occupied since paleopteroid wings were a hindrance far the insects in places such as water, underground, brushy areas, etc. The Neoptera are found in the geological strata from the lower Pennsylvanian to Recent, and have radiated into the numerous phyletic lines of our modern insects.
The Apterygota
It is the contention of some authors that the entognathous groups (Collembola, Protura, Diplura) should be excluded from the insects. This has been based on the following unusual features found in these groups;
Unusual Features of Protura
1) Lack of antennae.
2) The mandible is articulated to the cranium by a slender rod. as in Chilopoda. It should be pointed out, however. that this rod was not observed by TUXEN (43). He adds further that "perhaps it is the fanlike ligament taking this aspect in certain views."
3) The abdomen is composed of twelve segments. one segment too many.
4) The gonopores of both sexes are found on the eleventh abdominal segment. It should be recalled that in all insects, excepting Protura, Collembola, and Ephemeroptera, the gonopores are found on the eighth for the female and on the ninth for the male, although some females have the gonopores open on the ninth also. The gonopore in both sexes of Protura is surrounded by small external genitalia of an unusual form, comparable to that of no other group.
5) The Malpighian tubes are represented by six papillae.
6) The inmature instars have a myriapod-like deveIopment. They develop by anamorphosis, that is, when they hatch they have nine abdominal segments, the other segments are added during the postembryonic growth, which involves five larval stages.
Unusual Features of Collembola
1) Flagellar muscles in the antennae.
2) The mandible is also articulated to the cranium by a slender rod, as in Chilopoda. However, according to TUXEN (43), this is not a real rod but a ligament and therefore not homologous with the rod of Chilopoda.
3) The abdomen is composed of only six segments.
4) The gonopores of both sexes are found on the posterior margin of the fifth abdominal segments.
5) Malpighian tubes lacking.
6) Just behind the antennae of some Collembola (most Poduroidea and some Entomobryoidea) there is a peculiar structure known as the postantennal organ, possibly a sensory organ homologous to the organ of Tömösvary in Diplopoda and Symphyla.
7) Gonads with a lateral germarium; that is, unlike other insects, the germarium of the ovaries and testes is lateral and not apical in position.
8) The egg is holoblastic, as in Symphyla and some diplopods, that is, a total cleavage takes place in the embryonic development, instead of the usual meroblastic and centrolecithal deavages.
9) Lack of embryonic membranes, that is, the egg does not develop an amnion and serosa, as in the myriapods.
UNUSUAL FEATURES OF DIPLURA
1 ) FlageIlar muscles in the antennae.
2) Segmental ovarioles in some species.
3) Lack of embryonic membranes.
4) The Malpighian tubes are represented in some species by small papillae.
It should be pointed out, however, that the entognathous groups stand at least near the base of the phylogenetic tree of the more typical insects (Fig. 1). This would explain such unusual primitive features as the antennae with the flagellar segments provided with muscles; the presence of the postantennal organ; the mandible articulated to the cranium. by a slender rod; the abdomen composed of twelve segments, which agrees with the primitive number found in embryonic insects; malpighian tubes represented by papillae; anamorphosis; segmental ovarioles; total egg-cleavage; and even perhaps the lack of embryonic membranes, although this could be a secondary feature, as Is the case of some ants in which the amnion is wholly lacking and the serosais rudimentary, represented by only a few cells. The other unusual characters are mere specializations. Thus, the absence of antennae in Protura is probably a secondary feature; after all, these organs are reduced to minute papillae in many insect larvae. According to TUXEN (42), the pseudoculi are remnants of the antennae, a conclusion reached after investigating their structure, innervation and musculature. On the other hand, the entognathous group's have also typical insectan features. Their mouth-parts are insectan in character; their differentiated thorax and three pairs of legs are also insectan features, as well as the reduction of the abdominal appendages. To this we may add their ontogeny, which is in general insectan in character.
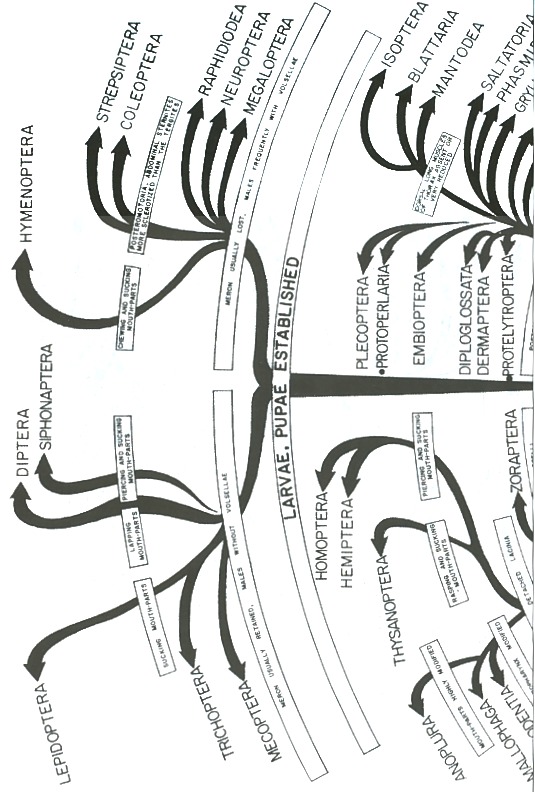
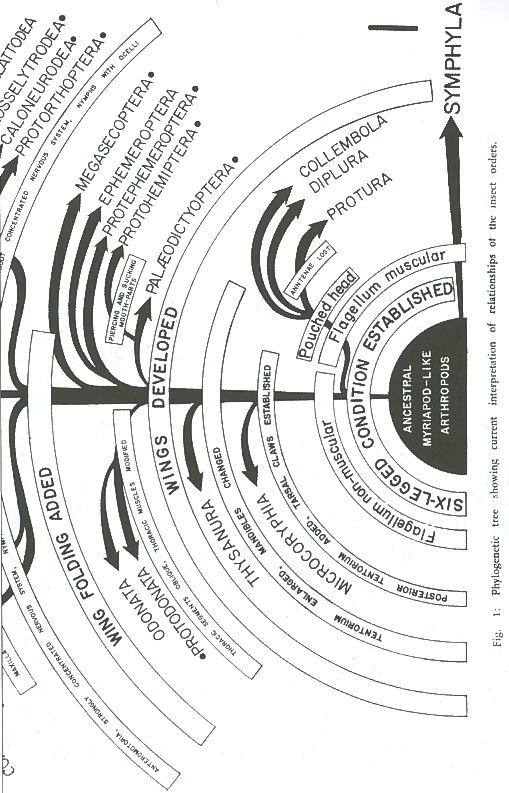
Those who believe that the entognathous group's should be excluded from the insects assume that the Apterygota are a polyphyletic assemblage of arthropods. However, it should. be pointed out that the entognathous apterygotes (Diplura, Protura, Collembola) appear to be related, as will be shown later on, to each other. Therefore, if polyphyly has in fact occurred in the Apterygota we should regard the Insecta as a diphyletic group. That this is actually the case we are not sure, but even if true, this would not constitute a satisfactory reason for restricting the definition of the class Insecta so as to 'exclude them. It has been argued that all the animals within a given group should have had a common origin, that is to say, to be monophyletic. However, we should remember that classification is, after all, a practical problem, and the idea that a group must be derived from a single species of a preceding group, cannot be always met in practice. To begin with, knowledge is far from complete and monophyly certainly cannot be demonstrated in any case. Furthermore, if theoretical monophyly is too strictly demanded, this may give as a result a classification in which the groups (with different names) cannot be distingllished morphologically. For this reason, SIMPSON (32) has suggested that a group, whatever its rank, should be composed of related animals that can be defined by morphological and related data, and have originated from a group of animals of lower rank than itself. That is to say, if the group in question corresponds to a class, it should have originated from a unified group smaller than a class, i. e. a superfamily, family, genus, etc. In closing this discussion, one should remember that the class Mammalia: is regarded by most paleontologists and mammalogist's as a polyphyletic group, and yet, most agree that is should be kept as a class.
