










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.63 n.2 San José Apr./Jun. 2015
Longevity and germination of Syagrus romanzoffiana (Arecaceae) seeds and its ecological implications
Longevidad y germinación de semillas de Syagrus romanzoffiana (Arecaceae) y sus implicaciones ecológicas
Longevidad y germinación de semillas de Syagrus romanzoffiana (Arecaceae) y sus implicaciones ecológicas
Túlio Gabriel Soares Oliveira1*, Anderson Cleiton José2*, Leonardo Monteiro Ribeiro3* & José Marcio Rocha Faria2
Abstract
Syagrus romanzoffiana is a palm tree native and widely distributed of South America. The present study investigated the longevity and germination of the buried seeds of this species in an experimental seed bank. Laboratory germination and viability tests were performed for comparison with field results. Pyrenes (seeds enclosed by the endocarp) were buried in a forest fragment edge in July 2012 (dry season) and exhumed monthly during one year, for the assessment of water content and percentage of germinated and viable seeds. Germination tests were conducted in a Mangelsdorf-type germinator at 30°C under constant light and the viability was assessed by the tetrazolium test. An additional sample of pyrenes was buried to evaluate the percentage of seedling emergence and survival. Climatic and soil moisture data were recorded. In the laboratory, the pyrenes were stored for one year in a temperature-controlled room at 20ºC and 75% (±10%) relative air humidity to assess changes in the percentage of germination and viability over time. In the field, a reduction in seed viability was observed over the study period, with a total loss of viability of non-germinated seeds at seven months after burial. The maximum germination (close to 26%) was observed in the samples that were exhumed between five and seven months after burial. In the field, seedling emergence did not exceed 10% and seedling mortality was not observed. The percentages of germination and of viable seeds decreased both in burial and stored seeds. The stored seeds maintained viability at up to six months, with marked reduction thereafter. After the germination tests (four months) in the laboratory, all of the remaining seeds were nonviable. The ecological, physiological and reproductive characteristics of the species are discussed, and we concluded that S. romanzoffiana seeds have short longevity after imbibition, and low potential for soil seed bank formation. Rev. Biol. Trop. 63 (2): 333-340. Epub 2015 June 01.
Key words: palm tree, queen palm, viability, seed bank, seed storage.
Resumen
Syagrus romanzoffiana (Arecaceae) es una palma nativa y ampliamente distribuida de América del Sur. El presente estudio investigó la longevidad y la germinación de las semillas enterradas de esta especie en un banco experimental. Se realizaron pruebas de germinación y viabilidad de laboratorio para hacer una comparación con los resultados de campo. Pirenos (semillas encerradas por el endocarpio) fueron enterrados en el borde de un fragmento de bosque en julio (época seca) y exhumados mensualmente por un año, para la evaluación del contenido de agua y del porcentaje de semillas germinadas y viables. Las pruebas de germinación se realizaron en un germinador tipo Mangelsdorf a 30°C bajo luz constante y la evaluación de la viabilidad se realizó con la prueba de tetrazólio. Una muestra adicional de pirenos fue enterrada para evaluar el porcentaje de emergencia de las plántulas. Se tomaron los datos climáticos y se determinó la humedad del suelo. En el laboratorio, los pirenos se almacenaron durante un año en una sala de temperatura controlada a 20°C y 75% (±10%) de humedad relativa del aire, para evaluar los cambios en el porcentaje de germinación y la viabilidad en el tiempo. En el campo, se observó una reducción de la viabilidad de las semillas durante el período de estudio, con una pérdida total de la viabilidad de las semillas no germinadas a los siete meses después del entierro. La germinación máxima (cerca de 26%) se observó en las muestras que fueron exhumadas entre cinco y siete meses después del entierro. En el campo, la emergencia de las plántulas no superó el 10% y no se observó mortalidad de plántulas. Los porcentajes de germinación y de semillas viables disminuyeron tanto en el suelo como en el laboratorio. Las semillas almacenadas mantuvieron la viabilidad hasta por seis meses de almacenamiento, con una reducción marcada a partir de entonces. Después de las pruebas de germinación (cuatro meses) en el laboratorio, todas las semillas restantes eran inviables. Discutimos las características ecológicas, fisiológicas y reproductivas de la especie, y llegamos a la conclusión de que las semillas de S. romanzoffiana tienen corta duración después de la imbibición y bajo potencial de formación del banco de semillas en el suelo.
Palabras clave: palma, palma de la reina, viabilidad, banco de semillas, almacenamiento.
