











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los ríos de montaña son ecosistemas con alta diversidad de especies y, por ende, estratégicos desde el punto de vista ambiental (Meneses-Campo et al., 2019). En las últimas décadas, los ríos andinos se han visto sometidos a fuertes presiones antrópicas (Meza-Salazar et al., 2020), asociadas con la contaminación del agua, sobreexplotación de los recursos y expansión de la frontera agropecuaria que causan degradación del hábitat y provocan efectos negativos sobre la biota (Milán-Valoyes et al., 2011).
El establecimiento y distribución de las comunidades acuáticas en los ríos está determinada principalmente por las condiciones físicas y químicas del hábitat, así como por la oferta de recursos (Townsend et al., 2003). La flexibilidad en los requerimientos de dichas condiciones determinará la sensibilidad de cada organismo frente a la alteración del hábitat y, por ende, la estructura de la comunidad. En el caso de los ríos, las unidades geomorfológicas del cauce (rápidos, rizos y pozas) son la escala más recomendable para evaluar dichas alteraciones, ya que difieren en su morfología, profundidad, velocidad y tipos de sustrato y, además, presentan interacciones entre el caudal, la rugosidad del lecho y la pendiente (Ríos-Pulgarín et al., 2022). Parasiewicz et al. (2009) señalan, por ejemplo, que los rápidos son mesohábitats que se caracterizan por una velocidad alta, sustratos más gruesos y turbulencia en la superficie, mientras que los rizos son tramos poco profundos, con velocidad moderada y algo de turbulencia. Por el contrario, las pozas se caracterizan como tramos de agua relativamente profundos, debido a procesos hidráulicos de socavamiento o embalsada por la obstrucción parcial del canal, con menor velocidad de flujo y disminución de la concentración de oxígeno.
Entre los organismos que habitan los mesohábitats descritos anteriormente se encuentran los macroinvertebrados, los cuales forman una de las comunidades más ampliamente estudiadas en corrientes de montaña, debido a su alto grado de sensibilidad frente a las variaciones de las condiciones ambientales que ocurren continuamente en los ecosistemas. Esta comunidad agrupa numerosas especies de diferentes grupos taxonómicos, en su mayoría artrópodos, y constituye el segundo eslabón en las redes tróficas de los ríos de montaña y frecuentemente es empleada como indicadora de la calidad del agua (Roldán-Pérez & Ramírez-Restrepo, 2022; Suter & Cormier, 2015). También, se ha encontrado que estos organismos presentan una estrecha dependencia con la disponibilidad de hábitat y los recursos alimenticios, lo que los hace muy importantes desde el punto de vista funcional en el sistema fluvial (Elosegi & Sabater, 2009).
Dada su movilidad restringida y su pequeño tamaño, que oscila entre 0.5 mm hasta unos pocos centímetros, el concepto de hábitat para esta comunidad debe considerarse en la escala de mesohábitat (Roldán-Pérez & Ramírez-Restrepo, 2022). Entendido este como una unidad geomorfológicamente homogénea en términos de profundidad, velocidad y tipo de sustrato (Parasiewicz, 2007). Los macroinvertebrados no solo responden a las condiciones de calidad de agua, también son afectados por los cambios en el régimen de flujo, que, a su vez, modifica la estabilidad del lecho de forma diferente en cada mesohábitat y en las diferentes épocas hidrológicas, más aún cuando se trata de ecosistemas sometidos a escenarios de regulación. De manera que, los cambios de profundidad, velocidad y arrastre de sedimentos afectan la variedad y estabilidad de mesohábitat favorables para el establecimiento de estos organismos (Motta-Díaz & Vimos-Lojano, 2020; Walteros-Rodríguez & Castaño-Rojas, 2020).
En términos metodológicos, el muestreo de macroinvertebrados por mesohábitat ofrece ventajas adicionales, ya que evita subestimar la riqueza local, tal como lo ratificaron Vimos-Lojano et al. (2020) en su estudio sobre la preferencia de microhábitat de macroinvertebrados altoandinos. Aún más, si dicho muestreo se sincroniza con periodos hidrológicos de aguas bajas, o en descenso, en los cuales se favorece la colonización. De manera que el esfuerzo de muestreo diferenciado por hábitat y el periodo hidrológico en el que se lleva a cabo la recolección son determinantes para la adecuada caracterización de esta comunidad.
Los macroinvertebrados han desarrollado diferentes adaptaciones específicas que les permite sobrevivir en un medio tan fluctuante como es el ecosistema fluvial, tales como estrategias alimenticias, muchas de ellas oportunistas, de acuerdo con el lugar donde se desarrollan (Elosegi & Sabater, 2009; Walteros-Rodríguez & Castaño-Rojas, 2020). Otro aspecto importante para el establecimiento de las comunidades de macroinvertebrados es el aporte de hojarasca que llega a la corriente de la vegetación de ribera, ya que, además de constituir una de las principales fuentes de materia y energía dentro del sistema acuático, también sirve como refugio y alimento para los macroinvertebrados, favoreciendo su establecimiento en diversos mesohábitats del ecosistema en función de la oferta de estos recursos (Salazar-Castellanos et al., 2020).
