










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Diversidad de escarabajos necrófilos (Coleoptera: Scarabaeidae, Silphidae, Staphylinidae y Trogidae) en una región semiárida del valle de Zapotitlán de las Salinas, Puebla, México
Necrophilous beetles diversity (Coleoptera: Scarabaeidae, Silphidae, Staphylinidae y Trogidae) in a semiarid area of Zapotitlán de las Salinas valley, Puebla, Mexico
Necrophilous beetles diversity (Coleoptera: Scarabaeidae, Silphidae, Staphylinidae y Trogidae) in a semiarid area of Zapotitlán de las Salinas valley, Puebla, Mexico
*Dirección para correspondencia:
Abstract
Mexico is constituted of arid and semiarid areas in more than half of its land extension, where most of their resources are available only during the rainy season. For those species that recycle resources, such as the carrion fauna, this represents a highly active season; however, the biological and diversity patterns of carrion fauna are poorly understood in these areas. Here, we studied the abundance, diversity and richness of the Scarabaeidae, Silphidae, Staphylinidae and Trogidae families in the Zapotitlán de las Salinas Valley in Puebla, Mexico. Over a one-year period, monthly samples were collected from five different vegetation systems that included the scrublands, a columnar cactus landscape, and altered vegetation. Samples were collected with the use of NTP-80 traps baited with squid, and data on abundance and richness were obtained and evaluated, with respect to monthly precipitation and sampling site location. We collected a total of 613 insects from 12 genera and 15 species. Across systems, Staphylinidae showed the greatest richness (nine species) and abundance (74.2%), followed by Scarabaeidae (21.9%), Silphidae (2.9%) and Trogidae (1%). Significant differences were observed between sites and months; nevertheless, no relationship was observed between abundance and temperature or precipitation. Across taxonomic groups, abundance and richness were most evident for scrub sites, while a greater abundance during the dry season was obtained. The results of this study indicated that the carrion faunal community composition is closely related to the type of vegetation and did not depend on the rainy season. In spite that lesser carrion fauna was observed in this area when compared to other regions of Mexico, this report constitutes a significant contribution to our understanding of the ecological role of this fauna in arid areas.
Key words: carrion beetles, NTP-80, seasonality, scrub, columnar cactus.
Resumen
Las zonas áridas y semiáridas ocupan más de la mitad del territorio de México, sin embargo, son ambientes donde los escarabajos necrófilos han sido escasamente estudiados. En esta investigación se presenta la diversidad y fenología de Scarabaeidae, Silphidae, Staphylinidae y Trogidae en el valle de Zapotitlán de las Salinas, Puebla. Se realizaron muestreos mensuales durante un año de abril de 1998 a marzo de 1999 empleando trampas tipo NTP-80 cebadas con calamar, distribuidas en un mezquital y matorral espinoso, en cactáceas columnares y vegetación alterada. Se capturaron 613 individuos de 12 géneros y 15 especies. Staphylinidae tuvo la mayor riqueza (9 especies) y abundancia (74.2%), le siguieron Scarabaeidae (21.9%), Silphidae (2.9%) y Trogidae (1%) con dos especies cada una. La fauna para las dos primeras fue inferior a la observada en otras zonas de México. La máxima abundancia y riqueza estuvieron en la época seca y las comunidades de escarabajos necrófilos prefirieron los sitios con mezquital y matorral, donde permanecieron más tiempo y se registraron todas las especies, por lo tanto los tipos de vegetación presentes fueron el factor más importante que determinó las variaciones locales de abundancia, diversidad y riqueza y no la época de lluvias.
Palabras clave: necrófagos, NTP-80, estacionalidad, matorral xerófilo, cactácea columnar.
Las zonas áridas y semiáridas ocupan más de la mitad del territorio de México, y están cubiertas en su mayor parte, por diversos tipos de comunidades vegetales entre las que se encuentran el matorral y el pastizal xerófilo (Rzedowski 1978, Valiente-Banuet et al. 1991). Como cualquier otra comunidad vegetal, estos tipos de vegetación tienen diferentes estratos y una fauna característica, que se ha adaptado a las condiciones climáticas que la limita, y han promovido el desarrollo de asociaciones más estrechas entre plantas y animales (Muller 1953, Fowler 1986, McAuliffe 1986). Las lluvias en las zonas áridas son ausentes o puntuales en el tiempo; Halvorson & Duncan (1975) han señalado que una época de lluvias marcada puede indicar el aumento de la actividad y la productividad biológica. Asimismo, Beatley (1974) y Whitford (1978), encontraron que en este periodo se estimula la sincronización en los procesos de alimentación, reproducción y crecimiento de los organismos, debido a que los recursos son más abundantes en tiempo y espacio.