The longevity or ability to preserve seed viability is essential for the formation of a persistent soil seed bank (Thompson, 1997), which plays a decisive role in the adaptation of species to different environments (Baskin, & Baskin, 2014; Long et al., 2015). Because of the peculiarities in different habitats, the most accurate method of assessing seed longevity is to bury them in the ground and remove samples at certain time intervals to determine their viability (Baskin, & Baskin, 2006; 2014).
Syagrus romanzoffiana (queen palm) is a native palm tree that is widely distributed in South America (Lorenzi, Souza, Medeiros-Costa, Cerqueira, & Behr, 1996). In Brazil, this species is found in several phytophysiognomies that have significant differences related to climate and vegetation structure and include subtropical forests, dry forests, Brazilian savanna (Cerrado), Atlantic Forest and Restinga (Guix, & Ruiz, 2000; Brancalion, Novembre, & Rodrigues, 2011). The queen palm produces fleshy fruits and is an ecologically important species because of its interaction with frugivorous birds and mammals that also act as seed dispersers (Giombini, Bravo, & Martínez, 2009).
Studies have reported low germination percentages for S. romanzoffiana seeds in the laboratory (Broschat, & Donselman, 1987; Brancalion et al., 2011; Goudel, Shibata, Coelho, & Miller, 2013), suggesting that the seeds have a certain degree of dormancy. The presence of dormancy and the ability to form persistent seed banks are adaptive strategies of plant species to overcome unfavorable periods for germination and/or seedling establishment (Moles, Warton, & Westoby, 2003; Long et al., 2015). Although the presence of dormancy may contribute to the formation of a persistent seed bank as a result of maintenance of viability (Jensen, 2004), certain studies have shown that there is no a close relationship between dormancy and persistence in the seed bank (Thompson, Ceriani, Bakker, & Bekker, 2003; Honda, 2008).
Under natural conditions, S. romanzoffiana seeds are dispersed within the fruit structures. After the pulp (mesocarp) is consumed, a woody endocarp remains enclosing the seed that forms a germination unit called pyrene. The presence of fruit structures (such as the endocarp) restricts germination in certain palm tree seeds by imposing a mechanical barrier for germination (Broschat, 1998; Ribeiro, Souza, Rodrigues-Jr, Oliveira, & Garcia, 2011; Neves et al., 2013). Moreover, these structures may play an important ecological role, especially in the prevention of water loss and protection against predation, thus contributing to an increase in seed longevity (Orozco-Segovia, Batis, Roja-Aréchiga, & Mendoza, 2003; Hu, Wang, & Wu, 2009).
The longevity of seeds, their mechanisms of dormancy, germination in the field and the effects of environmental conditions on these processes must be determined to understand the dynamics of plant populations, however, there is little information in the literature on these approaches (Vázquez-Yanes & Orozco-Segovia, 1993). In the present study, we sought to assess changes in the germination, viability and water content of S. romanzoffiana seeds in natural and laboratory conditions to determine the following: i) are there differences in the longevity of laboratory stored and buried seeds? ii) does the species form a persistent seed bank? and iii) does the behavior of seeds in the bank explain the reproductive success of the species?
Materials and methods
Collection and processing of seeds: Ripe fruits, which are characterized by an orange color, were collected in July 2012 immediately after natural abscission from eighteen individuals of S. romanzoffiana present at the campus of the Federal University of Lavras (Universidade Federal de Lavras-UFLA), Minas Gerais State, Brazil. The fruits were immediately placed in sealed plastic bags and left under shade for five days to soften the pulp. The fruits were then depulped using a vertical electric depulper device to obtain the pyrenes (endocarp+seed), which were then transported to the Laboratory of Forest Seeds/UFLA to carry out the experiments.
Initial characterization of the seeds: To assess the germination, the pyrenes (four replicates of 25) were surface sterilized in a 3% sodium hypochlorite solution for ten minutes, rinsed in tap water and sown in polyethylene trays containing autoclaved sand in a Mangelsdorf-type germinator at 30°C under constant light. Seedling emergence was used as the germination criterion, and it was evaluated weekly for four months to calculate the final germination percentage (%G). The water content of the seeds after removing the endocarp was determined by the oven method at 105ºC for 24h (Ministério da Agricultura, Pecuária e Abastecimento, Brasil, 2009) using four replicates of five seeds. The isolated seeds were obtained after breaking the pyrenes in a manual vise. The initial viability was determined by the 0.5% tetrazolium test according to the method for the macaw palm (Acrocomia aculeata) described by Ribeiro, Garcia, Oliveira & Neves (2010) using four replicates of ten embryos.