La subregión del Oriente antioqueño en los andes colombianos es un escenario adecuado para evaluar la relación entre los macroinvertebrados y los diferentes elementos del hábitat mencionados, debido a que presenta una topografía quebrada y gran riqueza hídrica (Corporación Autónoma Regional de las Cuencas de los Ríos Negro y Nare, 2011). En ese contexto, el presente estudio tiene el objetivo de determinar cuáles son las variables del hábitat que más influyen en el establecimiento de las comunidades de macroinvertebrados en algunas corrientes de montaña del Oriente antioqueño. Se plantea como hipótesis de investigación que las variables físicas (velocidad y profundidad) y la cobertura vegetal (vegetación de ribera) tienen igual o mayor importancia que las variables químicas del agua (pH, temperatura, oxígeno disuelto, saturación de oxígeno y conductividad eléctrica), en la distribución de estos organismos en las corrientes del Oriente antioqueño. Los resultados de este estudio aportarán información respecto al efecto de las diferentes características del hábitat sobre los diferentes taxones, lo cual es fundamental en los procesos de conservación de la diversidad y la funcionalidad fluvial, así como en la restauración de los ríos andinos degradados.
Materiales y métodos
Área de estudio: El Oriente antioqueño se ubica en al sureste del departamento de Antioquia, y se destaca por ser una región con amplia variedad de climas, gran diversidad natural, además de una gran riqueza hídrica, características que la convierten en una región estratégica para la construcción de proyectos de energía, agricultura, industrialización y urbanización, que amenazan los ecosistemas (Echeverri-Restrepo et al., 2017). El presente estudio se llevó a cabo en las corrientes del Oriente antioqueño en jurisdicción de los municipios San Luis, Cocorná, El Carmen de Viboral y Granada (Fig. 1). Esta región abarca zonas de vida (Biomas) que van desde bosque muy húmedo tropical (bmh-T), hasta bosque muy húmedo montano bajo (bmh-MB) (Holdridge, 1978). Su temperatura oscila entre 16 y 29 °C, con altitudes de 170 a 2 500 m.s.n.m., y precipitación entre 1 000 y 5 000 mm/año (Corporación Autónoma Regional de las Cuencas de los Ríos Negro y Nare, 2012). El régimen hidrológico es bimodal, con dos periodos secos (febrero y agosto) y dos periodos lluviosos en abril y octubre (Ríos-Pulgarín et al., 2016). Los sitios evaluados en el estudio se encontraron entre 800 y 1 600 m.s.n.m., en el piso térmico tropical, lo que las hace equivalentes para realizar el análisis.
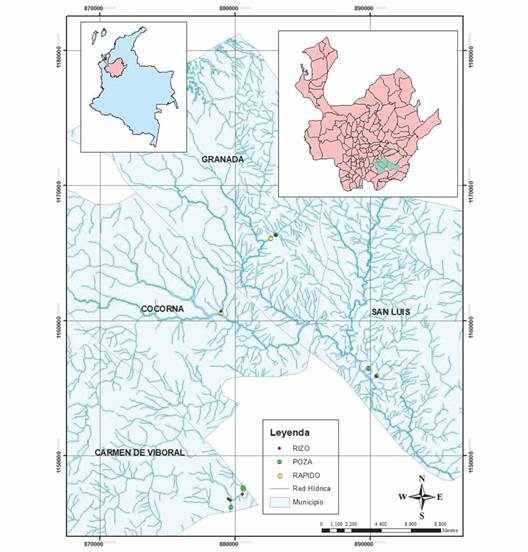
Fig. 1 Área de estudio y ubicación geográfica de los 10 sitios de muestreo en siete corrientes de montaña en los municipios de Granada, Cocorná, El Carmen de Viboral y San Luis. / Fig. 1. Area of study and geographic location of the 10 sampling sites in seven mountain streams in the municipalities of Granada, Cocorná, El Carmen de Viboral and San Luis.
Diseño de muestreo: Se establecieron 10 sitios de muestreo, distribuidos en las diferentes corrientes seleccionadas: quebrada Carbonera (S1), quebrada San Antonio (S2), quebrada La Calera (S3), quebrada La Isla (S5-S6), quebrada La Trinidad (S8), río Melcocho (S4-S7) y río Tafetanes (S9-S10). En cada corriente se seleccionó un tramo de aproximadamente 100 m, donde se identificaron todos los microhábitats presentes: rápido, rizo y poza (Parasiewicz, 2007). El mesohábitat rápido fue definido como aquellas zonas caracterizadas por profundidades menores con alta-velocidad y flujo-turbulento en la superficie, debido a la composición del sustrato que es más grueso que quiebra la superficie del agua. La turbulencia y el flujo de la corriente dan como resultado una concentración de oxígeno disuelto mayor. Los rizos fueron definidos por una menor profundidad, velocidad moderada y algo de turbulencia. La disposición de los sustratos hace que en la superficie del agua se formen ondulaciones. Las pozas se caracterizaron de acuerdo con la mayor profundidad, con una velocidad de flujo nula o baja, la composición del sustrato fino y la menor concentración de oxígeno. El muestreo fue realizado en época de transición (seco-lluvia) en febrero de 2021.
Recolección de muestras biológicas: En cada mesohábitat se midió un área de 20 m2. Se utilizó una red triangular (ojo de malla entre 0.5 y 1 mm), para la recolección de macroinvertebrados (N = 16), mediante varias técnicas de captura de acuerdo con el tamaño y el tipo de sustrato: a) en área de rocas, gravas y arenas ubicación de la red sobre el fondo en contracorriente y remoción manual del sustrato; b) en áreas con vegetación mediante barrido a lo largo del tramo; c) en áreas de pozas con depósito de hojarasca mediante resuspensión y recolección del material con la red, cada una de estas muestras se consideró una submuestra y se empleó para conformar una muestra integrada por mesohábitat. El esfuerzo de muestreo empleado en cada uno de los mesohábitats fue de 20 min.
El material de cada mesohábitat recolectado en la red triangular se almacenó en bolsas plásticas con cierre hermético y se cubrió con alcohol al 70 %. Simultáneamente, se realizó la recolección manual de macroinvertebrados levantando rocas y troncos al azar. Con una pinza se tomaron los organismos que se encontraron adheridos en estos sustratos y se depositaron en frascos PET de 30 mL con etanol al 70 %.