Uno de los grupos que habitan estas áreas son los escarabajos necrófilos. En este grupo se encuentran los de las familias Scarabaeidae, Silphidae, Staphylinidae y Trogidae, que agrupan especies degradadoras y depredadoras, las cuales son comúnmente atraídas por los excrementos, o las plantas y animales en descomposición (Morón & Terrón 1984, Morón & López-Méndez 1985, Morón et al. 1986, Deloya et al. 1987, Terrón et al. 1991, Sánchez-Ramos et al. 1993, Acuña 2004, Navarrete-Heredia 2009, Trevilla-Rebollar et al. 2010, Jiménez-Sánchez et al. 2011). En general, esos recursos también están limitados en tiempo y espacio, lo que hace que sean de alta demanda, por lo que hay una gran competencia entre este tipo de insectos (Hansky 1987, Giller & Doube 1989, Kremen et al. 1993, Estrada et al. 1998).
La actividad de estos escarabajos es de suma importancia en la descomposición y el reciclaje de los nutrientes (Hansky 1983, Hortal-Muñoz et al. 2000), y por lo tanto, cumplen un papel vital en el óptimo funcionamiento de los ecosistemas. Para la comprensión de esa funcionalidad, es necesario contar con datos sobre la diversidad; desafortunadamente, los patrones de diversidad de este tipo de fauna han sido escasamente estudiados en las regiones áridas de México. Por lo anterior, este trabajo tiene como objetivo, conocer la diversidad y observar si hay una diferencia de la actividad en las poblaciones de escarabajos necrófilos de Scarabaeidae, Silphidae, Staphylinidae y Trogidae, entre los periodos de lluvias y de sequía, en la región semiárida del valle de Zapotitlán de las Salinas, Puebla, la cual forma parte de la reserva de la biosfera Tehuacán-Cuicatlán, México.
Materiales y Métodos
Área de estudio: El valle de Zapotitlán de las Salinas se encuentra enclavado en la porción occidental del valle de Tehuacán-Cuicatlán, el cual a su vez se localiza en el sureste del estado de Puebla y el noroeste de Oaxaca. La región presenta un clima del tipo semicálido (BShw”(w)(e)g) con una temperatura media anual de 21ºC y una canícula bien definida de marzo a mayo (García 1988). La precipitación media anual es de 400-450mm y de acuerdo con los registros históricos, la precipitación total de mayo a septiembre es de 340mm y mayor de 48mm en cada mes, lo que define la época de lluvias, mientras que la época de sequía va de octubre a abril, teniendo un total de 61.2mm y mensualmente menos de 19mm (SMN 2012). La altitud promedio es de 1 450msnm. La vegetación es un matorral xerófilo (Rzedowski 1978), con una flora principalmente neotropical de alrededor de 3 000 especies de plantas vasculares en todo el valle Tehucán-Cuicatlán (Rzedowski 1978, Villaseñor et al. 1990, Dávila et al. 1993, Montaña & Valiente-Banuet 1998). Posee igualmente, una importante estructura de comunidades vegetales endémicas (Arias et al. 2000, Camargo-Ricalde et al. 2002, Méndez et al. 2005). Valiente-Banuet et al. (2000) caracterizaron la vegetación de este valle y reconocieron 29 asociaciones en seis categorías con base en criterios fisionómico-estructurales.
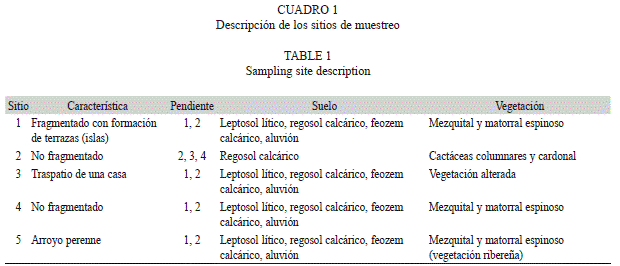
Trabajo de campo: Para el muestreo, fueron elegidos cinco sitios de acuerdo con las diferentes características de vegetación, tipo de suelo y pendiente (Cuadro 1), los cuales estuvieron separados aproximadamente 2km uno de otro, dentro de un área aproximada de 19 400km2 en el valle de Zapotitlán de las Salinas, entre los 18°18’ y 18°20’ N y los 97°27’ y 97°30’ W. La altitud de los sitios de muestreo fluctuó entre 1 400 y 1 500msnm.
Tres sitios se ubicaron en un mezquital combinado con matorral espinoso (Figs. 1A-C), el primer tipo de vegetación también denominado como selva baja perennifolia con espinas laterales estuvo dominado por Prosopis laevigata (Humb & Bonpl. ex Willd.) M. C. Johnston; mientras que el matorral espinoso de espinas laterales estuvo caracterizado por presentar arbustos espinosos, donde destacaron especies arbóreas y arbustivas caducifolias, con escasa o nula presencia de mezquite (Oliveros 2000). Un cuarto sitio se ubicó en un bosque de cactáceas columnares (Fig. 1D) de Neobuxbaumia tetetzo (F. A. C. Weber) Backeb. y el cardonal de Cephalocerus columna-trajani (Karw.) K. Schum. (Oliveros 2000). Por último, el quinto sitio con vegetación totalmente alterada, en el traspatio de una casa del poblado principal.