Seed behavior in an experimental bank: Pyrenes whose seeds had 18% of water content were buried in at the border of a forest fragment at UFLA to simulate the natural dispersion and formation of seed banks of this species. The pyrenes were placed in polyethylene nets (one for each sample point) with 1cm mesh and covered by 3cm of soil from the study area. The experiment consisted of 12 samples of 260 pyrenes, which were sampled monthly during one year (between July 2012 and July 2013) to assess germination percentage (four replicates of 40 pyrenes), viability (four replicates of 15 pyrenes) and seed water content (four replicates of 10 pyrenes). The emission of cotyledon petiole was considered as an indicator of germination (Ribeiro et al., 2011). The soil moisture of the study site was assessed concurrently with the exhumation of the samples by the gravimetric method after drying in an oven at 105ºC for 24 hours. Viability was determined only in the seeds that did not germinate. For this, the pyrenes were broken and the seeds were initially characterized as apparently viable (whole seed and embryo) or nonviable (deteriorated, oxidized or preyed upon seed or embryo). The embryos (when present) were then excised from the seeds and subjected to the tetrazolium test (Ribeiro et al., 2010). To calculate the viability, the percentage of non-viable seeds in the preliminary assessment was considered.
Additionally, eight replicates of 50 pyrenes were buried for the monthly evaluation of the emergence and survival of the seedlings. For this purpose, the samples were kept in the seed bank during the entire sampling period. Local climatological data were obtained from the UFLA Meteorological Station, which is located approximately 1km from the study site.
Storage of pyrenes: Pyrenes containing seeds with a water content of 18% were stored in polyethylene bags (semipermeable) in a temperature-controlled room at 20ºC and 75% (±10%) relative air humidity. The %G, viability by the tetrazolium test, and seed water content were determined after 3, 6 and 12 months of storage. Germination was assessed as previously described.
Data analysis: The percentage data were transformed to arcsine and subjected to the Shapiro-Wilk normality test. An analysis of variance was performed, and the Scott-Knott test was used at 5% probability.
Results
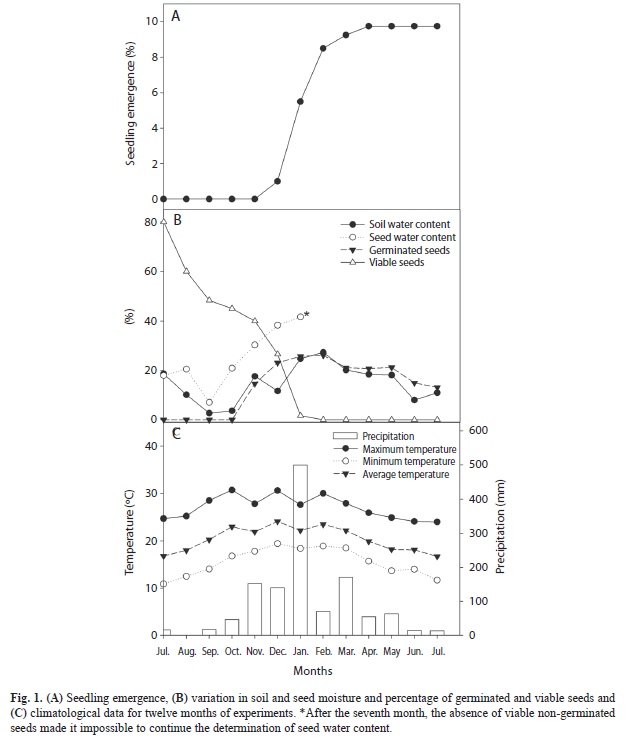
Seed behavior in an experimental bank: The seeds presented an initial viability of 80% by the tetrazolium test. After six months, there was a gradual reduction in viability until it almost reached zero. The seed water content declined after two months and was followed by a gradual increase until the sixth month, when 82% of the seeds were deteriorated. The increase in water content of seeds was related to increasing precipitation (Fig. 1C). In the seventh month, all of the seeds that did not germinate were deteriorated (Fig. 1B) and thus their water content was no longer evaluated. Germination began in the fourth month after burial and reached the maximum percentage in the seventh month (26%). Seeds that germinated (cotyledon petiole elongated) and subsequently died, were observed and counted as germinated. The soil moisture showed large variations throughout the experiment and presented a value below 3% (wet basis) in the second month. Seedling emergence was observed only after five months and stabilized at nine months when it reached its maximum percentage (10%) (Fig. 1A). No seedling mortality was observed during the study period.