Métodos de laboratorio: Una vez separados los macroinvertebrados de la hojarasca se almacenaron en frascos PET de 30 mL debidamente rotulados. Para la observación e identificación de los macroinvertebrados se utilizó un estereomicroscopio WD NICON JAPAN modelo SMZ 1500 y claves especializadas de Álvarez-Arango y Daza-Ospina (2005), Aristizábal-García (2002), Domínguez y Fernández (2009), Manzo (2005), Posada-García y Roldán-Pérez (2003), con el fin de llegar al menor nivel taxonómico posible. Se depositaron 62 individuos de estas muestras en la colección de referencia de la Universidad Católica de Oriente (UCO), la cual está registrada ante el Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, con los códigos de colección de CMA UCO 3718 hasta CMA UCO 3738; actualmente publicados en línea a través del sistema de información sobre biodiversidad-SIB Colombia.
Parámetros fisicoquímicos: En cada sitio se hicieron mediciones de pH, temperatura (°C), oxígeno disuelto (mg/L), saturación de oxígeno (%) y conductividad eléctrica (µS/cm) con un equipo multiparamétrico HACH HQ40D. Los valores de turbidez se obtuvieron en el laboratorio de Monitoreo Ambiental de la UCO a partir de muestras tomadas en campo, con el turbidímetro de marca HACH 2100Q. Los métodos de campo y de laboratorio se estandarizaron de acuerdo con los protocolos definidos por Baird y Bridgewater (2017).
Parámetros de hábitat: Las características físicas del hábitat se tomaron de Montoya-Cardona (2022) y Rincón-Echeverri (2022), quienes realizaron estudios simultáneos en los mismos sitios de muestreo. Los parámetros estimados en estos estudios fueron: velocidad, profundidad, tipo de sustrato y cobertura vegetal. La velocidad (m/s) en cada uno de los mesohábitats se determinó con un correntómetro OTT C-31, la profundidad media (m) se estimó como promedio de tres medidas utilizando una mira topográfica. Para determinar el tipo de sustrato (granulometría), se midieron 100 rocas del lecho en cada mesohábitat, de acuerdo con el método de Wolman (1954). En los mesohábitats que no fue posible realizar medidas de sustrato se recolectó muestra para realizar una curva granulométrica del material del lecho a partir de la metodología propuesta por Arévalo-Mendoza et al. (2017).
La información de cobertura vegetal se obtuvo a partir de la aplicación del Índice de calidad de la vegetación de ribera Andina (QBR-And) (Acosta et al., 2008), que corresponde a una adaptación del índice QBR desarrollado para ríos mediterráneos por Munné et al. (2003). Dentro de este se incluyen cuatro métricas: a) grado de cubierta de la ribera, b) estructura de la cubierta, c) calidad de la cubierta y d) grado de naturalidad del canal fluvial. Además, este índice toma en cuenta las principales formaciones vegetales andinas y presenta intervalos de calidad y conservación de las riberas.
Tratamiento de datos: Se determinó la composición taxonómica por mesohábitat, así como la estructura de la comunidad de macroinvertebrados mediante el cálculo de la abundancia y de los números de Hill (q0: Riqueza total; q1-Equitatividad: número de especies comunes y q2-Dominancia: número de especies dominantes) (Hill, 1973), los cuales fueron calculados en el programa PAST versión 3.0 (Hammer et al., 2001).
Se realizó un análisis de varianza ANOVA, previa verificación de los supuestos del modelo, para establecer el efecto del factor mesohábitat sobre las variables ambientales, la abundancia y la diversidad de los macroinvertebrados. Para determinar las diferencias en la composición taxonómica por mesohábitat se empleó un análisis discriminante basado en la abundancia de los órdenes. Los puntajes de los vectores canónicos fueron utilizados para graficar en términos de varianza explicada, las funciones canónicas más relevantes, evidenciando el espacio canónico de diferenciación entre la abundancia de macroinvertebrados y sus hábitats de preferencia (Friendly, 2007). Complementariamente se realizó un análisis PCA para identificar aquellas familias que tenían la mayor contribución a la varianza entre mesohábitats. Con las familias seleccionadas, se realizó un análisis clúster (conglomerado), empleando el método de distancia euclidiana y se graficó un mapa de calor para visualizar patrones en la composición entre los mesohábitats, usando colores para representar la intensidad de la relación (Guisande et al., 2010).
La preferencia de los órdenes de macroinvertebrados por los diferentes mesohábitats en función de las variables ambientales, se verificó a partir de un análisis de escalamiento multidimensional. Este análisis permitió, de acuerdo con la posición de los diferentes órdenes, establecer el grado de proximidad respecto a cada mesohábitat y sus características físicas o químicas (Borg et al., 2018). Además, se realizó un análisis discriminante canónico generalizado entre mesohábitats basado en las diferencias de abundancia y riqueza total, en relación con las variables físicas y químicas. Los valores de riqueza, abundancia y del índice QBR en cada mesohábitat fueron sometidos a análisis de componentes principales para establecer el efecto de las coberturas vegetales sobre los patrones de riqueza y abundancia de macroinvertebrados en los diferentes microhábitats. Los anteriores análisis fueron ejecutados en el software RWizard versión 4.1 (Guisande et al., 2014).
Resultados
Se presentaron diferencias significativas para la profundidad (F = 17, g.l. = 2, P = 0.0002) y la velocidad (F = 29.3, g.l. = 2, P = 0.00001). La profundidad presentó valores entre 0.13 y 0.31 m en los rizos, 0.28 y 0.54 m en los rápidos y 0.65 y 0.9 m en las pozas. La velocidad presentó valores entre 0.8 y 1.25 m/s en los rizos, 0.8 y 1.7 m/s en los rápidos y 0.1 y 0.4 m/s en las pozas. Entre las variables químicas del agua, solo el oxígeno disuelto mostró diferencias significativas entre hábitat (F = 27.4, g.l. = 2, P = 0.0004). Esta variable osciló entre 8.17 y 8.71 mg/l en los rizos, 7.91 y 8.56 mg/l en los rápidos y 8.19 y 8.7 mg/l en las pozas.