It is obvious that the present apterygotes are too specialized to be direct deseendants from existing myriapod-like arthropods. There have be en several attempts to show which order is the most primitive. According to CRAMPTON (5), the Machilis-like Apterygota are morphologically the most primitive, but according to EWING (11), the Protura and Collembola "are to be regarded as groups of very ancient origin." On the other hand, IMMS (18), regards Diplura as the closest relatives of the Symphyla. This diversity of opinions shows very clearly how the primitive characters together with the specialized ones, are diversely distributed among these orders. In other words, the machiloid and the entognathous apterygotes should be considered as a whole as the most primitive insects, or closest to the ancestral group. Therefore, if the insects have any connection with the myriapods, this connection is not to be found in any one of the apterygote orders but rather in all these groups.
The Diplura, Protura, and Collembola seem to be related to each other, as suggested by SNODGRASS (35) and TUXEN (43). That these three orders are related is indicated by the following characters:
1) The mandibles and maxillae are enclosed in pouches of the head wall. This condition is called entognathy; it comes about through the formation of two folds (plicae orales), one on each side of the head, uniting at or below the sides of the labium. Entognathy is found in no other arthropod group.
2) The structure of the mouth-parts similar, as shown by TUXEN (43). Thus "the mandibles of the Diplura and Collembola are rather closely alike both in themselves and as regards their muscles, whereas the mandibles of Protura have a different musculature, are not hollow, and have no teeth. The mandibles of Protura are piercing organs necessitating especially strong protracting and retracting movements; and this, in connection with the probably secondary fact that they are not hollow, may account for the difference... The general plan of the maxilIa in the three groups is very much alike; especially the shape and position of the cardo are identical... In all three groups the lacinia carries an arm or platelike process to which one or more muscles are attached, running to the stipes, to the hind wall of the head, or to both. And in all three groups powerful muscles connect the stipes and the distal part of the cardo, as it seems, to the fulcrum."
3) Myriapod type antennae, that is, composed of true segments, each with one or more muscles.
4) The endoskeletal head structure is superficially very similar to the simple myriapodan tentorial armo According to SNODGRASS (35), however, this tentorium does not correspond to that of myriapods, Thysanura, and Pterygota, but rather to the postoral sternal brachia of Crustacea. FOLSON (12) and HANSEN (13) have shown that the supposed anterior tentorial arms of the entognathous apterygotes are superficial sclerotizations of the sternal wall of the head, rather than true apodemes as in the Symphyla and other myriapods. In Diplura the anterior parts of there brachia are connected inside the head by a ligamentous bridge which seems to correspond to that of Chilopoda. In Collembola this ligamentous bridge is elaborated into a complicated framework, which according to SNODGRASS (35) has not been demonstrated to be a cuticular structure and is similar to the endosternum of Arachnida. In Protura the anterior parts of the brachia are united, and no ligamentous bridge is found. It has also anterior connections with the cranium which may be secondary apodemal processes.
5) The maxillae are articulated with the posterior parts of the endoskeletal head structure. This, according to HANSEN (13), is a further proof that this type of tentorium is not homologous with that of other arthropods because "in no other arthropod are the maxillae articulated to any internal part of a tentorium."
6) True tarsomeres lacking.
Therefore, the Diplura, Protura, and Collembola, seem to have evolved at first in the same direction, but then they followed different lines (Fig. 1). It appears, then, that these three groups are better represented in a phylogenetic tree as on the same plane or level after splitting up from a common branch. The Protura and Collembola are more specialized in many ways than the Diplura, although they retain several primitive characters not found in Diplura. The Diplura is the most insect like of the three and perhaps the least specialized.
The remaining groups of Apterygota are the lepismatids and machilids, which are usually regarded as forming the order Thysanura. However, they are separated here in two orders or phyletic lines, as has already been done by CRAMPTON (6), HENNING (14) and Ross (29). There is no doubt that these two groups are similar in their general appearancce, but on the other hand, they differ structurally on several important features. These differences are summarized in table 1.

Machiloidea by Handlirsch, 1903 - as an order
Microcoryphia by Verhoeff, 1904 (April 22) - as an order
Archaeognatha by Borner, 1904 (May 3) - as a suborder
Since Machiloidea has been used as a superfamily name (REMINGTON, 27), Microcoryphia seems the proper name to use for the order. The old name Thysanura should be retained for the lepismatids.
Although the Microcoryphia show no specialization of the mouth-parts like the entognathous apterygotes, they are a little higher in relative advancement, as indicated by the following characters:
1) Development without any of the primitive features of the entognathous apterygotes, namely without anamorphosis, without total cleavage, without dorsal organs in the developing egg; and with embryonic membranes.
2) Flagellar segments not musculated.
3) Tentorium with anterior, posterior, and dorsal arms, comparable to those of lepismatids and pterygotes. However, it is not fused into a large central plate. It should be pointed out that the anterior arms of the tentorium in machilids and lepismatids have a ventral position on the head.
4) Genitalia comparable to that of other pterygotes.
5) True tarsomeres present.
Although the Microcoryphia stand a little higher in the trunk of the evolutionary tree, they are not too far from the entognathous apterygotes, as is shown by the few similarities with the Diplura, namely a three-lobed hypopharynx, cerci, styli, and eversible sacs. On the basis of these resemblances the Diplura have been placed among the Thysanura but this arrangement does not fit the information discussed above.
Of all the apterygotes, the Thysanura (s. str.) stand highest in the evolutionary tree, and therefore closest to the pterygote insects. That they are less primitive than the Microcoryphia has been shown already in Table 1.
In summary, the Apterygota are composed of five main groups of insects: Diplura, Protura, Collembola, Microcoryphia, and' Thysanura. The first three show a few features in common which may indicate certain interrelationships among them. Although the Diplura, Protura and Collembola are groups of very ancient origin, they should be regarded as insects judging by the many characteristics which they share with other apterygotes and with the more generalized pterygotes. The Microcoryphia and Thysanura are more insect-like, but they still retain primitive features and should be regarded as ancestral groups; they have styli on the thoracic legs; appendages en the abdominal segments; labial kidneys; a short germ band; an open amniotic cavity; an amnion developed by invagination; and a medial frontal organ with double cells and a disk-shaped structure, as in Crustacea. On the other hand, as MARCUS (24) has already indicated, they "also show characters in common with the Pterygota: mouth-parts; distinct paranota on all segments; antenna divided into shaft and flagellum and provided with Johnston's organ; a typical number and position of the stigmata; and an orthopteroid ovipositor formed of the eighth and ninth abdominal legs."
The Pterygota
The first major explosive evolution of the insects began with the development of wings. This ability to fly changed the direction of insect evolution. The actual derivation of the pterygotes from the apterygotes has been the subject of much speculation.
There are three main theories as to the origin of the wings. All these views agree in that the wings were developed from paranotal processes on the meso-and meta-thorax. This is based on the fact that many of the fossil insects (Palaeodictyoptera) show these processes on the prothorax and sometimes on the abdomen.