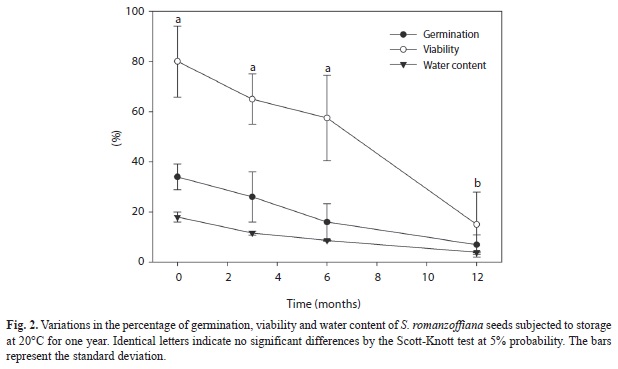
Seed storage: Freshly collected seeds, with 18% water content showed 34% germination and 80% viability. Seed viability was maintained for up to six months of storage (Fig. 2). After this period, there was a marked reduction down to 15% at the end of the storage. The germination percentage was maintained for up to three months of storage and declined thereafter. The seeds water content was reduced during the storage.
Discussion
S. romanzoffiana seeds buried in the experimental seed bank showed short longevity, with total seed mortality after seven months of burial, characteristic of a transient seed bank. The increased precipitation and consequential increase in seed water content were most likely related to increased deterioration. The seeds stored in dry conditions maintained 15% viability after one year, whereas in the field, the seeds imbibed after the first rain and those that did not germinate, lost their viability. The low longevity of seeds after imbibition was also observed in the laboratory. At the end of the germination tests (four months), all of the remaining seeds (non-germinated) were deteriorated or nonviable. Thus, the behavior of seeds in the field and laboratory is similar with respect to loss viability. The same behavior was also observed in stored and buried seeds of palm Thrinax radiata (Pérez, González, & Irabién, 2005). Such behavior in which the seeds that do not germinate eventually die by deterioration may be an indication that dormancy does not play an important role in the reproductive strategies of S. romanzoffiana (Brancalion et al., 2011) because the ecological advantage of dormancy cannot be exploited in seeds that have short longevity after imbibition. The observed high percentage of deterioration in S. romanzoffiana is most likely related to the high lipid content of the seeds (Coimbra, & Jorge, 2011) since they are tolerant to desiccation (unpublished data).
S. romanzoffiana seeds presented a low potential to form seed banks and are unable to form a persistent seed bank. Few studies have evaluated the palm trees capacity to form seed banks (Dickie, Balick, & Linington, 1992; Ribeiro et al., 2012), which is of great importance for the regeneration of plant communities (Fenner, 1992). S. romanzoffiana shows abundant fruiting throughout the year (Genini, Galetti, & Morellato, 2009; Freire, Closel, Hasui, & Ramos, 2013), therefore, the continuous replenishment of viable seeds in the soil appears to be crucial for the regeneration of the species, apparently compensating for the low seed longevity and inability to form a persistent seed bank.
The distribution of fruiting throughout the year implies that S. romanzoffiana seeds may be subject to different environmental conditions after dispersion because the species grows in seasonal environments. The peak fruiting is variable but usually concentrated between March and May (Genini et al., 2009; Freire et al., 2013). Considering the time required to reach full maturation and dispersal of the fruits, the seed bank installed in July managed to reflect the common natural behavior for the species. The seeds dispersed in July (dry season), however, may present different behaviors in relation to germination and maintenance of viability when compared with the seeds that were dispersed and incorporated in the bank in the rainy season.
S. romanzoffiana grows in contrasting environments (Guix, & Ruiz, 2000; Brancalion et al., 2011), and the characteristics of the place where the seed bank was installed in the present study reflect seasonal environmental conditions with lower vegetation cover, such as those in the Cerrado and dry forest. Schlindwein (2012) observed that seeds of the palm tree Butia odorata maintain higher percentages of viability when buried in forest fragments compared with seeds buried at the edge. Thus, we believe that the behavior of the seeds in the bank is dependent on the habitat and time of dispersal. S. romanzoffiana seeds are dispersed within fruits that contain a fleshy mesocarp that is primarily consumed by birds and mammals before and after dispersion (Giombini et al., 2009). Some of these dispersers may eventually bury the fruits, which favors conservation and allows seed germination when placed in favorable conditions (Orozco-Segovia et al., 2003).
The differences between the percentages of seed germination (26%) and seedling emergence (10%) can be explained by the type of germination found in S. romanzoffiana seeds, which present remote germination in which the cotyledon extends and grows at some distance from the seed (Bernacci, Martins, & Santos, 2008). Thus, many seedlings do not emerge immediately after germination, and the seeds can germinate and die before seedling emergence. This behavior explains the higher percentage of germinated seeds (evaluated after seed exhumation) compared to emergence. The reduction in the percentage of germination after seven months was most likely related to a concentrated germination over a short period of time after the onset of the rainy season. In the months close to the onset of germination, alive germinated seeds were still observed, however, in subsequent months, only dead germinated seeds were observed, which were considered to be germinated. In this study, the reduction in the percentage of germination over time was associated with the death (by predation or deterioration) of the cotyledon petiole of the germinated seeds that remained in the bank for longer periods, thus preventing some of them from being recorded as germinated.