Se recolectaron 4 484 macroinvertebrados, agrupados en 16 órdenes, 46 familias y 75 géneros, siendo más representativos en abundancia los géneros Leptohyphes, Smicridea, Thraulodes, Simulium, Macrelmis, Anacroneuria, Camelobaetidius y Nectopsyche, respectivamente. En los rizos se encontró una riqueza (q0) promedio de 33 taxones y la mayor abundancia (2 338 individuos) con un valor medio de 390 individuos. Destacándose tricópteros del género Smicridea (15 %, 357 individuos), efemerópteros de los géneros Leptohyphes (10 %, 243 individuos), Thraulodes (8.6 %, 202 individuos) y Camelobaetidius (8.4 %, 197 individuos), coleópteros como Macrelmis (7.6 %, 178 individuos) y plecópteros como Anacroneuria (7.2 %, 169 individuos). Los rizos también presentaron los mayores valores de diversidad q1 (13 especies en promedio) y q2 (8 especies en promedio). Los rápidos presentaron una abundancia y riqueza (q0) promedio de 336 individuos y 27 taxones respectivamente, en este mesohábitat predominaron dípteros del género Simulium (17 %, 291 individuos), seguidos nuevamente por efemerópteros como Leptohyphes (16 %, 272 individuos) y Thraulodes (9.5 %, 160 individuos), tricópteros del género Smicridea (7.7 %, 129 individuos) y coleópteros del género Macrelmis (7.3 %, 122 individuos). Por último, en el mesohábitat poza se encontró una abundancia y riqueza (q0) media de 83 individuos y 17 géneros, los más abundantes fueron hemípteros del género Rhagovelia (17 %, 71 individuos), tricópteros de los géneros Nectopsyche (14.6 %, 61 individuos), Grumichella (5.3 %, 22 individuos) y Helicopsyche (4.5 %, 19 individuos), así como efemerópteros del género Farrodes (8.6 %, 36 individuos).
De acuerdo con el análisis de varianza, se encontraron diferencias significativas en la abundancia entre hábitat (F = 4.257, g.l. = 2, P = 0.037), evidenciando los mayores valores en los rizos, seguido de los rápidos y finalmente las pozas con un promedio de 390, 336 y 83, respectivamente (Tabla 1).
Tabla 1 Valores de las variables físicas, químicas y biológicas de cada mesohábitat. / Table 1. Values of physical, chemical and biological variables in each mesohabitat.
| Variable | Mesohábitat | ||||||
| Rizo | Rápido | Poza | |||||
| Rango | Media | Rango | Media | Rango | Media | ||
| Temperatura (°C) | 18.7-25 | 21.5 | 21.2-22.8 | 21.72 | 18-25.9 | 22 | |
| OD (mg/l) | 8.16-8.19 | 8.4 | 7.9-8.6 | 8.29 | 8.19-8.7 | 8.43 | |
| Saturación de oxígeno (%) | 105.2-112.6 | 107.13 | 103.5-107.7 | 105.78 | 104.1-112 | 106.7 | |
| pH | 6.95-7.43 | 7.39 | 7.1-7.7 | 7.48 | 6.9-7.8 | 7.37 | |
| Conductividad (µS/cm) | 28.6-104 | 50.46 | 29.1-104.4 | 55.68 | 31.3-61.9 | 47.72 | |
| Turbiedad (NTU) | 0.54-2.77 | 1.27 | 0.69-3.31 | 1.8 | 0.59-3.31 | 1.39 | |
| Profundidad (m) | 0.13-0.31 | 0.25 | 0.28-0.54 | 0.35 | 0.65-0.90 | 0.64 | |
| Velocidad (m/s) | 0.80-1.25 | 1 | 0.80-1.70 | 1.32 | 0.10-0.40 | 0.26 | |
| Abundancia | 122-663 | 389.7 | 133-570 | 335.6 | 36-175 | 83.4 | |
| Riqueza (q0) | 17-51 | 33 | 4-26 | 17 | 11-15 | 13 | |
| Equidad (q1) | 7.4-24.8 | 13.2 | 1-18.4 | 12.08 | 4.3-12.5 | 8.87 | |
| Dominancia (q2) | 3.9-17.3 | 8.05 | 1-12.7 | 7.91 | 2.5-9.2 | 6 | |
El análisis discriminante generalizado mostró un 50 % de casos correctamente identificados por validación cruzada. El primer eje explicó el 90.48 % de la varianza y determinó que el rizo es el mesohábitat de mayor preferencia por los macroinvertebrados de los órdenes Coleóptera, Ephemeroptera, Plecoptera, Lepidoptera, Trichoptera, Megaloptera y Odonata, mientras que los órdenes Acari y Díptera están más relacionados con los rápidos. Finalmente, los órdenes Hemiptera, Tricladida y Haplotaxida presentan una tendencia de preferencias hacia las pozas (Fig. 2).
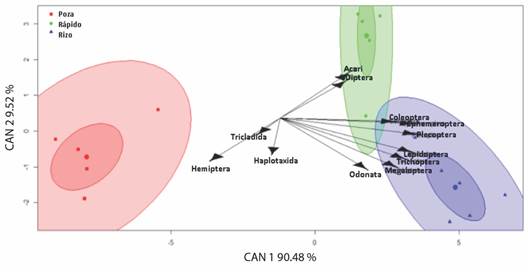
Fig. 2 Biplot del análisis discriminante de los mesohábitat con base en la composición de los órdenes de macroinvertebrados. / Fig. 2. Biplot of the discriminating analysis of the mesohabitat based on the composition of the macroinvertebrate orders.