The classic view is that of MARTYNOV et al. (25). According to them the paranotal extensions tended to grow sidewards and were used as gliding organs. Since these processes had a selective value they were enlarged further. This happened only with the meso-and metathoracic extensions, probably by a series of mutations fixed by natural selection. The next step was the acquisition of a line of flexibility at the base of each process. In this way the wings originated. These wings were at first paddling wings, but later some acquired the ability to flex them on the back of the abdomen, thus giving origin to the neopterous insects. According to this view the insects are naturally divided into two groups: The Paleoptcra, including those insects incapable of flexing the wings (primitive condition), such as Odonata and Ephemeroptera, plus the fossil groups Palaeodictyoptera, Protohemiptera, Protodonata, Protoephcmeroptera, and Megasecoptera; and the Neoptera, including those insects capable of flexing the wings on the back of the body (secondary condition), such as the rest of the insects.
SCHWANWIl'SCH (31) points out that an insect with immobile gliding paranota would be hindered in its crawling through the vegetation. He believes that the paranota were directed backwards and became mobile from the very beginning of their evolution. In this way they could help walking by their strokes as in the case of Bombyx mori, which has lost its flight. Later they were trans. formed into true wings by elongation. Therefore SCHWANWITSCH'S view is the opposite of that of MARTYNOV, because he assumes that the ability to flex the wings on the back of the abdomen should be regarded as primitive and the spread out position as secondary. He divides the Pterygota into two main types, based on the wing motor musculature. The first type is found in Odonata, which have two systems of dorsoventral muscles for the up and down movements of the wings; this he named Orthomyaria. The other type is found in all the rest of insects in which the upward movement of the wings is accomplished by dorsoventral muscles as in Odonata, but the downward movement is achieved by longitudinal muscles, this he named Chiastomyaria (SCHWANWITSCH, 30).
The third view is somewhat intermediate to the two previous ones in that both conditions, flexed and unflexed, are considered primitive. LEMCHE (22) believes that the wings originated from two different types of growth of the paranotal process. In one the growth was sidewards giving rise to the Palaeodictyoptera, Protohemiptera, Protodonata, Odonata, and Megasecoptera. In the other, growth was backwards and gave origin to the rest of the orders. And these two lines of development were independent of each other. LEMCHE based this belief on the following observations.
1) The fossil record does not contradict the diphyletic conception of the Pterygota. Both groups, Paleoptera and Neoptera, are found at the same time from the lower Pennsylvanian onward.
2) All the known nymphs of the pads. On the other band, the pads directed backward.
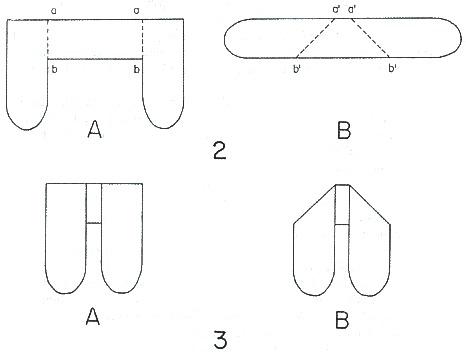
3) All present-day Pterygota develop the wing pads backwards at least in all the groups in which this growth can be observed, with the exception of the Saltatoria and Odonata. In the Saltatoria, however, the wing pads start growing backwards just like those of Blattaria, but in the next molt these wing pads are titled along axes parallelling the longitudinal axis of the body, thus showing a secondary condition. It is comparable to Blattaria. In the Odonata, the wings start growing as small vertical ridges at the sides of the meso-and metatborax. These ridges grow from lines forming an angle with the longitudinal axis of the body. At the same time the wings are tilted as in Saltatoria, therefore the growth of the wings in Odonata is essentially a sideward growth; on the other band, in the Saltatoria thc growth is backward. This is best illustrated by cutting two papers as shown in figure 2 and folding them along the lines marked a-b, a'-b'.
4) The tracheal ramifications in the wings that grow backwards show that the wing trachcae are bent toward the back from the very beginning, displaying the same relation to the hind comer of the paranotum as the veins in the full grown wings to the wing tip. The tracheal ramifications in the prothorax are quite similar. This seems to indicate that the wings have not changed secondarily to the backward growth. Thus the tracheae were originally directed backwards, parallelling the longitudinal direction of the wing or spreading - out in the shape of a fan. On the other hand, in the Palaeodictyoptera where the wings grew sidewards, the traeheal ramifications or veins must have run in curves toward the posterior edge. In fact, the Palaeodictyoptera do show the veins extended in even arches toward the posterior edge, and the subcosta tends to end farther out in the wing.
5) On the basis of wing growth type, the Pterygota can be divided into two group: The Plagioptera, including those with lateral wing pads, such as Palaeodictyoptera, Protohemiptera, Megasecoptera, Protodonata, and Odonata; and the Opisthopterit, including those with posteriorly directed wing pads, the rest of the orders. On this criterion the Ephemeroptera and Odonata are placed on different lines, and not in the same group, as in the division into Paleoptera and Neoptera. Here LEMCHE finds support on the morphology of these two groups. The Ephemeroptera actually show a thoracic musculature and wing base comparable to those of the Neoptera (MATSUDA, 26). On the other hand the Odonata show a wing base quite different from any other living insect. They have, instead of the epipleural and axillary sclerites, two large plates. Also the thoracic musculature of Odonata is different from any other insect; it is composed mainly of dorsoventral muscles, which perform the depression and elevation of the wings. The depressors of the wing are dorsoventral muscles inserted on the two large plates. The pleural wing process has two arms, each one supporting one of the large plates of the wing base. The depression and elevation of the wings in Odonata was solved, according to SNODGRASS (36), as follows: "Since the dorsal walls of the paranotal lobes or the wing bases are lateral extensions of the notum, muscles here attached have simply retained their original notal connections. The problem of the early dragon flies then was to divide these muscles into two functional groups pulling on opposite sides of the wing fulcrum, The problem was readily solved by curving the wing fulcrum inward until it supported the wing base between lateral and mesal groups of the muscles (fig. 23B). The muscles attached mesad of the fulcrum thus became wing elevators (B) and those laterad of it became wing depressors (C): "In all other insects the depression of the wings is performed by longitudinal muscles, instead of dorsoventral muscles as in Odonata. The flexion of the wing is possible by the position of epipleural and axillary sclerites. The flexor of the wing is a muscle (or muscles) inserted on the third axillary sclerite; therefore it is present in all the wing flexing insects (in: Odonata, a muscle from the pleural ridge to the posterior or axillary plate may correspond to the third axillary .muscle of other insects). The extension of the wing is accomplished mainly by the basalar muscles. The main reason why the Ephemeroptera cannot flex the wings is that the muscle of the third axillary sclerite is absent (according to MATSUDA, a muscle from the pleural ridge to the second axillary, in Ephemeroptera, probably corresponds to the muscle of the third axillary). It should be pointed. out also that according to HENRIKSEN (15), the Odonata molt in a way somewhat different from any other insect. The usual manner is. a splitting of the ski n along the middle of the tergum of the thorax. In Odonata this middle splitting of the thorax is confined only to the prothorax, and anterior portion of the mesothorax, the ecdysial line then branches toward the bases of the wings and runs backwards and out. wards on each sie.
