










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.49 n.2 San José Jun. 2001
Abstract
This is the first publication dedicated to the diversity of smut fungi in Panama bases on field work, the study of herbarium specimens, and referentes taken from literatura. It includes smuts parasitizing cultivated and wild plants. The latter are mostly found in rural vegetation. Among the 24 species cites here, 14 species are recorded for the first time for Panama. One of them, Sporisorium ovarium, is observes for the first time in Central America. Entyloma spilanthis is found on the host species Acmella papposa var. macrophylla (Asteraceae) for the first time. Entyloma costaricense and Entyloma ecuadorense are considered synonyms of Entyloma compositarum and Entyloma spilanthis respectively. For the new conbination Sponsorium panamensis see note at the end of this publication. Descriptions of the species are complemented by some illustrations, a checklist, and a key.
Key words
Checklist, neotropics, Panama, smut fungi, Sponsorium panamensis, Tilletiales, Ustilaginales
Smut fungi (Ustilaginomycetes and Microbotryales, Urediniomycetes; Basidiomycota; Bauer et al. 1997, 2001) are parasites of plants, especially herbs belonging to the Poaceae and Cyperaceae. Species causing economic losses on crops, e.g. Tilletia barclayana (Bref.) Sacc. & Syd. on rice, Ustilago maydis (DC.) Corda on corn, and Ustilago scitaminea Syd. on sugar cane, are well known, but the majority of species of smut fungi which parasitize plants growing in the wild have rarely been studied, especially in tropical regions.
Checklists of smut fungi for neotropical countries are published for Mexico (Durán 1987; 125 species), Costa Rica (Piepenbring 1996a-c; 54 species), Colombia (Molina-Valero 1980; 40 species), and Cuba (Piepenbring and Rodríguez 1998 a-b; Piepenbring 1999; 39 species). For Panama, the records of 10 species of smut fungi were found scattered in the literatura (Zundel 1939, 1953, Toler et aL 1959, Dennis 1970, Comstock et aL 1983).
The check list in the present publication is far from complete, heing based on only about two months of field work by the author and some data taken from the literatura. This investigation is part of extensive studies on smut fungi in the neotropics, carried out to elaborase a volume of the Flora Neotropica on smut fungi. With this publication the author hopes to stimulate further research on Panamanian smut fungi, which will yield many interesting results since the flora of Panarna is highly diverse.
Materials and Methods
The author spent about two months looking for smut fungi in different provinces of Panama during collecting trips in 1994 to 1998. In addition to the recently collected specimens, which were herbarium-dried like higher plants, some smut fungi were found on specimens of their host plants deposited in PMA (Herbario de la Universidad Nacional de Panamá, Ciudad de Panamá, Panamá).
Specimens collected in Panama are kept in PMA (Herbario del Departamento de Botánica, Universidad de Panamá, Panamá), in USJ (Herbario de la Escuela de Biología, Universidad de Costa Rica, San José, Costa Rica), and in the personal herbarium of the author.
Herbarium-dried spores were measured by light microscopy (LM) in lactophenol after heating. Spore measures include two size ranges, i.e. the smaller and the larger diameter, for globose spores and for flattened ones the thickness as well as the smaller and the larger diameter of the spores, as seen in plane view. The values of size ranges are the means of at least 20 spores ± 1 S.D. Extreme values are given in brackets. Spore measurements include ornamentation, but not hyaline appendages.
For scanning electron microscopy (SEM), herbarium-dried spores were dusted onto double-sided adhesive tape, fixed on specimen stubs, and sputter coated with gold-palladium, ca 20 nm. The spores were studied with a Cambridge Stereoscan 250 MK 2.
Germinations of teliospores were obtained on 1 % water agar (WA) supplemented with the antibiotic tetracycline. Germinating spores were observed using LM on sections of WA covered by a cover slip. Times of gerinination given in the legends refer to how much time had passed from the moment when the teliospores were spread in the petri dishes until the drawing was executed.
Species are presented according to the classification proposed by Bauer et al. (1997, 2001). Synonyms are placed in angular brackets ([]). The "!" following the acronym of a herbarium means that the respective specimen has been studied by the author.
RESULTS AND DISCUSSIONS OF SPECIES
1. Ustilaginomycetes
1.1 Ustilaginomycetidae
1.1.1 Ustilaginales
1.1.1.1 Ustilaginaceae
1.Cintractia axicola (Berk.) Cornu, Ann. Sci. Nat. Bot. VI 15: 279. 1883; Ustilago axicola Berk., Ann. Mag. Nat. Hist. II 9: 200. 1852. Type. On Fimbristylis dichotoma (L.) Vahl (some scirpoid plant). Santo Domingo. Sallé 74; Herb. Berkeley 4 745 (holotype, K!).
Ustilago peribebuyensis Speg., Anales Soc. Ci. Argent. 17: 89. 1884; Cintractia peribebuyensis (Speg.) Speg., Anales Soc. C¡. Argent. 26: 11. 1888; Cintractia peribebuyensis (Speg.) Sawada, Dept. Agric. Gov´t. Res. Inst. Formosa Report 2: 80. 1922; Cintractia axicola (Berk.) Cornu f. peribebuyensis (Speg.) Zambett., Bull. Soc. Mycol. France 95: 414. 1979 (the material he studied was C. limitata). Type. On Fimbristylis dichotoma [F. diphylla] (Cyperus). Paraguay. Cordillera de Peribebuy, 25 Mar 1883, Balansa 3 775 (holotype, LPS!).
Sori around the bases of peduncles, sometimes around the axes of spikelets or around floral organs; when young, covered by whitish brown, early rupturing peridia. Spore mass agglutinated; dark. Spores single; globose, mostly flattened; 10- 12 x 2-14(-16) x (12-)14-17(-18) m m; light to medium reddish brown. Wall ca 1 m m thick; as seen by LM, smooth to finely warty; as seen by SEM, covered by irregular, dense, fine warts. Germination with phragmobasidia; basidial cells mostly conjugate and develop two large, dikaryotic conidia or basidial cells develop basidiospores.
In Panama on Fimbristylis dichotoma [F. annua (All.) Roemer & Schultes, F. diphylla (Retz.) Vahl] (Cyperaceae):
Coclé: La Pintada, alt. ca 420 m, 14 Feb 1995, Piepenbring and Domínguez 1 462 (PMA, USJ 53 534); El Caño, alt. ca. 50 m, 29 Nov 1996, Piepenbring & Domínguez 2 211 (PMA).
Colón: Ciudad Colón, Barriada Sagrada Resurrección, alt. ca. 50 m, 15 Feb 1995, Piepenbring & Domínguez 1 480 (PMA).
Panamá: Canal zone, 1.5 km NW of Paraíso, alt. ca 70 m, 17 Nov 1994, Piepenbring1 231 (PMA); 1 km S of Jardín Botánico Summit, alt. ca. 70 m, 17 Sept 1994, Piepenbring 1 234, 1 235 (PMA); Jardín Botánico Summit, alt. ca. 80 m, 18 Nov 1994, Piepenbring 1265 (PMA); Jardín Botánico Summit, alt. ca 80 m, 27 Sept 1996, Piepenbring & Stein (formerly Wiech) 2 208 (PMA).