The seeds used in the experiments (field and laboratory) showed 18% water content (after processing). Seed germination in the laboratory and germination in the field reached 34 and 26%, respectively. However, in laboratory conditions, the seedlings were able to emerge and continue their development, whereas in the field, most of the seeds died after germination and consequently did not complete their development. Despite the low percentage of seedling emergence in the field (10%), the absence of seedling mortality suggests that at least some of the seedlings can reach the reproductive age.
We concluded that S. romanzoffiana seeds behave in the same way when stored in the laboratory or buried in the field. The seeds present short longevity after imbibition and, therefore, a low potential for formation of seed banks. The interaction with seed dispersers, abundant fruiting, extended reproductive phenology and hardiness of the seedlings are factors that allow the tree to adapt to contrasting environments (Falasca, Del Fresno, & Ulberich, 2012) and may explain the reproductive success and wide distribution of the species.
Acknowledgments
The authors would like to thank J. C. Martins, for help in collecting the seeds. T. G. S. Oliveira would like to thank the National Council for Scientific and Technological Development (Conselho Nacional de Desenvolvimento Científico e Tecnológico-CNPq) for the concession of a master’s grant.
References
Baskin, C. C., & Baskin, J. M. (2006). The natural history of soil seed banks of arable land. Weed Science, 54, 549-557. [ Links ]
Baskin, C. C., & Baskin, J. M. (2014). Seeds: Ecology, biogeography, and evolution of Dormancy and germination (second edition). San Diego, CA, USA: Academic Press. [ Links ]
Bernacci, L. C., Martins, F. R., & Santos, F. A. M. (2008). Estrutura de estádios ontogenéticos em população nativa da palmeira Syagrus romanzoffiana (Cham.) Glassman (Arecaceae) [Structure of ontogenetic stages in the native population of the palm tree Syagrus romanzoffiana (Cham.) Glassman (Arecaceae)]. Acta Botanica Brasilica, 22(1), 119-130. [ Links ]
Brancalion, P. H. S. A., Novembre, A. D. L. C., & Rodrigues, R. R. (2011). Seed development, yield and quality of two palm species growing in different tropical forest types in SE Brazil: implications for ecological restoration. Seed Science and Technology, 39(2), 412-424. [ Links ]
Broschat, T. K., & Donselman, H. (1987). Effects of fruit maturity, storage, presoaking, and seed cleaning on germination in three species of palms. Journal of Environmental Horticulture, 5(1), 6-9. [ Links ]
Broschat, T. K. (1998). Endocarp removal enhances Butia capitata (Mart.) Becc. (pindo palm) seed germination. Horttechnology, 8(4), 586-587. [ Links ]
Coimbra, M. C., & Jorge, N. (2011). Proximate composition of guariroba (Syagrus oleracea), jerivá (Syagrus romanzof?ana) and macaúba (Acrocomia aculeata) palm fruits. Food Research International, 44(7), 2139-2142. [ Links ]
Dickie, J. B., Balick, M. J., & Linington, I. M. (1992). Experimental investigations into the feasibility of ex situ preservation of palm seeds; an alternative strategy for biological conservation of this economically important plant family. Biodiversity & Conservation, 1(2), 112-119. [ Links ]
Falasca, S. L., Del Fresno, C. M., & Ulberich, C. (2012). Possibilities for growing queen palm (Syagrus romanzofianna) in Argentina as a biodiesel producer under semi-arid climate conditions. International Journal of Hydrogen Energy, 37(19), 14843-14848. [ Links ]
Fenner, M. (1992). Seeds: The ecology of regeneration in plant communities. Wallingford, UK: CABI Publishing. [ Links ]
Freire, C. C., Closel, M. B., Hasui, E., & Ramos, F. N. (2013). Reproductive phenology, seed dispersal and seed predation in Syagrus romanzoffiana in a highly fragmented landscape. Annales Botanici Fennici, 50(4), 220-228. [ Links ]