Of all the living groups of Pterygota the Ephemeroptera seem to be the most primitive. This, of course, does not mean that the ephemerids are not specialized in any way. On the contrary, every living group or form tends to show a combination of primitive and specialized characters, often to such an extent that it may disconcert the most expert phylogenist. Nevertheless, the balance of these characters, and what is still more -significant, the importance of these features which are considered primitive or specialized, will determine more or less the position of the group under consideration. To determine the importance of a character in a group it must be compared with those which are believed to be more primitive and more advanced. The primitive characters of the Ephemeroptera are best seen in the nymphs. They are the following:
1) The hypopharynx is a three-lobed structure, as in the Symphyla, Diplura, Collembola, and Microcoryphia (Machilis).
2) The mandibles with only a single articulation, as in Machilis.
3) Tentorium resembling that of Lepisma, with a broad central plate and four supporting arms; the anterior arms are likewise ventral in position as in Thysanura.
4) The nymphal gills of ephemerids have been homologized with the abdominal styli of Thysanura. However, there is not enough proof of their homology but the concept is reasonable and not too improbable.
5) Apical appendages of the abdomen (cerci and caudal filaments) similar to those of Thysanura.
The adult mayfly also presents some primitive characters:
1) The wing-venation of these forms is considered to be the most primitive of all living winged insects by TILLYARD (40) and EDMUNDS and TRAVER (10). This is based on the fact that the wing approaches the hypothetical archetype wing-venation (COMSTOCK, 3, and SNODGRASS, 33); with intercalary veins and a fluted condition. The last two features have been regarded by several workets as specializations rather than primitive characters. However EDMUNDS and TRAVER point out that probably the first winged insect had a completely fluted wing, with alternation of concave and convex veins. This was very important to strengthen the wing and necessary before the wing could become thin. Such thinning of the wing was a requisite to improve the sculling action of flight, and constitutes a trend found in most insects in which the wings are rigid along the anterior margin and membranous behind.
2) The Ephemeroptera are the only Pterygota that undergo a molt after reaching the state of imago. This seems to indicate a holdover from the Apterygota ancestors which molt periodically throughout life.
3) The male has paired gonopores with corresponding double penis. The female possesses also paired gonopores which are rather unique among insects; these gonopores are found on the seventh abdominal segment.
4) Thysanura-like ovaries.
The Odonata are an isolated group, a single side branch in the phylogenetic tree; this is also true of the Ephemeroptera. The features which make the dragon-flies an isolated group have been described. However, among all the living orders of Pterygota the Ephemeroptera are the closest to them. This fact is indicated by the following characters that they have in common.
1) lnability to flex the wing on the back of the abdomen, although morphologically speaking this common character may lack any significance,
2) A rather primitive wing venation, conserving the anterior media, which in most modern insects has been lost. In Ephemeroptera both branches of the media are retained (MA and MP), in Odonata only the primitive anterior media is retained. The wing shows more or less a regular alternation of convex and concave veins, at least in the sub-basal region. Anal veins tend to curve toward the rear.
3) Malphighian tubes numerous.
It should be remembered, however, that the Ephemeroptera and the Odonata are not related insects. As SNODGRASS (36) has indicated "they represent two early lines of pterygote evolution differentiated by the method adopted for moving the wings."
The second major explosive evolution of the insects began with the capacity to flex the wings, and is represented today by the so called Neoptera. In spite of the great diversity of the neopterous orders they can be characterized as follows:
1) Ability to fold the wings on the back of the abdomen, as a result of (he presence of the muscle of the third axillary sclerite.
2) A new region in the wing is developed, the jugal region or neala.
3) The anterior media is usually lost, thus leaving the entire spaces between R1 and Cu, with only concave veins. Anal veins are directed obliquely toward the posterior margin of the wing.
The Neoptera are represented by three main evolutionary trends or groups; The orthopteroid, hemipteroid, and neuroptcroid. MARTYNOV named them, according to the evolution of the jugal region or neala, Polyneoptera (orthopteroid) with well developed neala containing several veins; Paraneoptera (hemipteroid) with only a simple or a branched vein in the jugal region and Oligoneoptera (neuropteroid) with a simple and longitudinal vein in the neala. The characters of these groups are tabulated in table 2.

The characters indicated are trends rather than clear cut features. Some of the exceptions are characters secondary lost or primitively retained.
The Orthopteroid Group
In this group we can distinguish two lines: the orthopteroids proper, with clear affinities, and a line represented by Plecoptera, Embioptera and Dermaptera, with doubtful affinities (although Dermaptera be closer to the typical orthopteroids). The distinguishing trends of these two lines are indicated in table 3.
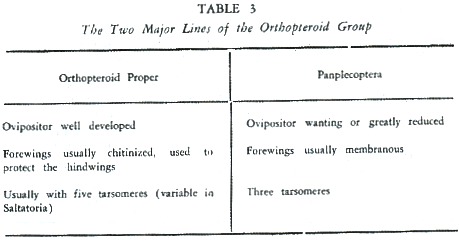
ln the orthopteroids proper we find two distinct lines: the Panorthoptera, represented by Grylloblattodea, SaItatoria, and Phasmida. The Dictyoptera, represented by BIattaria, Mantodea and Isoptera. The distinguishing characteristics of these two lines are indicated in table 4.

An interesting fact, as pointed out by SNODGRASS (36), is that the dorsal longitudinal muscles of the thorax, in the Dictyoptera, "are either absent or are too small to have any direct action on the wings. "For which reason these insects also make use of the dorsoventral muscles for wing motors, but not in the same way as do the dragonflies." The method by which the Dictyoptera move their wings is not well understood. To all these, Snodgrass adds that the Dictyoptera, "on the basis of their wing musculature and thoracic structure, are appropriately separated in classification from the rest of the orthopteroid insects."
Although the termites superficially seem to be far apart from the roaches, there is little doubt that they arose from roach-like insects. A close examination will reveal the true relation of these two groups ,Table 5).

To all these characters in common it can be added that according to JUDD (21), the structure of the proventriculus in both groups indicates a close relationship.
Regarding the Panplecoptera, OTAMPTON (6) has summarized the evidence that the Embioptera and Plecoptera are related as follows:
1) Mesothoracic postscutelluro well developed.
2) Mesotrochantin basally fused with the pleuron.
3) Mesothoracic coxae tend to become ring-like.
4) Three tarsomeres.
5) Ovipositor lacking.
The Hemipteroid Group
The Zoraptera are placed here as the most primitive group of the hemipteroids, at the base ,of their evolutionary branch and clase to the orthopteroids. The special position of Zoraptera is shown by the multiple affinities with the hemipteroids and orthopteroids. DELAMARE-DEBOUTTEVILLE (8, 9) has indicated remarkable similarities between Zoraptera and Isoptera (the latero-pleural region of wingless forms as compared with the winged forms in both groups, also the resemblance of the propleuron in both orders, plus the ability of the winged forms to shed the wings).