Reported for Panama by Zundel (1953) and Dennis (1970). The host species Scirpus brizoides Benth, mentioned by Dennis (1970), is doubtful.
2. Cintractia fimbristylicolaPavgi & Mundk., Indian Phytopathol. 1: 108. 1949. Type. On Fimbristylis complanata (Retz.) Link. India. Near Chatrapur, Ganjain, Orissa, 30 Aug 1904, Butler (type, ND, n. v.).
Sori few to numerous in an infected inflorescence; around ovaries; when young, covered by whitish brown, early rupturing peridia. Spore mass agglutinated; dark. Spores single; spherical to globose, sometimes flattened or bluntly angular; (7-)8- 0 x (9-)10-12(-13) m m; light reddish brown. Wall ca 1 m m thick; as seen by LM, irregularly warty; as seen by SEM, warts partly confluent forming an irregular reticulum. Gemination with phragmobasidia carrying basidiospores on sterigmata or growing with hyphae.
In Panama on Fimbristylis spadicea (L.) Vahl (Cyperaceae):
Herrera: Parque Nacional Sarigua, alt. ca. 5 m, 30 Sep 1996, Piepenbring & Domínguez 2 213 (PMA).
3. Cintractia limitataG. P. Clinton, Proc. Boston Soc. Nat. Hist. 31: 399. 1904; Cintractia utriculicola (Henn.) G. P. Clinton var. limitata (G. P. Clinton) Hirschh, Farlowia 3: 82. 1947. Type. On Cyperus ligularis L. Puerto Rico. Mayagüez, 23 Apr 1904, Clinton s. n. (holotype, BPI, according to Ling 1950, n. v; isotypes, M!, NY!).
Sori around the bases of peduncles, sometimes around the axes of spikelets or around floral organs; when young, covered by whitish brown, early rupturing peridia. Spore mass agglutinated; dark. Spores single; globose, mostly flattened; (7-) 8-9 (- 10) x 8-10 (-12) x (9-) 11-13 (-14) m m; medium reddish brown. Wall ca 1 m m thick; as seen by LM, almost smooth; as seen by SEM, covered by irregular, dense, fine warts. Germination with phragmobasidia; basidial cells mostly conjugase and develop two large, dikaryotic conidia, or basidial cells develop basidiospores.
In Panama on Cyperus ligularis (Cyperaceae):
Los Santos: 1 km W of Playa Monagre, alt. ca. 5 m, 1 Oct 1996, Piepenbring &Domínguez 2 221 (PMA).
On Cyperus rotundas L.:
Coclé: El Caño, alt. ca. 50 m, 29 Sep 1996, Piepenbring & Domínguez 2 212 (PMA).
Los Santos: Las Tablas, alt. ca. 15 m, 1 Oct 1996, Piepenbring & Domínguez 2 219 (PMA).
Panamá:La Chorrera, alt. ca. 1 300 m, 25 Sep 1994, Piepenbring 1 310 (PMA).
4. Farysia chardonianaZundel, Mycologia 35: 171. 1943. Type. On Carex sp. Venezuela. Carabobo, mountain above Hacienda Cura, Chardón and Alsten (type, BPI 2 772!).
Sori in few to severas spikelets of an infected plant; when young, covered by whitish, early rupturing peridia. Spore mass powdery; brown; permeated by strands of multicellular, dirty white bundles of sterile hyphae (elaters). Spores single; globose or subangular; (5-)6-7 x 6-8 m m; olivebrown. Wall ca 1 mm; covered by dense, fine, less than 0.5 m m high warts, which often fuse forming spirally arranged rows. Germination with phragmobasidia developing basidiospores; threecelled apical parts of the basidia often separate from the rest of the basidium.
In Panama on Carex polystachya Sw. ex Wahlenb. (Cyperaceae):
Chiriquí: East of Boquete, along road to Cerro Azul, alt. ca. 1 300-1 500 m, 1 1 Aug 1974, Croat 26 765 (on PMA 10 160, PMA); At the road to the top of the volcano Barú, alt. ca. 2 100 m, 9 Feb 1995, Piepenbring & Cueva 1 407 (PMA, USJ 53 531l).
5. Leucocintractia scleriae (DC.) M. Piepenbr., Begerow & Oberw., Mycologia 91: 497. 1999; Uredo scleriae DC., in Poir., Encycl. meth. bot. 8: 228. 1808; Ustilago?scleriae (DC.) Tul. & C. Tul., Ann. Sci. Nat. Bot. III 7: 89. 1847; Cintractia scleriae (DC.) L. Ling, Mycologia 43: 314. 1951. Type. On Rhynchospora corymbosa (L.) Britton (quelques espéces de Scleria). French Guiana. Cayenne (no further data) (holotype, G!).
Sori around all the peduncles of a rudimentary inflorescence; cylindrical, often curved; when young, covered by thick, shining white, relalively late rupturing peridia. Sporemass agglutinated; dark. Spores single; globose to flattened; (9-)10-15 x (11-)12-16(-17) m m; light reddish brown. Wall 0.5-1 m m thick; covered by irregular, coarse warts and labyrinthiform to reticulate ridges in a band around the spore. Germination with phragmobasidia developing basidiospores; two-to four-celled apical parts of the basidia usually separate from the spore.
In Panama on Rhynchospora corymbosa [R. aurea Vahl] (Cyperaceae):
Colón: Ciudad Colón, Billa Catharina, alt. ca 60 m, 16 Feb 1995, Piepenbring andDomínguez 1 476 (PMA, USJ 53 536); Puerto Pilón, alt. ca 20 m, 17 Feb 1995, Piepenbring and Domínguez 1 487 (PMA).
On Rhynchospora sp.:
Mindi (?), 28 Feb 1905, Cowell 188 (NY!) (incomplete and badly written label).
Reported for Panama by Zundel (1953) and Dennis (1970) as Cintractia leucoderma (Berk.) Henn. on Rhynchospora corymbosa. They certainly refer to Leucocintractia scleriae because Cintractia leucoderma, now Leucocintractia leucoderma (Berk.) M. Piepenbr., does not infect R, corymbosa (Piepenbring 2000). For the delimitation of the genus Leucocintractia from the genus Cintractia see Piepenbring et al. (1999).
6. "Sphacelotheca "panamensis Zundel and Dunlap, in Zundel, N. Amer. fl. 7: 995. 1939. Type. On Cymbopogon sp. Panamá. Chiriquí (Cherique): near "El Bogrete" (= Boquete?), Sep - Oct 1911, Hitchcock s. n. (type ubi?, not in BPI).
Sori in the ovaries; hidden by enveloping glumes; about 3 mm long; with peridia, which rupture disclosing the spore mass and a slender-pointed columella. Spore mass dusty. Spores single; chiefly subspheric or spheric; 14-18 m m; reddish-brown. Wall thin; smooth to minutely granular, as seen by LM. (Description based on the diagnosis)
In Panama on Cymbopogon sp. (Poaceae). This species is known only from the type specimen collected in Panama. The identification of the host species is very doubtful, because Cymbopogon citratus (DC.) Stapf ¡s the only species of Cymbopogon known from Panama and it flowers only rarely (Davidse and Pohl 1994). In addition to this, the type material, which could not yet be checked, is meagre (Zundel 1939) and the species is only known from the type collection. The validity of this species is therefore very doubtful.