Genini, J., Galetti, M., & Morellato, L. P. C. (2009). Fruiting phenology of palms and trees in an Atlantic rainforest land-bridge island. Flora-Morphology, Distribution, Functional Ecology of Plants, 204(2), 131-145. [ Links ]
Giombini, M. I., Bravo, S. P., & Martínez, M. F. (2009). Seed dispersal of the palm Syagrus romanzoffiana by tapirs in the semi-deciduous Atlantic forest of Argentina. Biotropica, 41(4), 408-413. [ Links ]
Goudel, F., Shibata, M., Coelho, C. M. M., & Miller, P. R. M. (2013). Fruit biometry and seed germination of Syagrus romanzoffiana (Cham.) Glassm. Acta Botanica Brasilica, 27(1), 147-154. [ Links ]
Guix, J. C., & Ruiz, X. (2000). Plant-disperser-pest evolutionary triads: how widespread are they. Orsis, 15, 121-126. [ Links ]
Honda, Y. (2008). Ecological correlations between the persistence of the soil seed bank and several plant traits, including seed dormancy. Plant Ecology, 196(2), 301-309. [ Links ]
Hu, X. W., Wang, Y. R., & Wu, Y. P. (2009). Effects of the pericarp on imbibition, seed germination, and seedling establishment in seeds of Hedysarum scoparium Fisch. et Mey. Ecological Research, 24(3), 559-564. [ Links ]
Jensen, K. (2004). Dormancy patterns, germination ecology, and seed-bank types of twenty temperate fen grassland species. Wetlands, 24(1), 152-166. [ Links ]
Long, R. L., Gorecki, M. J., Renton, M., Scott, J. K., Colville, L., Goggin, D. E., & Finch-Savage, W. E. (2015). The ecophysiology of seed persistence: a mechanistic view of the journey to germination or demise. Biological Reviews, 90(1), 31-59. [ Links ]
Lorenzi, H., Souza, H. M., Medeiros-Costa, J. T., Cerqueira, L. S. C., & Behr, N. (1996). Palmeiras no Brasil: nativas e exóticas [Palm trees in Brazil: native and exotic]. Nova Odessa, São Paulo: Plantarum. [ Links ]
Ministério da Agricultura, Pecuária e Abastecimento, Brasil. (2009). Regras para análise de sementes [Rules for seed testing]. Brasília, Brasil: Mapa/ACS. [ Links ]
Moles, A. T., Warton, D. I., & Westoby, M. (2003). Seed size and survival in the soil in arid Australia. Austral Ecology, 28(5), 575-585. [ Links ]
Neves, S. C., Ribeiro, L. M., Da Cunha, I. R. G., Pimenta, M. A. S., Mercadante-Simões, M. O., & Lopes, P. S. N. (2013). Diaspore structure and germination ecophysiology of the babassu palm Attalea vitrivir. Flora-Morphology, Distribution, Functional Ecology of Plants, 208(1), 68-78. [ Links ]
Orozco-Segovia, A., Batis, A. I., Roja-Aréchiga, M, & Mendoza, A. (2003). Seed biology of palms: a review. Palms, 47(2), 79-94. [ Links ]
Pérez, E., González, G. C., & Irabién, L. M. C. (2005). Germinación y supervivencia de semillas de Thrinax radiata (Arecaceae), una especie amenazada en la Península de Yucatán [Germination and survival of Thrinax radiata (Arecaceae) seeds, a threatened species in the Yucatan Peninsula]. Boletín de la Sociedad Botánica de México, 77, 9-20. [ Links ]
Ribeiro, L. M., Garcia, Q. S., Oliveira, D. M. T., & Neves, S. C. (2010). Critérios para o teste de tetrazólio na estimativa do potencial germinativo em macaúba [Criteria for the tetrazolium test in estimating the germination potential in macaw palm]. Pesquisa Agropecuária Brasileira, 45(4), 361-368. [ Links ]
Ribeiro, L. M., Souza, P. P., Rodrigues-Jr, A. G., Oliveira, T. G. S., & Garcia, Q. S. (2011). Overcoming dormancy in macaw palm diaspores, a tropical species with potential for use as bio-fuel. Seed Science and Technology, 39(2), 303-317. [ Links ]
Ribeiro, L. M., Oliveira, T. G. S., Carvalho, V. S., Silva, P. O., Neves, S. C., & Garcia, Q. S. (2012). The behaviour of macaw palm (Acrocomia aculeata) seeds during storage. Seed Science and Technology, 40(3), 344-353. [ Links ]
Schlindwein, G. (2012). Implicações ecológicas da dormência de sementes em Butia odorata Arecaceae Ecological implications of seed dormancy in Butia odorata Arecaceae Doctoral dissertation. Universidade Federal do Rio Grande do Sul, Brasil. [ Links ]