The main evolutionary trend in the hemipteroids is a gradual development of sucking mouth-parts. The Zoraptera present a conservative type of mouth-parts with only slightly elongated maxillae. In Corrodentia the maxillae present a peculiar modification, the lacinia seems to have detached completely from the stipes, forming a chisel-like rod, toothed or forked at its apex. This rod is provided with a protractor muscle from the stipes of the maxilla and a retractor muscle from the head wall. Another important modification is found in the hypopharynx. The lingual sclerites (basal bars) of the typical hypopharynx have been transformed into a pair of large sclerites of ovoid shape. The suspensory sclerites and the sitophore have been modified also into a conspicuous cup-shaped sclerite, placed just before the mouth. These two structures are connected by a branched filament. The ovoid sclerites have been regarded as glands, and the filament as a duct. This concept has been proved to be wrong, as WEBER (45) and others have shown conclusively that the sclerites in question do not have a glandular structure in the epithelium beneath them, and are devoid of lumen. According to WEBER (45) there is a small and hard process arising from the clypeal wall of the preoral cavity which fits into the cup-shaped sclerite thus forming a "mortar-and-pestle" apparatus (v. SNODGRASS, 34, p. 25). These morphological changes are also found in the Mallophaga; a rodlike lacinia free fram the maxilla, with the corresponding retractor muscle from the head wall, and protractor fibers from the stipes. This rod, however, has been lost in many Mallophaga. The ligular scIerites and the suspensory sclerites, including the sitophore, have undergone the same transformation as in Corrodentia, The function of the sitophore scIerite in Mallophaga is unknown, an opposing "pestle" as in Corrodentia being absent in this order.
The Anoplura are a specialized side branch of the Mallophaga, in which the mouth-parts have been highly modified. In Thysanoptera we already find a short and conical beak with three styli. One stylet is the transformed left mandible, the right mandible is absent or very reduced. The other two styli correspond to the maxillary lacinia. REYNE (28) has shown that the maxillary stylet of the Thysanoptera is developed from a secondarily detached part of the maxilla. Thus, the maxillary styli would correspond to the chisel-like rods of Corrodentia and Mallophaga, that have become elongated and setiform. In some forms the maxillary stylet is connected with the base of the stipes by a lever arm. According to SNODGRASS (34) this lever arm belongs to the lacinia. No lever has been differentiated in Corrodentia and Mallophaga. The maxillary stipes and galea are placed at the sides of the conical beak.
In Hemiptera-Homoptera the beak reaches the highest development. The maxillary structure of Thysanoptera is again found in this group. This is best seen in cicadas, in which the maxillary stylet is connected with the stipes by a lever sclerite, just as in Thysanoptera (v. SNODGRASS, 34, p. 97), The main difference consists in that the maxillary lobe has become fused with the lateral wall of the head and the maxillary palpus is absent.
The Neoropteroid Group
Like the Hemipteroids, the Neuropteroids probably descended from forms closely allied to Protorthoptera. Therefore it is not surprising to find that some forms (Coleoptera) are very orthopteroid in several features. As a matter of fact, WEBER (46) includes the Coleoptera among the orthopteroids. The complicated interrelationships among the neuropteroids make it very difficult to establish evolutionary trends and exact relations of these orders. The Neuroptera (in the broad sense) and Mecoptera are the, most primitive orders and probably originated from a common stock. Although Neuropteraand Mecoptera are closely related they are placed here as representing two lines of evolution: The Panneuroptera including Neuroptera, Megaloptera, Raphidiodea, Coleoptera, Strepsiptera and Hymenoptera, and the Panmetoptera (= Panorpoidea) including Mecoptera, Trichoptera, Lepidoptera, Diptera and Siphonaptera. In both groups there is a tendency toward developing sucking mouth-parts. At the base of the two groups (Mecoptera-Neuroptera) we find that the mesothoracic coxa is divided into eucoxa and meron, but in the Panneuroptera line this division of the coxa is lost (Coleoptera, Strepsiptera, and Hymenoptera). On the other hand, most of the forms of the Panmecoptera retain the meron. Males of the Panneuroptera tend to develop processes on the gonocoxopodites, usually called volsellae (found in Neuroptera, Cupedidae in Colcoptera, and Hymenoptera).
Recently, HINTON (17) has elevated the family Boreidae of Mecoptera to ordinal rank (Neomecoptera ). The main distinctions between the Boreidae and Mecoptera proper are given by Hinton follows:
Larva:
1) The cranium is without a distinct epistomal suture.
2) The cardo is not fused to the basistipes, and the tentorial adductors of the cardo are present; if homologues of these muscles are present in the Mecoptera they are now tentorial adductors of the cardostipes.
3) The postmentum is well developed instead of lost or reduced to an articulating membrane between the prementum and the cranium.
4) The abdomen lacks prolegs, whereas in the Mecoptera prolegs are present on the first eight abdominal segments.
5) The larvae feed on moss, whereas those of the Mecoptera are carnivorous.
Adult:
1) The ovaries are panoistic instead of polytrophic as in the Mecoptera.
2) The 11th abdominal segment of the female lacks cerci, whereas one-or two-segmented cerci are present in the Mecoptera.
3) The ninth and tenth abdominal segments are modified to form a large functional ovipositor but no such "ovipositor" is found in the Mecoptera.
4) The adult gut lacks the .six special rectal glands of the Mecoptera.
AIthough the Boreidae seem actually a well differentiated group among the Mecoptera, it still has a great number of characters in common with that order, indicating its relation with it. For this reason, it would be more logical to retain the Boreidae as a suborder of Mecoptera, rather than forming an entirely new order for it.
The relation of Trichoptera to Mecoptera is well illustrated by comparing the wing venation of a primitive Caddis-fly such as Stenopsychodes hiemalis (Policentroprodidae) with that of the fossil group Paratrichoptera (a mecopteron). They resembIe each other as follows: A four-branched Rs; staIk of R2 + 3, longer than that of R4 + 5; a four-branched M; stalk of M1 + 2 Ianger than that of M3 + 4; with a closed radial cell (re), and a closed median cell (mc). The Lepidoptera are very closely related to Trichoptera (Table 6); it seems that both originated from a common ancestor. The Micropterygidae, although the most primitive of all the Lepidoptera, have been usually regarded as belonging to that order. However, HINTON (16) has suggested that it should be regarded as a distinct order (Zeugloptera). Although there is no reason for excluding the Micropterygidae from the Lepidoptera on the basis of adult and pupa structure, HINTON (17) in a recent paper listed the following important differences between the larva of the Micropterygidae and Lepidoptera:

1) The cranium of the Zeugloptera lacks the adfrontal ridge and adfrontal sutures.
2) The tentorial bridge is short and broad as in the Mecoptera instead of being long and narrow.
3) The anterior tentorial pit is dose to the inner side of the antennal base instead of far behind the antenna.
4) The maxilla has a separate galea and lacinia instead of an undivided lobe.
5) A cranial flexor of the dististipes is present.
6) Lateral labral rettactors ate present.
7) A pair of cibarial muscles is inserted in the labrum.
8) A spinneret is absent.
9) The leg has the coxa, trochanter, and femur fused but the tarsus and pretarsus are discrete segments, a type of reduction unknown in the Lepidoptera.
10) The ventral abdominal prolegs lack retractor muscles.
11) The spiracles of the metathorax are functional instead of non-functional.
12) The chaetotaxy of the thorax and abdomen is of quite a different type the distribution of the primary setae of the two orders is entirely different.
Furthermore, he adds that "in many respects, especially in the structure and musculature of the head and mouth-parts of the larva, the Zeugloptera appear to be much more closely related to the Trichoptera than to the Lepidoptera."
The, Diptera seem to have originated from a Mecoptera-like insect. This is indicated by the following features in common:
1) The protodipteron, Permotipula patricia (a tipuloid dipteron) has a wing venation very similar to that of Paratrichoptera, the main difference between the two is the narrowing of the wing base in Permotipula, and as a consequence the third anal vein is missing, and the second one reduced.