This species belongs to the genus Sporisorium because it is describes from a species of Poaceae and because species of Sphacelotheca infect only dicotyledons (Langdon and Fullerton 1978, Vánky 1987).
7. Sporisorium holwayi(G. P. Clinton and Zundel) Vánky, Mycotaxon 48: 40. 1993; Sphacelotheca holwayi G. P. Clinton & Zundel, in Zundel, Mycologia 22: 143. 1930. Type. On Andropogon bicomis L. Bolivia. Sur Yungas: Villa Aspiazu, 31 May 1920, W E. D. & M. M. Holway 686 (type, BPI 177 831!).
Sori in dense groups in congested partial inflorescences (witches' brooms); long cylindiical, curved; 0.7 x 15-40 mm; a single sorus has one central columella of host tissue and an early rupturing peridium. Spore mass powdery; dark. Spores single; spherical to globose or cylindrical; (9-)10-12(-13) x (1 1-)12-15(-17) m m; dark reddish-brown. Wall 1.5-2.5 m m; with gern pore and ca 0.5 m m high warts; seen by SEM, with srialler warts between the large ones. Germination with phragmobasidia developing basidiospores.
In Panama on Andropogon bicornis (Poaceae):
Chiriquí: Road from Gualaca to Chiriqui Grande, alt. ca. 650 m, 22 Aug 1998, Piepenbring, Arrocha, Caballero and Cáceres 2 385 (PMA).
Coclé: La Pintada, Corregimiento Harino, El Copé, 2-4 km N of La Chata, alt.ca. 400 m, 22 Sep 1994, Piepenbring & Lorenzo 1 271 (PMA, USJ 44 251); La Pintada, Corregimiento Harino, El Copé, close to El Alto, Aserradero, alt. ca. 780 m, 23 Sep 1994, Piepenbring & Domínguez 1 282 (PMA); El Copé, from El Copé to Alto Calvario 5-7.2 km, alt. ca. 800-1 100 m, 13 Feb 1995, Piepenbring and Domínguez 1 451 -1 453 (1 451-1 453 PMA, 1 452 USJ 53 533); El Copé, 1 km N of Alto Calvario, alt. ca. 1 160 m, 13 Feb 1995, Piepenbring & Domínguez 1 454 (PMA); La Pintada, 4-5 km behind the entrance of Coclecito on the way to Bolteadero, alt. ca. 500-600 m, 14 Feb 1995, Piepenbring and Domínguez 1 458-1459 (PMA); Valle de Antón, alt. ca. 660 m, 15 Feb 1995, Piepenbring & Domínguez
1 470 (PMA).
Colón: Ciudad Colón, Santa Rita Arriba, alt. ca. 260 m, 15 Feb 1995, Piepenbring &Domínguez 1 473 (PMA).
Panamá: 2.6 km N from the Interamerican highway direction Cerro Campana, alt. ca. 450 m, 23 Feb 1995, Piepenbring 1 493 (PMA); Chicá, 4.4 km N of INRENARE direction Cerro Campana, alt. ca. 800 m, 23 Feb 1995, Piepenbring 1 494 (PMA).
8. Sporisorium ovapium (Griffiths) Vánky, Mycotaxon 65: 138. 1997; Sorosporiumovarium Griffiths, Bull. Torrey Bot. Club 34: 209. 1907. Lectotype (designated by Zundel 1953) en Urochloa reptans (L.) 0. Stapf in Prain [Brachiaria reptans (L.) C. Gardner and C. E. Hubb., Panicum caespitosum Sw.]. Mexico. Dublan, Hidalgo, 9 Sep 1905, Griffiths (type, BPI 180 088!).
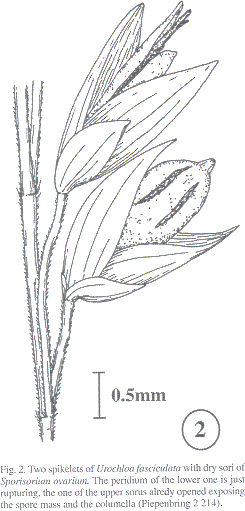
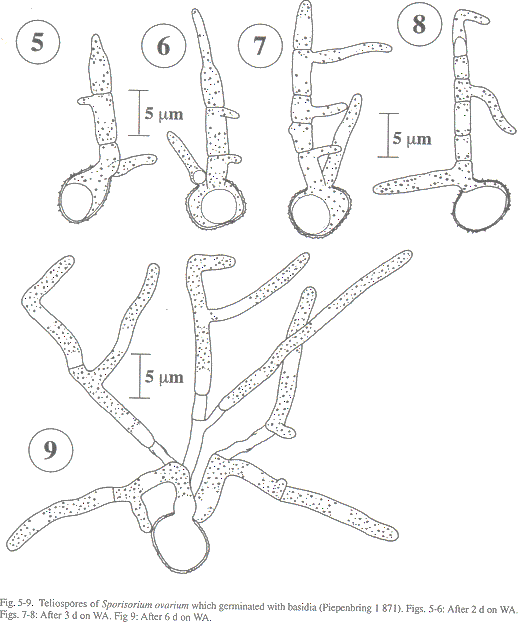
Sori usually in all the spikelets of an inflorescence (Fig. 1); globose to cylindrical; 3-5 mm long; a single sorus with one central columella of host tissue and a longitudinally rupturing peridium (Fig. 2), which opens in wet environment exposing the spore mass (Fig. 1, comp. Piepenbring et al. 1998). Spore mass mixed with sterile cells; powdery; dark brown. Spores in loose groups (Fig. 3); single spores globose or subangular; (8-)10-12 x 11-13 m m; light reddish-brown. Wall ea 0.8 m m; covered by ca 0.7 m m high warts; seen by SEM, with smaller warts between the large ones (Fig. 4). Germination with phragmobasidia of three to numerous cells forming hyphal branches without prior conjugation (Figs. 5-9).

In Panama on Urochloa fasciculata (Sw.) R. Webster [Brachiaria fasciculata (Sw.) Parodi]
(Poaceae):
Herrera: Parque Nacional Sarigua, alt. ca. 5 m, 30 Sep 1996, Piepenbring & Domínguez 2 214 (PMA).
Further material illustrated: Mexico. Yucatán: Sisal, 7 Oct 1995, Piepenbring 1 871 (XAL).
Sporisorium ovarium was hitherto only known from Mexico (see type; Durán 1987). This is its first observation in Central America.penbring 2 214). Fig. 3: Patl of a ball of teliospores. Scale bar: 10 pm. Fig. 4: Teliospores.

9. Trichocintractia utriculicola (Henn.) M. Piepenbr., Canad. J. Bot. 73: 1095. 1995; Cintractia leucoderma (Berk.) Henn. f. utriculicola Henn., Hedwigia 34: 336. 1895; Cintractia utriculicola (Henn.) G. P. Clinton, J. Mycol. 8: 143. 1902. Type. On Rhynchosporagigantea Link. Brazil. Santa Catharina: Blumenau, Möller 293 (type probably lost).