Thompson, K. (1997). The soil seed banks of North West Europe: methodology, density and longevity (vol. 1). Cambridge: Cambridge University Press. [ Links ]
Thompson, K., Ceriani, R. M., Bakker, J. P., & Bekker, R. M. (2003). Are seed dormancy and persistence in soil related? Seed Science and Research, 13(2), 97-100. [ Links ]
Vázquez-Yanes, C., & Orozco-Segovia, A. (1993). Patterns of seed longevity and germination in the tropical rainforest. Annual Review of Ecology and Systematics, 24, 69-87. [ Links ]
Baskin, C. C., & Baskin, J. M. (2014). Seeds: Ecology, biogeography, and evolution of Dormancy and germination (second edition). San Diego, CA, USA: Academic Press. [ Links ]
Bernacci, L. C., Martins, F. R., & Santos, F. A. M. (2008). Estrutura de estádios ontogenéticos em população nativa da palmeira Syagrus romanzoffiana (Cham.) Glassman (Arecaceae) [Structure of ontogenetic stages in the native population of the palm tree Syagrus romanzoffiana (Cham.) Glassman (Arecaceae)]. Acta Botanica Brasilica, 22(1), 119-130. [ Links ]
Brancalion, P. H. S. A., Novembre, A. D. L. C., & Rodrigues, R. R. (2011). Seed development, yield and quality of two palm species growing in different tropical forest types in SE Brazil: implications for ecological restoration. Seed Science and Technology, 39(2), 412-424. [ Links ]
Broschat, T. K., & Donselman, H. (1987). Effects of fruit maturity, storage, presoaking, and seed cleaning on germination in three species of palms. Journal of Environmental Horticulture, 5(1), 6-9. [ Links ]
Broschat, T. K. (1998). Endocarp removal enhances Butia capitata (Mart.) Becc. (pindo palm) seed germination. Horttechnology, 8(4), 586-587. [ Links ]
Coimbra, M. C., & Jorge, N. (2011). Proximate composition of guariroba (Syagrus oleracea), jerivá (Syagrus romanzof?ana) and macaúba (Acrocomia aculeata) palm fruits. Food Research International, 44(7), 2139-2142. [ Links ]
Dickie, J. B., Balick, M. J., & Linington, I. M. (1992). Experimental investigations into the feasibility of ex situ preservation of palm seeds; an alternative strategy for biological conservation of this economically important plant family. Biodiversity & Conservation, 1(2), 112-119. [ Links ]
Falasca, S. L., Del Fresno, C. M., & Ulberich, C. (2012). Possibilities for growing queen palm (Syagrus romanzofianna) in Argentina as a biodiesel producer under semi-arid climate conditions. International Journal of Hydrogen Energy, 37(19), 14843-14848. [ Links ]
Fenner, M. (1992). Seeds: The ecology of regeneration in plant communities. Wallingford, UK: CABI Publishing. [ Links ]
Freire, C. C., Closel, M. B., Hasui, E., & Ramos, F. N. (2013). Reproductive phenology, seed dispersal and seed predation in Syagrus romanzoffiana in a highly fragmented landscape. Annales Botanici Fennici, 50(4), 220-228. [ Links ]
Genini, J., Galetti, M., & Morellato, L. P. C. (2009). Fruiting phenology of palms and trees in an Atlantic rainforest land-bridge island. Flora-Morphology, Distribution, Functional Ecology of Plants, 204(2), 131-145. [ Links ]
Giombini, M. I., Bravo, S. P., & Martínez, M. F. (2009). Seed dispersal of the palm Syagrus romanzoffiana by tapirs in the semi-deciduous Atlantic forest of Argentina. Biotropica, 41(4), 408-413. [ Links ]
Goudel, F., Shibata, M., Coelho, C. M. M., & Miller, P. R. M. (2013). Fruit biometry and seed germination of Syagrus romanzoffiana (Cham.) Glassm. Acta Botanica Brasilica, 27(1), 147-154. [ Links ]
Guix, J. C., & Ruiz, X. (2000). Plant-disperser-pest evolutionary triads: how widespread are they. Orsis, 15, 121-126. [ Links ]
Honda, Y. (2008). Ecological correlations between the persistence of the soil seed bank and several plant traits, including seed dormancy. Plant Ecology, 196(2), 301-309. [ Links ]
Hu, X. W., Wang, Y. R., & Wu, Y. P. (2009). Effects of the pericarp on imbibition, seed germination, and seedling establishment in seeds of Hedysarum scoparium Fisch. et Mey. Ecological Research, 24(3), 559-564. [ Links ]