2) According to TILLYARD (41) the family Nanoochoristidae (Mecoptera) are the closest living relatives of the Diptera. This family has a head-capsule and mouth-parts similar to that of some flies. This is well seen by comparing the head of Nannochorista with that of Edwardsina (Blepharoceridae). In both types we find five-segmented maxillary palpi, with a very short basal segment and the third with a peculiar sense-organ; two-segmented labial palpi, which are placed very close together forming a labellum-like structure. It should be pointed out that CRAMPTON (4, 5), has indicated that the labellar lobes are probably the labial palpi in Diptera. Also the labium-epipharynx and mandibles are elongated (bur more so in Edwardsina).
3) The genitalia of certain Diptera are very similar to those of Mecoptera. For example, in the Tanyderidae (Diptera) and in the Meropeidae (Mecoptera) the gonopods are transformed into large forceps with reduction of all the other parts.
As to the Siphonaptera the origin is still more doubtful, perhaps both Diptera and Sjphonaptera originated from a common ancestor in the Mecoptera-Triehoptera complex.
With the orders of Panneuroptera one is faced immediately with unsolved phylogenetic problems, and all that can be done here is merely to mention some of the suggestions which have been made.
The similarity of certain coleopterous larvae with those of Neuroptera may indicate a possible relationship. The family Cupedidae, considered to be the most primitive of all the Coleoptera, shows reticulated elytra, simulating veins. This, it is said, may indicate that the elytra of Coleoptera may have arisen by a condensation of the veins from a wing like that of Neuroptera. In a very recent paper, CROWSON (7) has pointed out that "the remarkable lower Permian fossil Tshekardocoleus looks like an intermediate form between cupedid-type Coleoptera and Megaloptera of the Corydalis type and may be taken as direct evidence in support of the neuropteran theory... The fossil evidence seems to be at least consistent with the theory that Coleoptera took their origin from megaloptera-like ancestors during the Permian period, very soon after the first adaptive radiation of the Endopterygota."
That Strepsiptera are related to Coleoptera is and has been the general opinion among entomologists. But JEANNEL (19) has pointed out that the Strepsiptera seem to be more dosely related to Hymenoptera than to Coleoptera. This is supported, according to him, by the following characters shared by both the Strepsiptera and Hymenoptera.
1) First abdominal segment incorporated to the thorax (Clistogastra among the Hymenoptera), and with epimeron and episternum obliquely placed.
2) The head completely free from the prothorax, and orthognathous* (this is different from that of most Coleoptera).
3) The fIabellated antennae of Strepsiptera are as similar to those of some tenthredinids as to those of Rhipidit.J.
4) The legs of Mengeidae are of the hymenopterous type. A five-segmented tarsus with the basal segment (basitarsus) enlarged. The structure of the empodium resembles that of the Hymenoptera.
5) The triungulins of Strepsiptera are actually more similar to the planidium of chalcids than to those of meloids and rhipiphorids.
6) The Strepsiptera and the Hymenoptera are the only two orders of pterygotes in which the embryo has only one envelope instead of the usual two (amnion and serosa).
7) Polyembryony is known to occur in Halictophagidae, and also in some Hymenoptera, especially in chalcids.
However, it should. be pointed out that most of the above characters are also found in some Coleoptera and furthermore many of these characters can be explained as due to other reasons than relationship. Thus, the doubled antennae of Strepsiptera probably arose quite independently from similar conditions in either Hymonoptera of Coleoptera. Although the Strepsiptera and Hymenoptera have only one envelope in the developing embryo, it is the amnion which is present in the Strepsiptera, while in Hymenoptera it is the serosa. Actually, it appears, as CROWSON (7) has indicated, "that no features of the Stylopoidea are inconsistent with a derivation of the group from normal coleopterous ancestors." He gives the following features as an indication of the relationship of the Strepsiptera and Coleoptera:
1) The use only of the hind wings in flight.
2) The more extensive sclerotization of the abdominal sternites than of the tergites (contrary to the usual endopterygote condition).
3) The nature of the metendosternite.
4) The structure of the first-instar larva, which does not seem to differ from similar "triungulins" of Meloidae and Rhipiphoridae in any character which could possibly be regarded as of oardinal importance.
Conclusions
1) Our present knowledge seems to indicate that the closest relatives of insects are the myriapods, of which, the Symphyla have the greatest number of characters in common with most primitive insects. This may suggest that the Insecta and Symphyla were derived from a common stock of myriapod-like arthropods. There is also some evidence that the insects may have originated by neoteny from myriapod-like arthopods.
2) The exclusion of the entognathous apterygotes from the class Insecta does not seem to be well justified.
3) There is good evidence that the entognathous apterygotes (Protura, Collembola, Diplura) are interrelated.
4) Since the differences between lepismatids and machilids include several features of great phylogenetic importance (Table 1), they are regarded here as representing two orders: the Microcoryphia, including the machilids, and the Thysanura proper, including the lepismatids.
5) The origin of the pterygotes from the apterygotes is still a question of much speculation. There are three main theories as to the origin of the wings; all these views agree in that the wings were developed from paranotal processes on the meso- and meta-thorax. a) According to MARTYNOV the paranatal extensions tended to grow sidewards and were used as gliding organs. The wings originated by the acquisition of a line of flexibility at the base of each process. Later some acquired the ability to flex the wings over the back of the abdomen. b) SCHWANWITSCH, on the other hand, believes that the paranota were directed backwards and became movable from the very beginning of their evolution. c) Finally, according to LEMCHE, wings originated from two different types of growth of the paranotal processes. In one, the growth was sidewards, giving rise to the Palaeodictyoptera, Protohemiptera, Protodonata, Odonata, and Megasecoptera. In the other, growth was backwards and gave origin to the rest of the orders.
6) Of all the living groups of Pterygota the Ephemeroptera seem to be the most primitive. The Odonata are an isolated group, this is also true of the Ephemeroptera. However, of all the living orders of Pterygota the Ephemeroptera are the closest to Odonata.
7) The Neopteran orders comprise three main evolutionary trends or groups: The orthopteroid, hemipteroid, and neuropteroid lines (Table 2).
8) In the orthopteroid group two lines can be distinguished: The orthopteroid proper, with clear affinities, and a line (Panplecoptera) represented by Plecoptera, Embioptera, and Dermaptera (Table 3) with doubtful affinities. In the orthopteroid proper two other lines can be recognized: The Panorthoptera, represented by GrylIoblattodea, Saltatoria, and Phasmida; the Dictyoptera, represented by Blattaria, Mantodea and Isoptera (Table 4). Although the termites superficialIy seem to be far apart from the roaches, carefnl examination will reveal a close relation of these two groups (Table 5).
9) The main evolutionary trend in the hemipteroids is a gradual development of sucking mouth-parts.
10) Two lines of evolution can be recognized in the neuropteroid group: The Panneuroptera including Neuroptera, Megaloptera, Raphidiodea, Coleoptera, Strepsiptem and Hymenoptera, and the Panmecoptera ( = Panorpoidea) including Mecoptera, Trichoptera, Lepidoptera, Diptera and Siphonaptera. In both groups there is a tendency toward developing sucking mouth-parts.
By necessity, the above account has be en somewhat superficial and to a certain degree dogmatic. This; it is hoped, shall be corrected in the future as our knowledge on these matters expands. It is also hoped that more and more entomologists shall become aware of the scientific importance of this fascinating and everlasting challenge which the phylogeny and the interrelationship of the insect orders offer to anyone interested.