El mapa de calor identificó 12 familias que presentaron el menor valor de varianza en el análisis PCA y no fueron incluidas en el cluster. Particularmente bajos fueron las familias Ampullaridae, Planorbidae, Thiaridae, Noteridae y Notonectidae que no presentan variaciones reconocibles entre mesohábitat. También presentan baja diferenciación entre mesohábitat las familias Dugesiidae, Calopterygidae, Leptoceridae, Chironomidae, Polycentropodidae y Empididae, que presentan la misma tonalidad de color igual o semejante en todo el gradiente. El grupo conformado por las familias Elmidae, Hydrobiosidae, Ptilodactylidae, Leptohyphidae, Plythoridae, Philopotamidae, Glossosomatidae, Perlidae, Crambidae y Baetidae mostró mayor preferencia por los mesohábitats rizo y rápido, presentándose como un grupo diferenciado en la parte superior del dendrograma, separados de Corydalidae que prefiere los rizos. Asimismo, Acari, Psephenidae y Simuliidae muestran afinidad con el mesohábitat rápido y contribuyen en su diferenciación. Mientras que las familias Euthyplociidae, Veliidae, Calopterygidae, Odontoceridae y Haplotaxidae muestran preferencia por el mesohábitat poza, al igual que la familia Dugesiidae que, aunque tiene amplia distribución, presentó en este hábitat la máxima abundancia. Las familias Naucoridae, Coenagrionidae, Helicopsychidae y Blepharoceridae estuvieron presentes tanto en rizos como en pozas (Fig. 3).
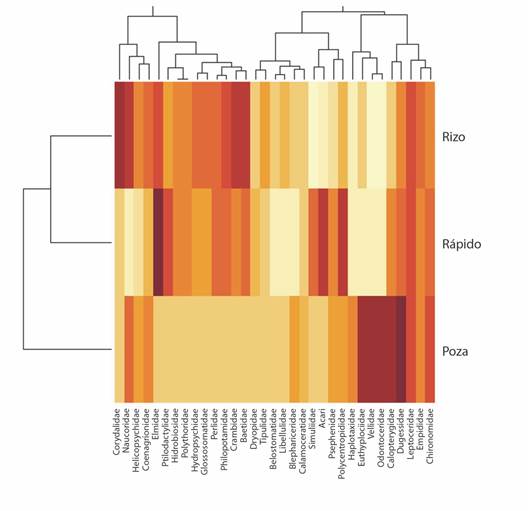
Fig. 3 Mapa de calor de familias de macroinvertebrados a partir de su distribución en diferentes mesohábitats. Las variables que tienen la misma tonalidad son las que menos influyen en la clasificación, las máximas relaciones están representadas por colores más oscuros y la mínima relación por los colores más claros. / Fig. 3. Heat map of macroinvertebrates families from distribution in different mesohabitats. The variables that have the same hue are the ones that least influence the classification, the maximum relationships are represented by the darkest colors and the minimum relationship by the clearest colors.
El análisis de escalamiento multidimensional métrico de la abundancia de distintos órdenes de macroinvertebrados mostró un buen ajuste al modelo (stress 0.027) y evidenció la diferencia de la abundancia de los órdenes Acari, Coleoptera, Plecoptera, Trichoptera, Odonata, Ephemeroptera, Lepidoptera, Diptera y Megaloptera en los rizos y los rápidos, así como la diferencia de la abundancia de los órdenes Tricladida, Hemiptera, Haplotaxida y Neotaenioglosa en las pozas. Asimismo, los rizos y los rápidos se diferenciaron por variables ambientales como la velocidad, la conductividad, la temperatura, el oxígeno disuelto, la turbiedad y el tipo de sustrato, en tanto que las pozas presentan mayor cercanía con la profundidad (Fig. 4).
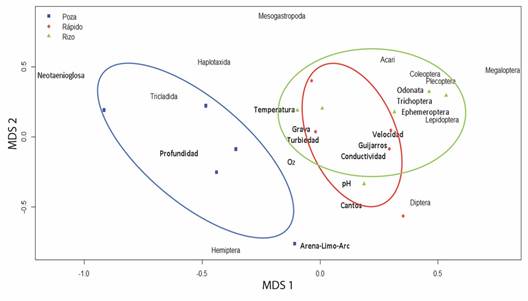
Fig. 4 Análisis de escalamiento multidimensional entre ordenes de macroinvertebrados con variables ambientales en relación con su preferencia entre hábitat. / Fig. 4. Multidimensional scaling analysis between macroinvertebrate orders with environmental variables in relation to its habitat preference.
El análisis discriminante mostró un 72 % de casos correctamente identificados por validación cruzada. El primer eje explicó un 96.3 % de la varianza y determinó que los mesohábitat rizo y rápido, que presentan mayor velocidad, fueron discriminados por mayores valores de abundancia y riqueza de macroinvertebrados (promedio = 365 individuos y 30 taxones). El mesohábitat poza, fue discriminado por una mayor profundidad y menores valores de abundancia y riqueza de macroinvertebrados, con un promedio 83 individuos y 17 taxones (Fig. 5).