Jensen, K. (2004). Dormancy patterns, germination ecology, and seed-bank types of twenty temperate fen grassland species. Wetlands, 24(1), 152-166. [ Links ]
Long, R. L., Gorecki, M. J., Renton, M., Scott, J. K., Colville, L., Goggin, D. E., & Finch-Savage, W. E. (2015). The ecophysiology of seed persistence: a mechanistic view of the journey to germination or demise. Biological Reviews, 90(1), 31-59. [ Links ]
Lorenzi, H., Souza, H. M., Medeiros-Costa, J. T., Cerqueira, L. S. C., & Behr, N. (1996). Palmeiras no Brasil: nativas e exóticas [Palm trees in Brazil: native and exotic]. Nova Odessa, São Paulo: Plantarum. [ Links ]
Ministério da Agricultura, Pecuária e Abastecimento, Brasil. (2009). Regras para análise de sementes [Rules for seed testing]. Brasília, Brasil: Mapa/ACS. [ Links ]
Moles, A. T., Warton, D. I., & Westoby, M. (2003). Seed size and survival in the soil in arid Australia. Austral Ecology, 28(5), 575-585. [ Links ]
Neves, S. C., Ribeiro, L. M., Da Cunha, I. R. G., Pimenta, M. A. S., Mercadante-Simões, M. O., & Lopes, P. S. N. (2013). Diaspore structure and germination ecophysiology of the babassu palm Attalea vitrivir. Flora-Morphology, Distribution, Functional Ecology of Plants, 208(1), 68-78. [ Links ]
Orozco-Segovia, A., Batis, A. I., Roja-Aréchiga, M, & Mendoza, A. (2003). Seed biology of palms: a review. Palms, 47(2), 79-94. [ Links ]
Pérez, E., González, G. C., & Irabién, L. M. C. (2005). Germinación y supervivencia de semillas de Thrinax radiata (Arecaceae), una especie amenazada en la Península de Yucatán [Germination and survival of Thrinax radiata (Arecaceae) seeds, a threatened species in the Yucatan Peninsula]. Boletín de la Sociedad Botánica de México, 77, 9-20. [ Links ]
Ribeiro, L. M., Garcia, Q. S., Oliveira, D. M. T., & Neves, S. C. (2010). Critérios para o teste de tetrazólio na estimativa do potencial germinativo em macaúba [Criteria for the tetrazolium test in estimating the germination potential in macaw palm]. Pesquisa Agropecuária Brasileira, 45(4), 361-368. [ Links ]
Ribeiro, L. M., Souza, P. P., Rodrigues-Jr, A. G., Oliveira, T. G. S., & Garcia, Q. S. (2011). Overcoming dormancy in macaw palm diaspores, a tropical species with potential for use as bio-fuel. Seed Science and Technology, 39(2), 303-317. [ Links ]
Ribeiro, L. M., Oliveira, T. G. S., Carvalho, V. S., Silva, P. O., Neves, S. C., & Garcia, Q. S. (2012). The behaviour of macaw palm (Acrocomia aculeata) seeds during storage. Seed Science and Technology, 40(3), 344-353. [ Links ]
Schlindwein, G. (2012). Implicações ecológicas da dormência de sementes em Butia odorata Arecaceae Ecological implications of seed dormancy in Butia odorata Arecaceae Doctoral dissertation. Universidade Federal do Rio Grande do Sul, Brasil. [ Links ]
Thompson, K. (1997). The soil seed banks of North West Europe: methodology, density and longevity (vol. 1). Cambridge: Cambridge University Press. [ Links ]
Thompson, K., Ceriani, R. M., Bakker, J. P., & Bekker, R. M. (2003). Are seed dormancy and persistence in soil related? Seed Science and Research, 13(2), 97-100. [ Links ]
Vázquez-Yanes, C., & Orozco-Segovia, A. (1993). Patterns of seed longevity and germination in the tropical rainforest. Annual Review of Ecology and Systematics, 24, 69-87. [ Links ]
1. Universidade Federal de Minas Gerais, Departamento de Botânica, Laboratório de Fisiologia Vegetal, Av. Presidente Antônio Carlos, 6627, Belo Horizonte, Brasil; tuliogsoliveira@gmail.com
2. Universidade Federal de Lavras, Departamento de Ciências Florestais, Laboratório de Sementes Florestais, Campus Universitário, Lavras, Brasil; acjose@dcf.ufla.br, jmfaria@dcf.ufla.br
3. Universidade Estadual de Montes Claros, Departamento de Biologia Geral, Laboratório de Micropropagação, Campus Universitário Professor Darcy Ribeiro, Montes Claros, Brasil; leomrib@hotmail.com
Received 13-VIII-2014. Corrected 08-XI-2014. Accepted 04-XII-2014.


{kind=link}
{kind=link}