Sori in some spikelets of an infected inflorescence; globose to egg-shaped; 2-2.5 x 3-5 mm; with greyish white, persistent peridia which rupture with several lobes and forrn sacs. Spore mass dark brown; powdery; permeated by groups of long sterile fungal cells in the basal part. Spores single; spherical to globose; (9-)10-13 x (10-)11-15(-17) m m; light ochre-brown to dark brown. Wall 1-2.5 m m; two-layered; covered by fine warts. Germination with phragmobasidia; often twocelled parts of the basidia separate from the rest of the basidium, and plasma left in the teliospore germinates again; basidial cells develop basidiospores or hyphae.
In Panama on Rhynchospora corymbosa (L.) Britton [R. aurea Vahl] (Cyperaceae):
Bocas del Toro: 4 km SW of Chiriquí Grande, alt. ca. 5 m, 10 Feb 1995. Piepenbring 1 437 (material lost).
Colón: Ciudad Colón, Billa Catharina, alt. ca. 60 m, 16 Feb 1995, Piepenbring &Domínguez 1 475 (PMA, USJ 53 535); Ciudad Colón, Puerto Pilón, alt. ca. 20 m, 17 Feb 1995, Piepenbring and Domínguez 1 486 (PMA).
10. Ustilago hypodytes (Schltdl.) Fr., Syst. Mycol. 3: 518. 1832; Caeoma hypodytes Schltdl, FI. berol. 2: 129. 1824; Erysibe hypodytes (Schltdl.) Wallr., FI. crypt. Gern. 2: 216. 1833; Uredo hypodytes (Schltdl.) Desm., Ann. Sci. Nat. Bot. II 13: 182. 1840; Cintractia hypodytes (Schltdl.) Maire, Bull. Soc. Bot. France 53: 198. 1906. Lectotype (sel. by Hirschhorn, Farlowia 3: 74. 1947). On Elymus arenarius L. Germany. Near Berlin, "Meso Marchia", Oct 1884, P Sydow s. n. (lectotype ubi?, isolectotypes in Rabenh., Fungi. eur. No. 3201, e. g. in H.U.V., the personal herbarium of K. Vánky, Tübingen).
Sori surrounding the internodes of culms; starting just below the node and extending almost down to the next node; when young, hidden by sheaths of leaves; upper internodes and leaves usually stunted; infected plants sterile. Spore mass dark brown; semi-agglutinated to powdery. Spores single; mostly globose, subglobose to globose; 3.5-5.5 x 4-6(-7) m m; medium to dark olive-brown. Wall ca 0.5 m m; smooth. Gemination with phragmobasidia.
In Panama on Stipa setigera Presl, according to Zundel (1953).
11. Ustilago maydis (DC.) Corda, Icon. fung. 5: 3. 1842; Uredo maydis DC., FI. franç. 6: 77. 1815 ' Type. On Zea mays L. Europe (type ubi?).
Sori single to numerous in one plant; in vegetative and generative organs, e. g. partial inflorescences, kernels, or organs of male flowers; galls formed by hypertrophic, parenchymatic host tissue; sometimes more than 10 cm large; yellow-white to reddish; later rupturing. Spore mass powdery; dark brown. Spores single; spherical to globose; (9-)10-12(-13) m; light reddish-brown. Wall ca 1 m m; covered by ca 0.5 m m high warts, smaller warts between the high cone-shaped ones, as seen by SEM. Germination with three- to four-celled phragmobasidia; two or three cells of a basidium can separate from the rest of the basidium.
In Panama on Zea mays (cult.) (Poaceae):
Coclé: Corregimiento Harino, below El Copé, alt. ca. 190 m, 24 Sep 1994, Piepenbring & Domínguez 1 300 (PMA).
Ustilago maydis is reported for Pamana by Toler et al. (1959), being of minor economic importance. Young white galls can be eaten after cooking (called "huitlacoche" in nahuat, Mexico; e. g. Valverde et al. 1995).
12. Ustilago schroeteriana Henn., Hedwigia 35: 215. 1896. Type. On Paspalum sp. Brazil. Santa Catharina: near Itajahy, Sep 1887, Ule 1 615 (type, CH, n. v.).
Sori usually destroying all the spikelets of an inflorescence including tissue of the inflorescence axis; first covered by host tissue which flakes away. Spore mass powdery; dark brown. Spores single; spherical to globose; (11-) 1 5-17(-20) m m; light reddish-brown. Wall 1-1.5 m m; often thinner and lighter at one spot; covered by dense, 0.8 m m high warts; smaller warts between the large warts, as seen by SEM. Germination with phragmobasidia forming basidiospores on short sterigmata.
In Panama on Paspalum virgatum L. (Poaceae):
Chiriquí: Ciudad David, alt. ca. 200 m, 30 Aug 1998, Piepenbring & Cáceres 2 388 (PMA).
Reported for Panama by Zundel (1953) on Paspalum repens Bergius [P paniculatum Walter].
13. Ustilago scitaminea Syd., Ann. Mycol. 22: 281. 1924. Lectotype. On Saccharum officinarum (sel. by Vánky, Mycotaxon 41: 492. 1991). India. Bhagalpur, Bengal, 26 Aug 1907, Butler (lectotype ubi?; isolectotypes in Sydow, Ustilagineen 384, e. g. H.U.V 4 454, in the personal herbarium of K. Vánky, Tübingen).
Sorus as whip-shaped, elongated main axis; infected plants sterile; whip often more than 1 m long, the diameter decreasing from ca. 1 cm at the base to few millimeters in the distal part; young spore mass covered by host tissue which flakes away. Spore mass mixed with sterile cells; powdery; dark brown. Spores single; spherical to globose; (5-)6-7(-8) m m; reddish-brown. Wall ca. 0.8 m m; covered by few low warts; wall between the warts smooth, as seen by SEM. Sterile cells scattered between the spores; mostly in irregular groups of chains; larger than the spores; hyaline. Wall 1-1.5 m m, smooth. Germination with phragmobasidia; basidial cells often conjugase; dikaryotic cells grow with hyphae elaborating elongated dikaryotic blastoconidia.
In Panama on Saccharum sp. cult. (Poaceae):
Chiriquí: Distrito Alanje, Corregimiento Alanje, Central Azucarera de Alanje, 21 Mar 2000, L. de Castillo s. n. (Piepenbring 2 733, PMA).
Ustilago scitaminea can cause economic losses in sugar cane plantations, especially on susceptible cultivars. It is reported for Panama by Comstock et al. (1 983).
1.1.1.2 Mycosyringaceae
14. Mycosyrinx cissi (DC.) Beck, Ann. K. K. Naturhist. Hofmus. 9: 123. 1894; Uredocissi (cyssi) DC., in Poir., Encycl. meth. bot. 8: 228. 1808; Ustilago? cissi (DC.) Tul. and
C. Tul., Ann. Sci. Nat. Bot. III 7: 92. 1847. Schroeteria cissi De Toni, in Sacc., Syll. fung. 7: 501. 1888; Geminella exotica var. de candollei A. A. Fischer v. Waldh. (nomen novum illegit.), Aperçu syst. Ustilag., p. 43. 1877. Type. On Cissus sicyoides L. Dominican Republic (Santo Domingo). Poiteaux s. n. (type ubi?).