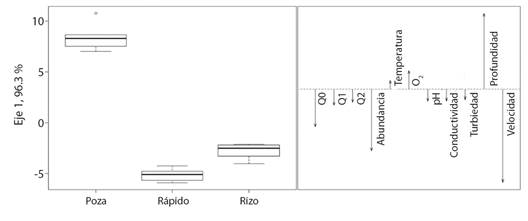
Fig. 5 Análisis discriminante canónico para la abundancia de macroinvertebrados y variables ambientales. O2-Oxígeno disuelto (mg/l), temperatura (°C), pH, conductividad (µS/cm), Turbiedad (NTU), profundidad (m), velocidad (m/s), riqueza (q0), equidad (q1), especies dominantes (q2). / Fig. 5. Canonical discriminating analysis for the abundance of macroinvertebrates and environmental variables. O2-Dissolved oxygen (mg/l), temperature (°C), pH, conductivity (µs/cm), turbidity (NTU), depth (m), speed (m/s), wealth (q0), equitativity (q1), dominant species (q2).
De acuerdo con los resultados del índice de la calidad de la vegetación de ribera Andina (QBR-And), las quebradas La Carbonera (S1), La Isla (S5 y S6) y el Río Melcocho (La Isla, S7), presentan una vegetación de ribera sin alteraciones y se encuentra en estado natural (QBR-And > 95); el río Melcocho (Puente Amarillo (PA) S4) presenta una vegetación ligeramente perturbada (QBR- And entre 75 y 95), no obstante, de buena calidad, caracterizado por un bosque de ribera con buena estructura, poco intervenido, árboles de buen porte (Montoya-Cardona, 2022). La Calera (S3) presentó inicio de alteración importante, con calidad intermedia (QBR-And entre 51 y 75) y las quebradas San Antonio (S2), La Trinidad (S8) y Río Tafetanes (S9 y S10) degradación extrema, calidad pésima en su vegetación de ribera (QBR-And < 25) (Fig. 6).
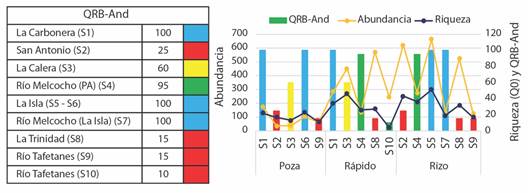
Fig. 6 Índice QBR-And comparando la riqueza y abundancia de macroinvertebrados en los diferentes sitios de muestreo. Azul (> 96): Vegetación de ribera sin alteraciones, calidad muy buena, estado natural. Verde (75-95): Vegetación ligeramente perturbada, calidad buena. Amarillo (51-75): Inicio de alteración importante, calidad intermedia. Naranja (26-50): Alteración fuerte, mala calidad. Rojo (< 25): Degradación extrema, calidad pésima. / Fig. 6. QBR-And index comparing the richness and abundance of macroinvertebrates in the different sampling sites. Blue (> 96): Ribera vegetation without alterations, very good quality, natural state. Green (75-95): Slightly disturbed vegetation, good quality. Yellow (51-75): Start of important alteration, intermediate quality. Orange (26-50): Strong alteration, poor quality. Red (< 25): Extreme degradation, lousy quality.
El análisis de componentes principales realizado para establecer la varianza de la diversidad y la abundancia entre muestras, obtuvo una varianza acumulada de 99.92 % en los dos primeros ejes y mostró la separación se sitios con valores de riqueza contrastantes en función del índice QBR. El primer eje diferenció las muestras con mayores valores de QBR-And en los que la riqueza fue mayor, asimismo (en su eje negativo) los mesohábitats con una alteración fuerte y mala calidad del bosque de ribera, en los que la riqueza de macroinvertebrados presentó los menores valores. La abundancia no mostró relación con el índice QBR-And.
Discusión
Los resultados muestran que no hay diferencias significativas en las variables químicas entre los mesohábitats excepto para la concentración de oxígeno disuelto. De acuerdo con Roldán-Pérez y Ramírez-Restrepo (2022), el oxígeno es fundamental para varios procesos vitales como la respiración, oxidación de la materia orgánica y nitrificación. Tal como lo describen Dias da Silva et al. (2015), su concentración en los mesohábitats ocurre en función de las condiciones hidráulicas creadas por la combinación de la velocidad, características de los sustratos y la profundidad. Es así como el ingreso de oxígeno por subsidio atmosférico, que sería mayor en un mesohábitat como los rápidos, afectaría procesos funcionales clave en este tipo de ecosistemas. Las diferencias en las variables físicas como la profundidad y la velocidad son notables entre mesohábitat, evidenciando características propias de cada uno de ellos. Los rizos y rápidos se caracterizaron por valores mayores de velocidad y menor profundidad propiciando una adecuada oxigenación, mientras que las pozas se distinguieron por ser más profundas y presentan un flujo más lento similar a lo reportado por Parasiewicz et al. (2009).
Este estudio evidencia una alta diversidad de macroinvertebrados que se encuentran presentes en los diferentes mesohábitats de las corrientes de montaña del Oriente antioqueño, tanto en términos de riqueza total (q0), como de especies comunes (q1) y dominantes (q2). Además, los valores de riqueza se consideran representativos de la región en relación con los reportados por Barros-Núñez (2020). Congruente con otros estudios (Burgazzi et al., 2021; Principe et al., 2007; Vimos-Lojano, 2017), los mesohábitat con mayor diversidad y abundancia fueron los rizos y rápidos debido a la influencia en la disponibilidad de recursos alimenticios, tipo de sustrato y perturbaciones físicas que permiten explicar una mayor abundancia de organismos ETP en este tipo de mesohábitat. En especial, porque muchos de ellos cuentan con diversas adaptaciones morfológicas que les permiten sobrevivir a condiciones de mayor velocidad, tales como ganchos, ventosas, cuerpos delgados y aplanados (Elosegi & Sabater, 2009; Molina et al., 2008; Roldán-Pérez & Ramírez-Restrepo, 2022). En las pozas la entrada continua y la acumulación de materia orgánica bentónica gruesa y fina permite explicar las mayores abundancias de invertebrados trituradores como Nectopsyche reconocido por su preferencia por hábitats con bajas velocidades donde se acumula material de piedras y hojas que es utilizado para la construcción de casas o como recurso alimenticio (Holzenthal & Ríos-Touma, 2018). Nuestros resultados destacan una comunidad de macroinvertebrados espacialmente estructurada, siendo la composición y la diversidad variables con mayor dependencia del tipo de mesohábitat, tal como fue registrado por Burgazzi et al. (2021). Mientras que la abundancia solo presenta disminución en las pozas, pero no diferencia rápidos y rizos.