Resumen
El trabajo presente consiste en un análisis y evaluación de la literatura sobre la evolución e interrelación de los órdenes de los insectos, dando como resultado las siguientes conclusiones:
1) Nuestros conocimientos actuales parecen indicar que los artrópodos más cercanos a 105 insectos son los miriápodos, de los cuales los Symphyla poseen el mayor número de caracteres en común con los insectos primitivos. Esto parece sugerir que Insecta y Symphyla se derivaron de un tronco común, formado de artrópodos de tipo miriápodo. Hay evidencia también de que los insectos pueden haberse originado por medio de neotenia de dicho tronco común.
2) La tendencia reciente de excluir los apterigotos entognatos de la clase Insecta no parece estar bien justificada.
3) Investigaciones recientes indican que los apterigotos entognatos (Protura, Collembola y Diplura) están interrelacionados.
4) En vista de que las diferencias entre lepismátidos y maquílidos incluyen varias características de gran importancia filogenética (Cuadro 1), se considera que forman dos órdenes: Microcoryphia, que comprende a los maquílidos, y Thysanura propiamente dichos, o sea los lepismátidos.
5) El origen de los pterigotos aun continúa sin solución.
6) De todos los grupos vivientes de Pterygota los Ephemeroptera parecen ser los más primitivos. Los Odonata forman un grupo aislado, lo cual también es cierto de los Ephemeroptera, pero de todos los órdenes vivientes de Pterygota los efemerópteros son los que más se aproximan a los odonatos.
7) Los órdenes neópteros comprenden tres grupos o tendencias evolucionarias principales: Las líneas Orthopteroidea, Hemipteroidea y Neuropteroidea (Cuadro 2).
8) En el grupo ortopteroideo se pueden distinguir dos líneas: los Orthopteroidea propiamente dichos, con afinidades claras, y una línea (Panplecoptera) representada por Plecoptera, Embioptera, y Dermaptera (Cuadro 3), con afinidades dudosas. En el grupo ortopteroideo propiamente dicho pueden reconocerse ottas dos líneas: los Panorthoptera, representados por Grylloblattodea, Saltatoria, y Phasmida; los Dietyoptera, representados por Blattaria, Mantodea e Isoptera (Cuadro 4). Aunque los comejenes (Isoptera) superficialmente parecen estar muy lejanos de las cucarachas, un examen cuidadoso revelará una relación cercana entre los dos grupos (Cuadro 5).
9) La principal tendencia evolucionaria en los hemipteroideos es un desarrollo gradual de un aparato bucal chupador.
10) Dos líneas de evolución pueden ser reconocidas en el grupo neuropteroideo: los Panneuroptera incluyendo Neuroptera, Megaloptera, Raphidiodea, Coleoptera, Strepsiptera e Hymenoptera, y los Panmecoptera (= Panorpoidea) incluyendo Mecoptera, Trichoptera, Lepidoptera, Diptera y Siphonap.tera. En ambos grupos hay una tendencia hacia el desarrollo de un aparato bucal chupador.
Schlussfolgerungen
Ein ausführliches Studium und Schatzung der Literatur über die Evolution und Beziehungen der verschiedenen Insektenordnungen erlauben dem Verfasser wie folgt abzuschliessen:
1) Die gegenwartige Kcnntnis scheint hinzuweisen, dass die Myriapoden die den Insekten nahre Gliederfüssler sind, von denen die Symphila die meisten charakteristischen Merkmale gemeinsam mit den ursprünglichen Insekten besitzen. Das Hisst schliessen, dass Insecta und Symphila vall einem gemeinsamen, van myriapoden-artigen Gliederfüsslern gebildeten Stamm sich ableiten. Es besteht aneh Merkmale, die daranf hinweisen, dass die Insekten vermittels Neotenie von besagten Stamm herrühren.
2) Es scheint, dass es keinen Beweis gibt, um die entognatischen Apterygoten von der Klasse Insecta ausznsehliessen.
3). Neueste Untersuchungen weisen darauf hin, dass die entognatischen Apterygoten (Protura, Collembola, Diplura) in Beziehung zueinander sind.
4) Da die Untersehiede zwisehen Lepismatiden und Machiliden mehrere Merkmale von grosser phylogenetischer Bedeutung einsehliessen (Tafel 1), betrachtet der Verfasser die Bildung zweier Ordnungen: die Microeoryphia, weldle die Maehilíden umfassen, und die Thysanura, welche die Lepismatiden sind.
5) Der Ursprung der Pterygoten bleibt naeh wie vor im Ungewissen.
6) Von allen lebenden Gruppen der Pterygoten scheinen die Epheme roptera die Primitivsten zu sein. Die Odonata bilden eine Gruppe für sieh, was auch für die Ephemeroptera erwiesen ist, aber von allen lebenden Ordnungen der Pt-erygota sind es die Ephemeroptera die am meisten den Odonata ahnlích sind.
7) Die Ordnungen der Neoptera bestehen aus drei Gruppen oder hauptsachlichen evolutionären Bestrebungen: die Orthopteroidea; Hemiptcroidca und Neuroptcroidca (Tafel 2).
8) In der Gruppe der Orthopteroidea kann man zwei Linien unterseheiden: die tatsachlichen Orthopteroidea mit bestimmten Affinitaten und cinc Linie (Panplecoptera) gebildet durch Plecoptera, Embioptera und Dermaptera (Tafel 3) mit undeutlichcn Affinitaten. In der Gruppe der tatsacWichen Orthop. teroidea lassen sich weitere zwei Linien bestimmen: die Panorthoptera, gebíldet durch Grylloblattoidea, Saltatoria und Phasmida; die Dictyoptera, gebildet dureh Blattaria, Mantodea und Isoptera (Tafel 4). Wennglcich die Termiten oberflachlicherwcise weit entfernt von den Schaben sind, ein genaues Examen wird eine nahe Beziehung zwischen beiden Gruppen aufweisen (Tafel 5).
9) Die vorherrschcnde cvolutionare Bestrebung bei den Hemipteroidea ist cine abgcstufte Entwicklung des Saugwerkzeuges.
10) Zwei evolutionare Linien lassen sich in der Gruppe der Neuropteroidea erkennen: die Panneuroptera einschliesslích die Neuroptera, Megaloptera, Raphidiodea, Coleoptera, Strepsiptera und Hymenoptera, und die Panmecoptera (= Panorpoidea) cinsehliesslích die Mecoptera, Trichoptera, Lepidoptera, Diptera und Siphonaptera. In beiden Gruppen besteht die Tendenz zur Entwícklung eines Saugapparates.
Literature Cited
1. BEER, G. R. DE 1940. Embryos and ancestors, x + 108 pp., Oxford University Press. [ Links ]
2. CARPENTER, F. M. 1953. The evolution of Insects. American Scientistis, 41: 256-270. [ Links ]
3. COMSTOCK, J. H, 1918. The wings of Insects. 430 pp., 10 pIs., Ithaca, Comstock Publishing Co. [ Links ]
4. CRAMPTON, G. C. 1923. A comparison of the labium in certain holometabolous insects from the standpoint of phylogeny. Proc. Ent. Soc. Wash., 25: 171-180. [ Links ]
5. CRAMPTON, G. C. 1925. A phylogenetic study of the labium of holometabolous insects, with particular reference to the Diptera. Proc. Ent. Soc., Wash., 27: 68-91. [ Links ]
6. CRAMPTON. G. C. 1935. The interrelationships and lines of descent of living insects. Pomona Jour. Ent. Zool., 45: 165-181. [ Links ]
7. CROWSON. R. A. 1960. The phylogeny of Coleoptera. Ann. Rev. Ent., 5: 111-134. [ Links ]
8. DBLAMARE-DEBOUTEWILLE. C. 1948, Sur la morphologie des adultes apteres et ailés de Zorapteres. Ann. Sci. Nat, 9: 145-154. [ Links ]
9. DELAMARE.DEBOUTEVILLE. C. 1948. Observations sur l'écologie et léthologie des Zorapteres. La question de leur vie sociale et de leurs prétendus rapports avec les termites. Rev. ent., 19: 347-352.