Sori in witches' brooms with numerous branches; often more than 1 m long; sori 1-2 x 5-30 mm; tubes of host tissue enclose the spore mass; old tubes rupture laterally disclosing the spores. Spore mass dark; powdery. Spores in firmly united pairs; the two outer halves of the cells semi-spherical, the halves directed towards the contact point fiat cone-shaped; one spore seen from the side 7-8(-9) x 12-13(-14) m m; dark olive-brown. Wall ca 1.5 m m; on the spherical half finely granular, on the conical side smooth. Germination with basidiospores successively developing directly from the teliospore; basidiospores folded and partly thickened; probably up to four basidiospores originate from one spore cell.
In Panama on Cissus sicyoides(Vitaceae):
Colón: Portobelo, Nuevo Torosí, on the way to More, Río Sucio, alt. ca. 10 m, 17 Feb 1995, Piepenbring, Domínguez & Girón 1 488 (PMA, USJ 53 538).
Darién: Río Cocalito, on river island, 7 Feb 1982, Whitefoord & Eddy 67 (on PMA 35 383, PMA).
Reported for Panama by Zundel (1953).
1.1.2 Urocystales
1.1.2.1 Doassansiopsaceae
15. Doassansiopsis limnocharidis(Cif.) Vánky, Mycotaxon 43: 424. 1992; Doassansia limnocharidis Cif., Ark. Bot. 23(A): 23. 1931. Type. On Limnocharis flava (L.) Buchenau. Dominican Republic. La Vega: Cordillera Central, Bonao, Aug 1929, Ciferri2 529 (type ubi?).
Sori as thickened leaf spots; often in rows parallel to the midrib; round to ovoid; ca 15 mm diam; when dry, brown; when old, breaking out of the leaf. Spore balls mostly in the air canals of the mesophyll; globose; flattened at contact to adjacent balls; 100-200 m m; firmly composed of central sterile pseudoparenchyma and a layer of fertile cells covered by a layer of sterile cortical cells. Fertile cells (teliospores) globose; their longest axis 11 - 14 m m; yellowish. Wall slightly thicker than the wall of sterile cells. Centralcells very irregularly isodiametric. Cortical cells radially flattened; often collapsed. Germination of fertile cells with holobasidia carrying apically numerous filiform basidiospores; basidiospores develop yeast-like cells or hyphae.
In Panama on Limnocharis flava (Limnocharitaceac):
Bocas del Toro: Chiriquí Grande, close to the ferry station, alt. ca. 2 m, 11 Feb 1995, Piepenbring 1 438 (PMA, USJ 53 532).
1.2 Exobasidiomycetidae
1.2.1 Tilletiales
1.2.1.1 Tilletiaecae
16. Tilletia ayresii Berk., in Massee, Buil. Misc. Inform. 1899: 146. 1899; Conidiosporomyces ayresii (Berk.) Vánky, in Vánky & R. Bauer, Mycotaxon 43: 429. 1992. Type on Panicum maximum Jacq. Mauritius. Mountains above Port Louis, Ayres 4 754, in Herb. Berkeley (type, K, n. v.).
Sori in several to numerous flowers of an inflorescence; sack-shaped galls formed by ovary tissue; when young, closed; early rupturing apically with stellate lobes; ca 3 x 3-5 mm;
apically gray, at the base reddish. Spore mass powdery; olive; composed of spores, single sterile cells, and groups of conidia. Spores single; spherical; (12-)14-15(-16) µm; brown. Wall 2.5-3.5 µm; covered by up to 2 µm high warts. Sterile single cells spherical to globose; (9-)11-16(-19) x (10-)12-18(-21) µm; colorless. Wall 2 µm; covered by 1 µm high, dense warts. Transitonal forms between single sterile cells and spores are present. Groups of conidia globose; up to 40 x 70 µm. Conidia loosely joined; mostly "y" -shaped; up to 20 µm long; hyaline; wall thin; smooth. Germination of the spores after severas days; with holobasidia apically carrying numerous filiform basidiospores. Germination of conidia within one day; with hyphae, ballisto-, and blastoconidia.
In Panama on Panicum maximum (Poaceae):
Chiriquí: David, alt. ca. 50 m, 6 Feb 1995, Piepenbring 1 390 (PMA); Boquete, Volcancito, 12 Mar 2000, Piepenbring, Cáceres and students 2 717 (PMA).
Coclé: La Pintada, Corregimiento Harino, El Copé, La Chata, alt. ca. 390 m, 22 Sep 1994,
Piepenbring & Lorenzo 1 273 (PMA); Penonomé, El Chorrillo, alt. ca 70 m, 29 Sep 1996, Piepenbring & Domínguez 2 210 (PMA).
Colón: Ciudad Colón, Billa Catharina, alt. ca. 60 m, 16 Feb 1995, Piepenbring & Domínguez 1 479 (PMA, USJ 53 537).
Herrera: Road to the Parque Nacional Sarigua, alt. ca. 15 m, 30 Sep 1996, Piepenbring & Domínguez 2 218 (PMA).
Los Santos: Las Tablas, alt. ca. 15 m, 1 Oct 1996, Piepenbring & Domínguez 2 220 (PMA).
Panamá: Jardín Botánico Summit, alt. ca. 80 m, 15 Sep 1994, Piepenbring 1 229 (PMA); 1.5 km NW Paraíso, alt. ca. 70 m, 17 Sep 1994, Piepenbring 1 232 (PMA).
On Setaria paniculifera (Steudel) Fourn. (Poaceae):
Coclé: La Pintada, Corregimiento Harino, El Copé, La Chata, 22 Sep 1994, Piepenbring &Lorenzo 1 275 (sparse material only in USJ 44 245).
17. Tilleda barclayana (Bref.) Sacc. & Syd., in Sacc., Syll. fung. 14: 422. 1899; Neovossiabarclayana Bref., Unters. Gesammtgeb. Mykol. 12: 170. 1895. Type on Pennisetum triflorum Nees. India. Himalaya, Simla, Barclay s. n. (type ubi?).
Tilletia horrida Tak., Bot. Mag. Tokyo 10: 20. 1896; Neovossia horrida (Tak.) Padw. and
A. Khan, Imp. Mycol. Inst. Mycol. Papers 10: 2. 1944. Type on Oryza sativa. Japan. Kyoto, 1895 (type ubi?). (Synonym according to Tullis & Johnson 1952)
Sori in some to all the ovaries of an inflorescence; concealed by glumes. Spore mass powdery; black. Spores single; globose; ca (20)23-32 µm; dark olive brown. Wall covered by spines, ca 2.5-4 µm high; covered by hyaline sheath. Germination with holobasidia carrying numerous filiform basidiospores.
In Panama on Oryza saliva L. (Poaceae), according to Toler et al. (1959).
This species, called kernel smut or anublo del grano, is reported for Panama by Cralley (1954, cited in Toler et al. 1959) as Neovossia barclayana. Toler et al. (1959) and Panamanian phytopathologists consulted by the author consider it to be of minor economic importance in Panama. However, as rice from areas with infection by T. barclayana is imported to Panama, constant survey ¡s necessary to detect infection as early as possible.
The delimitation of the genus Neovossia from the genus Tilletia is difficult, because the sets of character states of the different species show a morphologic continuum. Until further data are available, species of Neovossia other than the type species of the genus, N. moliniae Körn., are better cited in the genus Tilletia.