Buenas condiciones físicas y químicas como variedad de sustratos, disponibilidad de recursos, aguas con buena oxigenación y poco profundas son factores comunes en corrientes de montaña y preferidas por la mayoría de los macroinvertebrados (Barros-Núñez, 2020). Tal es el caso de los géneros Leptohyphes, Smicridea, Thraulodes, Simulium, Macrelmis, Anacroneuria, Camelobaetidius y Nectopsyche, que fueron predominantes en los mesohábitats rizo y rápido. Mientras que, en las pozas las condiciones de baja velocidad, la acumulación de materiales como hojas y piedras es aprovechado por algunos macroinvertebrados como Grumichella, Helicopsyche y nuevamente Nectopsyche, para la elaboración de casas a partir de restos vegetales, piedras pequeñas y granos de arena. Este resultado concuerda con lo reportado por Díaz-Rojas et al. (2020) y Motta-Díaz et al. (2016) en donde se muestra una preferencia notable de determinados organismos como Simulium y Nectopsyche por hábitats con características contrastantes en respuesta a sus rasgos morfológicos y funcionales (Domínguez & Fernández, 2009; Rolls et al., 2012). En las pozas predominaron también Rhagovelia y Farrodes, que al igual Grumichella, Helicopsyche, poseen adaptaciones como tubos o sifones respiratorios, escamas o pelos microscópicos, branquias más grandes y cuerpos pequeños para vivir en este mesohábitat (Merritt et al., 2017). De acuerdo con Rolls et al. (2012), el caudal regula la variedad, la disponibilidad y el área de los mesohábitat. Su efecto depende, en gran medida, de la interacción a escala local entre la hidrología y la geomorfología, regulada a su vez por cambios en las condiciones hidráulicas. De manera que, los caudales bajos disminuyen el volumen, el área y la profundidad y cambian la velocidad de las corrientes, provocando aumento temporal en la abundancia y riqueza de la biota, especialmente los invertebrados (Rolls et al., 2012). En este sentido, tanto Blanckaert et al. (2013) como Motta-Díaz y Vimos-Lojano (2020) destacan la influencia de los parámetros hidráulicos como velocidad, profundidad y rugosidad del lecho en la estructura de la comunidad de macroinvertebrados.
En concordancia con los resultados de este estudio, Galeano-Rendón y Mancera-Rodríguez (2018) encontraron que durante el período de transición de seco-lluvia destacan órdenes como Trichoptera, Ephemeroptera y Diptera con mayores valores de abundancia. Vázquez et al. (2020) en su estudio sobre idoneidad de hábitat para macroinvertebrados también encontraron que en ríos andinos tropicales algunos géneros de estos órdenes se asocian a flujos más rápidos y turbulentos, como resultado de sus adaptaciones específicas a este entorno. Asimismo, González-Tuta y Gil-Padilla (2020) y Principe et al. (2007) encontraron que familias como Coenagrionidae, Hydrobiosidae, Leptohyphidae, Glossosomatidae, Perlidae, Baetidae, Elmidae, Psephenidae y Simuliidae tienen mayor preferencia por mesohábitats con sustratos conformados por cantos rodados y en general, superficies rocosas donde la velocidad es mayor. Quesada-Alvarado et al. (2021) ratifican la preferencia de estas familias por velocidades de agua moderadas a rápidas (0.2-1 m/s), como se encontró en este estudio. La presencia de diversos sustratos como grava, guijarros, rocas grandes y cantos combinados con elementos como la hojarasca y restos de madera propician las condiciones necesarias para el establecimiento de dichas comunidades, favoreciendo así una mayor diversidad (Duarte-Ramos & Reinoso-Flórez, 2020; Váquiro-Capera et al., 2020). Además, mientras los rizos están formados por cantos rodados, los rápidos están formados de rocas grandes ancladas en la tierra, las cuales, como consecuencia de las variaciones en la velocidad del agua, son influenciadas continuamente por procesos de deriva favoreciendo la colonización debido a la retención de vegetación y de macroinvertebrados. De acuerdo con Ríos-Touma et al. (2011) su persistencia depende de la disponibilidad de áreas que sean menos susceptibles a los impactos de la perturbación como refugios, cuyas características físicas aumentan la probabilidad de subsistir a condiciones severas y, por lo tanto, permiten explicar la presencia de comunidades de macroinvertebrados con mayor riqueza y diversidad en los mesohábitat de rizos y rápidos. Estos resultados resaltan tanto la importancia de adaptaciones específicas de los organismos para ocupar un mesohábitat determinado, así como la dependencia de los atributos de la comunidad de la estructura física del sistema (Burgazzi et al., 2021).