10. EDMUNDS, G. F. JR., & J. R. TRAVER 1954. The flight mechanics and evolution of the wings of the Ephemeroptera, with notes on the archetype insect wing. Juor. Wash. Acad. Sci., 44: 390-400. [ Links ]
11. EWING, H. E, 1943. The origin and classification of the Apterygota. Proc. Ent. Soc. Wash., 44: 75-98. [ Links ]
12. FOLSOM, J, W. 1900. The development of the mouth-parts of Anuria Maritina. Guér. Bull. Mus. Comp. Zool., 36: 87-157. [ Links ]
13. HANSEN, H. J. 1930. Studies on Arthropoda III. On the comparative morphology of the appendages in the Arthropoda. 376 pp., 16 pIs. Copenhagen. [ Links ]
14. HENNING, W. 1953. Kritische Bemerkungen zum phylogenetischen System der Insekten. Beitr Ent., 3: 1-85. [ Links ]
15. HENRIKSEN, K. 1932, The manner of moulting in Arthropods. Notul. Ent., 11: 103-127. [ Links ]
16. HINTON, H. E. 1946. On the homology and nomenclature of the setae of Lepídopterous larvae, with some notes on the phylogeny of the Lepídoptera. Trans R. Ent Soc. Lond., 97: 1-37. [ Links ]
17. HINTON, H. E. 1958. The phyIogeny of the panorpoid orders. Ann. Rev. Ent., 3: 181-206. [ Links ]
18. IMMs, A. D. 1945,. The PhyIogeny of Insects. Tijdscbr. Ent., 88: 63-66. [ Links ]
19. JEANNEL, R. 1945. Sur la position systémtique des Strcpsipteres. Rev. Franc. Ent., 11: 11-118. [ Links ]
20. JOHANNSEN, O. A. & F. H. BUTT 1941. Embryrology of insects and myriapods. xi + 462 pp., 370 figs. McGraw Hill Book Company, Inc. [ Links ]
21. JUDD, W. W. 1948. A comparative study of the proventriculus of Orthopteroid insects with references to its use in taxonomy. Canad. J. Res. (D), 26: 93-161. [ Links ]
22. LEMCHE, H. 1940. The origin of winged insects. Vidensk, Medd. Dansk Naturh Foren., 104: 121-168. [ Links ]
23. MANTON, S. M. 1953. Locomotory habits and the evolution of the larger arthropodan groups. Symp. Soco exp. Biol., 7: 339-476. [ Links ]
24. MARCUS, E. 1958. Evolution of the animal phyla. Quart. Rev. Biol., 33: 24-58. [ Links ]
25. MARTYNOV, A. V. 1925. Über zwei Grundtypen del' Fliigel bei den Insekten und ihre Evolution. Zeit. Morph. Oeko/. Tiere, 4; 465-501. [ Links ]
26. MATSUDA, R. 1956. Morphology of the thoracic exoskeleton and musculature of a mayfly Siphlonurus columbianus McDunnough ( SiphoIonuridae, Ephemeroptera). Jour. Kansas Ent. Soc., 29; 92-113. [ Links ]
27. REMINGTON, C. L. 1954. The suprageneric classification of the order Thysanura. Ann. Ent. Soc. Amer., 47: 277-286. [ Links ]
28. REYNE, A. 1927. Untersuehungen über die Mundteile der Thysanopteran. Zool, Jahrb., Anat., 49: 391-500. [ Links ]
29. Ross. H. H. 1955. Evolution of insect orders. Ent. News, 66; 197-208. [ Links ]
30. SCHW ANWITSCH, B. N. 1943. Subdivision of Insecta Pterygota into subordinate groups. Nature, London, 152; 727-728. [ Links ]
31. SCHWANWITSCH, B. N. 1946. On the interrelations of the orders of Insecta Pterygota as dependent on the origin of flight. Zool. Journ. (Moscow) 24:529-542. [ Links ]
32. SIMPSON, G. G. 1945. The principles of classification and a classification of mammals. Bull. Amer, Mus. Nat. Hist., 85: 1-350. [ Links ]
33. SNOPGRASS, R. E. 1935. Principles of insect morphology. ix + 667 pp., 319 figs. McGraw Hill Book Company, Inc., New York. [ Links ]
34. SNODGRASS, R. E. 1944. - The feeding apparatus of biting and sucking insects affecting man and animals. Smithsonian Mise. Coll., 104: 1-113. [ Links ]
35. SNOPGRASS, R. E. 1951. Comparative studies on the head of mandibulates arthropods. viii + 118 pp., 37 figs. Comstock Publishing Company, Inc. [ Links ]
36. SNODGRASS, R. E. 1958. Evolution of arthropod mechanisms. Smithsonian Misc. Coll., 138: 1-77. [ Links ]
37. TIEGS, O. W. 1940. The embryology and affinities of the Symphyla based on a study of Hanseniela agilis. Quart. Journ. Micr. Sci. 82: 1.225.
38. TIEGS., O. W. 1945. The post-embryonic development of Hanseniela agilis (Symphyla). Quart. Journ. Micr. Sci. 85: 191-328. [ Links ]
39. TLEGS, O. W. & S. M. MANTON 1958. The evolution of the Arthropoda. Biol. Rev. Cambridge Phil. Soco 33: 255-337. [ Links ]
40. TILLYARP, R. J. 1932. Kansas Permian insects. Pt. 15. The order PIectoptera. Amer. Jour. Sci., 23: 95.134, 237-272.
41. TILLYARD, R. J. 1935. The evolution of the scorpion flies and their derivatives. Amer. Ent. Soc. Amer., 28: 1.45. [ Links ]
42. TUXEN, S. L. 1931. Monographie der Proturen. I. MorphoIogie nebst Bemerkungen über Systematik und OekoIogie. Z. Morph. Dekol. Tiere, 22: 671.720,20 figs. [ Links ]
43. TUXEN,S. L. 1959. The phylogenetic significance of entognathy in entognathous apterygotes. Smithsonian Misc. Coll., 137: 379-416. [ Links ]
44. WEBER, H. 1936. Copeognatha. Biologie der Tiere Deutschlands. 3921: 1-50. [ Links ]
45. WEBER, H. 1938. Beitriige zur Kenntnis der überordnung Psocoidea. l. Die Labialdrüsen der Copeognathen. Zool. Jahrb., Anat., 64: 243-286. [ Links ]
46. WEBER, H. 1954. Grundiss der Insektenkunde. xi - 428 pp., 220 figs. Gusbav .fisher Velag Stuttgart, 3rd. Edn. [ Links ]
* Departamentocle Entomología, Facultad de Agronomía, Universidad de Costa Rica.
* The exclusion of most of the Apterygota orders from the Insecta by some authors does not seem to be well justified; this controversy shall be discussed at some length later on. In the meantime, al! the orders of the Apterygota are regarded here as belonging to the class lnsecta.
* According to Ryuichi Matsuda, in a personal communication, abdominal legs are more conspicuous in embryos of insects, for which reason loss of abdominal legs cannot be derived by neoteny.
* For a general discussion of the extinct orders see CARPENTER (2) .
* This condition is called "hypognathous" by. American authors.