1.2.2 Georgefischeriales
1.2.2.1 Eballistraceae
18. Ebailistra oryzae (Syd. and R Syd.) Bauer et al., Mycol. Res. in press. Entylomaoryzae Syd. & P Syd., Ann. Mycol. 12: 197. 1914. Type. On Oryza sativa L. Philippines.
Laguna: Los Baños, 10 Dec 1913, Raimundo 2 202 (last number almost iIlegible), Baker
2 203 (holotype, B Pl 176 14 1!).
Sori as spots on leaf blades and sheaths; numerous; 0.2-0.3 mm x 0.5-2 (-4) mm and larger by fusion; linear-rectangular by adjacent veins; on both sides of the leaf lead-colored; slightly raised; covered by the epidermis. Spore mass firmly agglutinated; dark. Spores adhere in irregular masses; mostly polyhedral by compression, subglobose, or flattened; (5) 7-9 (-10) x (6-) 8-11 µm; light olive-greyish. Wall 0.5-1 µm; smooth.
In Panama on Oryza sativa (Poaceae) according to Cralley (1954, cited in Toler et al. 1959), not affecting yields.
1.2.3 Entylomatales
1.2.3.1 Entylomataceae
19. Entyloma bidentis Henn., in Engl., Pflanzenw. Ost-Afrikas C, p. 49. 1895. Type. On Bidens pilosa L. East Africa: Marangu, near the Kilimanjaro, Tanganyika Terr., 1 580 m, Volk 2 283 (type ubi?).
Sori as leaf spots; few to several on a single leaf; circular; sometimes convex but not thickened; diam 1-2(-3) mm, sometimes larger by fusion; when fresh, yellow-greenish, sometimes with violet centre; when old, brown. Spores single; densely packed between the host cells; spherical to globose, sometimes subangular; (9-)11-13(-14) x (12-)13-15(-18) µm; hyaline or yellowish-brown. Wall two-layered; the inner layer 1 µm, the outer 1-2 µm; smooth. Germination with holobasidia carrying apically few sessile basidiospores; basidiospores and basidial cell conjugate and develop sickle-shaped and filiform conidia.
In Panama on Bidens pilosa (Asteraceae):
Chiriquí: Road from Gualaca to Chiriquí Grande, alt. ca 840 m, 22 Aug 1998, Piepenbring,Arrocha, Caballero & Cáceres 2 383 (PMA).
20. Entyloma compositarum Farlow, Bot. Gaz. 8: 275. 1883. Type. On Aster puniceus L. USA. Massachusetts: White Montains, Woods' Hole, Sep 1881, Trelease 1 085 (holotype BPI 195048!).
Entyloma costaricense Cif, Nuovo Giorn. Bot. Ital. 40: 259. 1933. Type. On Viguiera sylvatica Klatt. Costa Rica. Alajuela: Grecia, 13 Jan 1925, H. Sydow 383 (lectotype, M!; isolectotypes BPI 175119! and BPI 175120!; designated here).
Sori as leaf spots; numerous on a single leaf, angular by adjacent veins; diam 1-2(-2.5) mm; when fresh, whitish; when dry, adaxially green (almost invisible) and abaxially whitish-green. Spores single; not densely packed between the host cells; spherical to globose; (7) 8- 10(-11) x (7-)9-11(-12) µm; hyaline. Wall, as seen by LM, mostly one-layered; ca 1 µm; smooth. Germination with holobasidia carrying apically few sessile basidiospores; basidiospores and basidial cell conjugate and develop hyphae with retraction septae.
In Panama on Viguiera sp. (Asteraceae):
Chiriquí: Road from Gualaca to Chiriqui Grande, alt. ca. 840 m, 22 Aug 1998, Piepenbring, Arrocha, Caballero & Cáceres 2 384 (PMA).
Entyloma costaricense is a synonym of E. compositarum because the size and the shape of the sori as well as characteristics of the teliospores are similar in both species (comp. Sydow 1926, Savile 1947).
21. Entyloma dahliae Syd. and P. Syd., Ann. Mycol. 10: 36. 1912. Type. On Dahlia variabilis Desf. (cult.). South Africa. Natal, Harden Heights, 11 Apr 1911, Pole-Evans s. n. (type, S!).
Sori as leaf spots; few to several on a single leaf, subcircular; limits mostly fading, rarely angular by adjacent veins; diam ca 10 mm, variable depending on the age and degree of fusion with adjacent spots; when fresh, adaxially light green and abaxially whitishgreen; when old, brown to whitish-ochre, abaxially sometimes with whitish centre. Spores single; densely packed between the host cells; spherical to globose or irregularly shaped, subangular by contact to other spores; (11-)12-14(-16) x (11-)13-16(-17) µm; light yellow to brown. Wall two-layered; inner layer 0.7 µm; the outer up to 3.5 µm; smooth. Germination with holobasidia carrying apically few sessile basidiospores; basidiospores and basidial cell conjugate and develop sickle-shaped and filiform conidia.
In Panama on Dahlia sp. cult. (Asteraceae):
Chiriquí: Bajo Boquete, alt. ca. 1 500 m, 23 Aug 1998, Piepenbring, Cáceres, Cueva &Esquivel 2 386 (PMA).
22. Entyloma spilanthis Speg., Anales Mus. Nac. Hist. Nat. Buenos Aires 12: 292. 1909. Type. On Spilanthes leptophylla DC. [S. arnicoides DC.]. Argentina. Ensenada, La Plata, 28 Oct 1906 (type, LPS 3 351!, no sori could be found).
Entyloma ecuadorense Syd., Ann. Mycol. 37: 328. 1939. Type. On Acmella oppositifolia (Lam.) R. K. Jansen var. oppositifolia [Spilanthes americana (Mutis) Hieron.]. Ecuador Tungurahua: Hacienda San Antonio, near Baños, 1 Dec 1937, H. Sydow 410 (lectotype, H.U.V. 819!, designated by Piepenbring 1996a: 66); 4 Dec 1937, H. Sydow 435 (paratype).
Sori as leaf spots (Fig. 10); irregularly circular or angular by adjacent veins; diam 1-4 mm; when fresh, light yehow-green to brownish; when dry, ochre. Spores (Fig. 11) single; densely packed between the host cells; spherical, globose, or subangular; (9-)10-13(-16) µm; colorless, sometimes yellowish brown. Wall two-layered; inner layer ca 0.7 µm; the outer up to 2 µm; smooth. Germination with holobasidia carrying apically few sessile basidiospores; basidiospores and basidial cell conjugase and develop hyphae.
In Panama on Acmella papposa (Hemsl.) R. K. Jansen var. macrophylla (Greenm.) R.
K. Jansen (Asteraceae):
Chiriquí: Boquete, Alto Chiquero, alt. ca. 2 000 m, 7 Feb 1995, Piepenbring and Cueva1 397 (PMA, USJ 53 529); Boquete, Alto Quiel, 12 Mar 2000, Piepenbring, Cáceres and students 2 716 (PMA).