De acuerdo con Roldán-Pérez y Ramírez-Restrepo (2022) y Quesada-Alvarado et al. (2021), las familias Veliidae, Calopterygidae y Dugesiidae, que mostraron afinidad con el mesohábitat poza, se caracterizan por vivir en aguas con poca corriente o baja velocidad, con sedimentos ricos en detritos orgánicos y hojarasca y baja concentración de oxígeno. Euthyplociidae, por su parte ha sido reportada para aguas rápidas con sustratos arenosos, lo que no corresponde con lo encontrado en este estudio. No obstante, el bajo número de individuos recolectados no permite un resultado concluyente sobre sus preferencias de hábitat. Debido a sus hábitos minadores que les permite cavar túneles en el sustrato del cuerpo de agua y dificultan su captura (Gutiérrez & Reinoso-Flórez, 2010). Familias como Naucoridae, Helicopsychidae, Coenagrionidae y Polycentropodidae aprovecharon tanto pozas como rizos, pero fueron restringidas por velocidades muy altas en los rápidos. Polycentropodidae, por ejemplo, se caracteriza por construir refugios fijos y redes amplias de hilo en aguas poco profundas (para depredación y recolección), en rangos de velocidad de lentos a moderados (Van den Brink et al., 2013), de ahí que se encuentre tanto en pozas como en rizos. Naucoridae presenta géneros adaptados a velocidades moderadas a rápidas como Cryphocricos y lentas como Limnocoris y Coenagrionidae estuvo representado por el Argia, adaptado a velocidades moderadas, pero sobre gran variedad de sustratos como rocas, hojarasca, troncos o macrófitas (Mosquera-Murillo & Mosquera-Mosquera, 2021; Quesada-Alvarado et al., 2021). Esta variabilidad en las preferencias genéricas por el hábitat, también se observa en otros grupos. De acuerdo con Oliveros-Villanueva et al. (2020), los Tricópteros son organismos adaptados para sobrevivir en corrientes de agua fuertes, que contribuyen a la retención de oxígeno y acumulación del material orgánico a lo largo del cauce. Este patrón se cumple para la mayoría de los géneros recolectados en este estudio. No obstante, también se ratifica la variabilidad en las preferencias genéricas por el hábitat, en el caso particular de Nectopsyche, que fue encontrado tanto en pozas como en rizos y rápidos, lo cual se relaciona con el amplio rango de velocidades en que puede establecerse (Quesada-Alvarado et al., 2021). En cuanto a los coleópteros, González-Córdoba et al. (2020) describen igualmente un amplio rango de preferencia por el tipo de sustrato y velocidad. Pese a la versatilidad de muchos grupos, debido a que en las pozas son más susceptibles a la depredación y no disponen de sustratos heterogéneos para su establecimiento (Brand et al., 2012), este mesohábitat presentó menor abundancia y diversidad de macroinvertebrados que los rápidos y rizos.
Acompañando las variables hidráulicas, la cobertura vegetal aparece como un elemento importante en el establecimiento de los macroinvertebrados en las corrientes estudiadas. La diversidad y abundancia de macroinvertebrados es mayor en los sitios que presentan más variabilidad en el sustrato y mejores condiciones de cobertura vegetal, afectando especialmente su composición funcional, tal como se ha documentado en Colombia (Barros-Núñez, 2020; Galeano-Rendón & Mancera-Rodríguez, 2018; Meza-Salazar et al., 2012; Vera-Sánchez & Pinilla-Agudelo, 2020; Walteros-Rodríguez & Castaño-Rojas, 2020) y en otros países (Fierro et al., 2017; Oester et al., 2022; Ono et al., 2020). Esta dependencia entre las características de la ribera y la estructura de la comunidad de macroinvertebrados obedece a que la entrada de material alóctono como hojarasca, semillas, ramas y troncos representa una fuente fundamental de energía y nutrientes para los macroinvertebrados, especialmente los trituradores (Vimos-Lojano, 2017). Esto se refleja en la abundancia de géneros ETP con este hábito en los ríos que presentaron mayores coberturas de vegetación. Además, la hojarasca y restos vegetales que son aportados desde la ribera también se convierten en sustrato para la colonización del perifiton, un biofilm que proporcionan alimento para invertebrados raspadores, incluyendo a muchos tricópteros y efemerópteros (Flowers & De la Rosa, 2010; Wiggins, 2004). Asimismo, provee refugio frente a las variaciones del caudal y la presencia de depredadores (Ríos-Touma et al., 2011).
Por último, si bien los macroinvertebrados continúan siendo organismos fundamentales para el establecimiento de las condiciones de calidad de agua en una corriente, es recomendable ser cauteloso con la manera en que son usados en la bioindicación, ya que, como se observa en este estudio, los patrones de composición y abundancia de macroinvertebrados pueden estar reflejando también, en gran medida, las condiciones físicas del hábitat. Es por ello, que cuando se evalúan las condiciones de calidad del agua, simultáneamente se debe caracterizar el hábitat físico y considerar sus efectos sobre la dinámica de esta comunidad. A fin de garantizar que no se subestime la riqueza ni se omita información determinante para el establecimiento o no de las especies. Los resultados de este estudio evidencian que las variables del hábitat como la velocidad, la profundidad y el sustrato, tienen mayor importancia que las variables químicas del agua en la distribución de macroinvertebrados en las corrientes del Oriente antioqueño y contribuyen a diferenciar la composición de los mesohábitats rizo, poza y rápido. Siendo la concentración de oxígeno, la única variable química relevante en la distribución de estos organismos, confirmando así la hipótesis planteada. De igual forma se encontró que las corrientes que no presentan alteración en su cobertura vegetal ofrecen mejores condiciones para el establecimiento de los macroinvertebrados, independientemente del mesohábitat, lo cual se refleja en mayores valores de riqueza y diversidad, mientras que en corrientes deforestadas se observaron abundancias relativamente altas pero muy baja diversidad, lo que significa que dichas afectaciones repercuten en la composición de la comunidad de macroinvertebrados. En este contexto, garantizar tanto la cobertura vegetal como las condiciones hidráulicas de las corrientes es fundamental para conservar la diversidad de los macroinvertebrados.
Declaración de ética: Los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría, que no hay conflicto de interés de ningún tipo y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