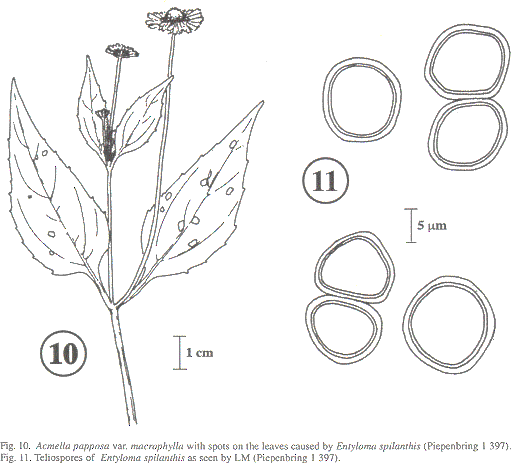
Entyloma spilanthis is known on Acmella oppositifolia var. oppositifolia from Costa Rica (Piepenbring 1996a) as E. ecuadorense. Soral morphology and the size of the spores [(9)10-12 x (10-)11-13 µm] of the infection found on A. papposa var. macrophylla are very similar to those on A. oppositifolia var. oppositifolia. Acmella papposa var. macrophylla is therefore a new host species.
Sydow described E. ecuadorense as a new species differing from E. spilanthis Speg. by larger sori, slightly smaller spores (E. spilanthis: 11-14(-16) µm, E. ecuadorense: 11-14 µm), and a hyaline, ca 1 µm thick spore wall. According to him, the sori of E. spilanthis are very small and difficult to detect on herbarium material, the walls of its spores are yellow-brown and 1.5-1.8 µm thick. The author could not check soral morphology or the size of the spores of the type of E. spilanthis without destroying too much of the type specimen.
Material collected by the author in Costa Rica and Panama corresponds to E. ecuadorense by relatively large, conspicuous spots on the leaves and to E. spilanthis by the size of the spores and lhe thickness of the spore walls. The color of the spore walls varies from hyaline to slightly yellowish and definitely yellowish brown. It seems to be impossible to distinguish E. ecuadorense from E. spilanthis, so they are considered synonyms.
2 Urediniomycetes
2.1 Microbotryales
2.1.1 Microbotryaceae
23. Sphacelotheca cf. hydropiperis (Schumach.) de Bary, Vergl. Morph. Biol. Pilze, p. 187. 1884; Uredo hydropiperis Schumach., Enum. pl. Saell. 2: 234. 1803; Ustilago hydropiperis (Schumach.) J. Schröter, in Cohn, Beitr. Biol. Pflanzen 2: 355. 1877. Type. On Polygonum hydropiper L. Denmark, Sjaelland (type ubi?).
Sori in some to numerous flowers of an inflorescence; a single sorus with one central columella and a peridium, both made of fungal cells; columellae ca 4-7 mm long; peridia dirty white to brownish, rupturing with 3-4 lobes. Spore mass powdery; brown-violet. Spores single or in chains; spherical to globose; 8- 10 x 10-14(-18) µm; reddish-brown; with 2(-3) short appendices, mostly on opposite sides of a spore. Wall ca 1.5 µm; covered by dense, fine, 0.5 µm high warts, which can fuse forming a more or less complete reticulation, visible only by SEM. Germination with phragmobasidia with sessile basidiospores, which can conjugate.
In Panama on Polygonum punctatum Elliott (Polygonaceae):
Chiriquí: Boquete, Bajo Chorro, alt. ca. 1 850 m, 8 Feb 1995, Piepenbring and Cueva
1 400 (PMA, USJ 53 530); Bajo Boquete, alt. ca. 1 500 m, 23 Aug 1998, Piepenbring, Cáceres, Cueva & Esquivel 2 387 (PMA).
On Polygonum persicarioides Humb., Bonpl. and Kunth:
Close to Baños, Hacienda San Antonio, 10 Dec 1937, H. Sydow; Sydow, Fungi exotici exsiccati 1 043 ("Sphacelotheca hydro piperis", M!).
According to Vánky & Oberwinkler (1994), these specimens should be called Sphacelotheca koordersiana (Bref.) Zundel because they differ from typical S. hydropiperis by having sori in only some flowers of an inflorescence and warts on the spores mostly united in rows. Further investigation is necessary to ascertain whether these differences are enough to merit an additional species.
2.1.2 Ustilentylomataceae
Sori in groups of spikelets of an inflorescence; as swellings of the pedicel in pisfillate spikelets and as swollen rhachillae in staminate spikelets; when young, green, when old, orange colored; in old sori, vascular strands are visible as ramified threads. Spore mass crumbly; orange. Spores in irregular, subglobose to elongated groups, 30-70 x 50-110 µm, or single; a single teliospore mostly subglobose or blundy angled; (8-)10-12(-13) x (10-)13-17(-19) µm; young light yellow; old orange colored. Wall ca 2 µm thick; in LM two-layered; in SEM with an írregularly folded surface. Germination with phragmobasidia with sessile basidiospores.
In Panama on Scleria melaleuca (Cyperaceae):
Panamá: N of Ipetí, alt. ca. 110 m, 19 Feb 1995, Piepenbring & Domínguez 1 490 (PMA, USJ 53 539).
Discussion
This first account of smut fungi in Panama includes four species based only on literatura and 20 species collected by the author. Among the latter are 14 new records for Panama, constituing 6 1 % of the total of 24 species cited here. This high percentage is not surprising, because hitherto nobody had studied the diversity of Panamanian smut fungi.
Most wild host species of smuts grow in rural plant communities, mostly at roadsides and close to human settlements. This was also the case in Costa Rica (Piepenbring 1996b).
Tilletia ayresii, the most often collected smut species in Costa Rica, is also one of the most common smut species in Panama. Sporisorium holwayi, however, was only rarely collected in Costa Rica (Piepenbring 1996a), while it was often collected at the pacific slopes of the Panamanian Cordillera.
In comparison with the lists of smut fungi for other neotropical countries (comp. introduction) the list of the species known for Panama is short and includes only four species which are not also known for Costa Rica (Table 1). Sphaceloteca panamensis is known only for Panama. The small size of the checklist of Panamanian smut fungi is mainly due to the very short time spent for the investigation of this group in Panama. The high similarity with the smut diversity in Costa Rica and the lack of new species in the present publication may be caused by the fact that the author finds species already known to her more easily than new ones. lt may also be related to the fact that immigration of terrestrial species to Panainanian grounds was limited by the ocean covering most of it until the latter Tertiary (ca. 3.5 mio. yrs ago), when the mesoainerican land-bridge closed in the area of Panama (Stanley 1994). The high diversity of its present flora, however, is the best guarantee for interesting results which will be obtained by a more thorough investigation on Panamanian smut fungi.


Acknowledgements
Q. Domínguez and further members of the INRENARE (now ANAM) and P. Caballero, C. Arrocha, and 0. Cáceres from the UNACHI (Universidad Autónoma de Chiriqui) are thanked for help with organization and company during the field work in Panama. M. Correa supported this study as director of the herbarium PMA. In Germany, Y Uhle-Schneider is thanked for mounting the drawings, H. Schoppmann and F. Albrecht for technical assistance. R. Cropper is thanked for revising the English and S. T. Garnica for revising the Spanish of the manuscript. Curators of cited herbaria are thanked for loans of specimens. The DAAD (Deutscher Akademischer Austauschdienst) is thanked for financial support.
Resumen
Esta es la primera publicación dedicada a la diversidad de carbones en Panamá. Tiene su base en trabajo de campo, estudio de especímenes herborizados y referencias de la literatura. Se incluyen carbones patógenos de plantas cultivadas y silvestres. Las últimas se encontraron sobre todo en zonas rurales. Entre las 24 especies citadas en este estudio, 14 especies son primeros registros para Panamá y una de éstas, Sporisorium ovarium, para América Central. Se encontró Entyloma spilanthis por primera vez en la planta hospedera Acmella papposa var.macrophylla (Asteraceac). Entyloma costaricense y Entyloma ecuadorense son sinónimos de Entyloma compositarum y Entyloma spilanthis respectivamente. "Sphacelotheca" panamensis es una especie dudosa. Se complementan las descripciones de las especies con algunas ilustraciones, una lista de especies y una clave.
Note
The type of "Sphaceloteca" panamensis was recendy located in BPI. It differs from other species known on Cymbopongon and related genera of Poaceae by the precense of sori in spikelets and relatively large teliospores with several germ areas each. lt belongs to the genus Sporisorium because of columellae and fungal peridia in the sori.
Sporisorium panamensis (Zundel & Dulap) M.
Piepenbr., comb. nov.
Basionym: Sphaccioreca panamensis Zundel and Dulap, in Zundel, N. Amer. fi. 7: 995. 1939. Type. On Cymbopogon citratus (DC.) Stapf. Panamá. Chiriquí, Hitchcock 8295 (holotype, BPI 190260!).
References
Bauer, R., F. Oberwinkler & K. Vánky. 1997. Ultrastructural markers and systematics in smut fungi and allied taxa. Canad. J. Bot. 75: 1 273-1 314. [ Links ]
Bauer, R., D. Begerow, F. Oberwinkler, M. Piepenbring & M. L. Berbee. 2001. Ustilaginomycetes, p. 57-83. In D. McLaughlin, E. McLaughlin and Lemke (eds.). Mycota VII. Systematics and Evolution. Springer, Berlin. [ Links ]
Comstock, J.C., S.A. Ferreira & T.L. Tew. 1983. Hawaii's approach to control of sugarcane smut. Plant Dis. 67: 452-457. [ Links ]
Davidse, G. & R.W. Pohl. 1994. 161. Cymbopogon Sprengel, p. 390-391. In: G. Davidse, M. Sousa S., & A. 0. Chater (eds.). Flora Mesoaniericana, Vol. 6, Alismataceae a Cyperaceae. Universidad Nacional Autónoma de México, México. [ Links ]
Dennis, R.W.G. 1970. Fungus flora of Venezuela and adjacent countries. Kew Bull. Add. Ser. 3: 1-531. [ Links ]
Durán, R. 1987. Ustilaginales of Mexico. Taxonomy, symptomatology, spore germination, and basidial cytology. Washington State University, Pullman, Washington. 331 p. [ Links ]
Langdon, R.F.N. & R.A. Fullerton. 1978. The genus Sphacelotheca (Ustilaginales): criteria for its delimitation and the consequences thereof. Mycotaxon 6: 421-456. [ Links ]
Ling, L. 1950. Studies in die genus Cintractia. II. Cintractia axicola and related species. Mycologia 42: 646-653. [ Links ]
Molina-Valero, L.A. 1980. Los hongos de Colombia - VI. Reconocimiento e identificación de Ustilaginales en Colombia. Caldasia 13: 49-96. [ Links ]
Piepenbring, M. 1996a. Smut fungi (Ustilaginales and Tilletiales) in Costa Rica. Beih. Nova Hedwigia 113: 1-155. [ Links ]
Piepenbring, M. 1996h. Ecology, seasonal variation, and altitudinal distribution of Costa Rican smut fungi (Basidiomycetes: Ustilaginales and Tilletiales). Rev. Biol. Trop. 44 (Suppl. 4): 115-123. [ Links ]
Piepenbring, M. 1996c. Two new , Entyloma species (Ustilaginales) in Central America. Sydowia 48: 241-249. [ Links ]
Piepenbring, M.1999. New and poorly known smut fungi in Cuba. Mycol. Res.103: 459-467. [ Links ]
Piepenbring, M. 2000. The species of Cintractia s. 1. (Ustilaginales. Basidiomycota). Nova Hedwigia 70: 289-372. [ Links ]
Piepenbring, M. & M. Rodríguez. 1998a. Carbones (Fungi: Ustilaginomycetes) de Cuba. I parte. Rev. Jard. Bot. Nac. (Cuba) 19: 121-131. [ Links ]
Piepenbring, M. & M. Rodríguez. 1998b. Carbones (Fungi: Ustilaginomycetes) de Cuba. Il parte. Revista Jard. Bot. Nac. (Cuba) 19: 132-146. [ Links ]
Piepenbring, M., G. Hagedom and F. Oberwinkler. 1998. Spore liberation and dispersal in smut fungi. Bot. Acta 111: 444-460. [ Links ]
Piepenbring, M., D. Begerow & F. Oberwinkler. 1999. Molecular sequence data assess the value of morphological characteristics for a phylogenetic classification of species of Cintractia. Mycologia 91: 485-498. [ Links ]
Savile, D.B.O. 1947. A study of the species of Entyloma on North American composites. Canad. J. Res., C, 25: 105-120. [ Links ]
Stanley, S.M. 1994. Historische Geologie. Spektrum Akademischer, Heidelberg. 632 p. [ Links ]
Sydow, H. 1926. Fungi in itinere costaricensi collecti. Ann. Mycol. 24: 283-426. [ Links ]
Toler, R.W., R. Cuellar and J.B. Ferrer. 1959. Preliminary survey of plant diseases in the Republic of Panama, 1955-1958. PI. Dis. Reporter 43: 1 201-1 203. [ Links ]
Tuilis, E.C. & A.G. Johnson. 1952. Synonymy of Tilletia horrida and Neovossia barclayana. Mycologia 44: 773-788. [ Links ]
Valverde M.E., 0. Paredes-López, J.K. Pataky & F. GuevaraLara. 1995. Huitiacoche (Ustilago maydis) as a food source - biology, composition, and production. CRC Crit. Rev. Food Sci. Nutr. 35: 191-229. [ Links ]
Vánky, K. 1987. Illustrated genera of smut fungi. Gustav Fischer, Stuttgart. 159 p. [ Links ]
Vánky, K. & F. Oberwinkler. 1994. Ustilaginales on Polygonaceae- a taxonomic revision. Beih. Nova Hedwigia 107:1-96. [ Links ]
Zundel, G.L.1939. Additions and coffections to Ustilaginales. North Amer. Flora 7: 971-1045. [ Links ]
Zundel, G.L. 1953. The Ustilaginales of the world. Pennsylvania State Coll. School Agric. Dept. Bot. Contr. 176: I-XI, 1-410. [ Links ]
1 Lehrstuhl Spezielle Botanik/Mykologie, Botanisches Institut, Universität Tübingen, Auf der Morgenstelle 1, 72076 Tübingen, Germany. Fax: 0049 7071 295344
E-mail: meike.piepenbring@uni-tuebingen.de