Los muestreos fueron mensuales en un ciclo anual de abril 1998 a marzo 1999, para estos se emplearon 15 necro trampas permanentes del tipo NTP-80, cebadas con calamar. Como líquido conservador se usó una mezcla de 95 partes de alcohol al 70% y cinco partes de ácido acético glacial (Morón & Terrón 1984). La trampa estuvo instalada todo el año; tanto el cebo como el líquido fueron sustituidos aproximadamente cada 30 días, y la muestra recuperada llevada al laboratorio para su separación. En cada sitio se instalaron dos trampas excepto en la vegetación alterada que tenía una extensión limitada, y fue el lugar donde se permitió colocar una trampa en el poblado principal.
Trabajo de laboratorio: La identificación de los ejemplares se realizó con las claves de Smetana (1995), Deloya (1995), Navarrete-Heredia et al. (2002) y Navarrete-Heredia (2009). Las especies fueron corroboradas por parte de especialistas como C. Deloya (Scarabaeidae y Trogidae) y J.L. Navarrete-Heredia (Staphyinidae y Silphidae). La subfamilia Aleocharinae (Staphylinidae) fue excluida del análisis por su dificultad taxonómica. Los ejemplares se depositaron en las siguientes colecciones entomológicas: Facultad de Estudios Superiores Iztacala (FESI), Universidad Nacional Autónoma de México, Estado de México, México y Centro de Estudios en Zoología (CZUG), Universidad de Guadalajara, Jalisco, México.
Para el análisis ecológico se cuantificó el número de individuos y la riqueza de especies por mes y por sitio; con estos valores se calculó el índice de diversidad de Shannon (H’) para cada sitio y época del año (sequía y lluvias), se aplicó la prueba de t para comparar la diversidad entre estos (Magurran 1989). También, se calculó el índice de Jaccard que utiliza datos de presencia-ausencia para obtener la similitud faunística entre los pares de sitios. Los datos fueron sometidos a un análisis de varianza de medidas repetidas con la finalidad de compensar la diferencia del número de trampas instaladas en la vegetación alterada (Zar 1984). Los valores de abundancia fueron transformados a rangos y el análisis se realizó sobre los rangos por medio de Proc Rank y Proc MIXED. El tratamiento de los datos se realizó a través de los programas PAST, versión 2.12 (Hammer et al. 2001) y SAS versión. 9.2 (SAS Inst. 1988). Además, debido a que la riqueza local obtenida en las muestras para grupos muy diversos como los insectos, subestima el número de especies real, se emplearon cinco estimadores de especies no paramétricos ACE, ICE, Chao 1, Jack 1 y Bootstrap, calculados con el programa EstimateS V. 8.2.0 (Colwell 2006).
Resultados
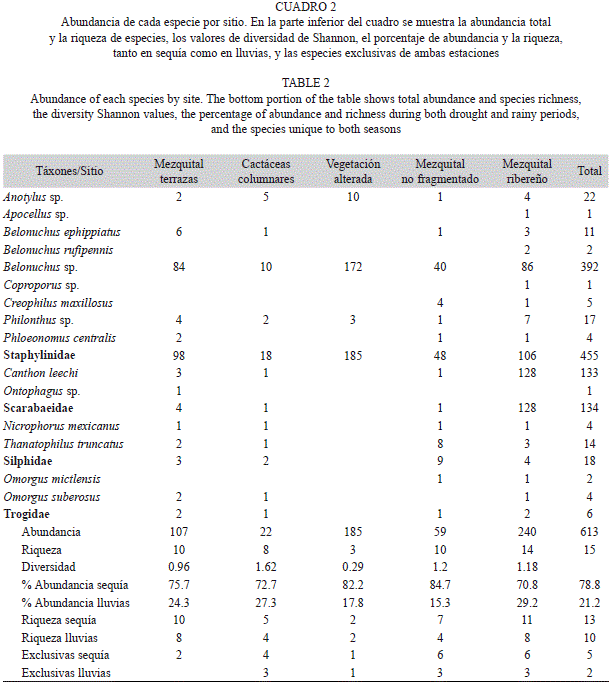
Se obtuvieron un total de 613 individuos agrupados en 12 géneros y 15 especies, de las cuales nueve se determinaron a nivel de especie y seis a género (Cuadro 2). La mayor riqueza de especies y géneros fue para Staphylinidae con nueve y siete respectivamente, mientras que Scarabaeidae, Silphidae y Trogidae estuvieron representados por dos especies y dos géneros, con excepción de Trogidae, cuyas especies pertenecieron a un solo género (Cuadro 2). Los estimadores de riqueza (ACE=17, ICE=18, Chao1=17, Jack1=19, Bootstrap=17) indicaron la existencia de entre 17 y 19 especies, lo que sugiere que faltarían por capturar entre dos y cuatro especies adicionales a las 15 registradas. La mayor cantidad de especies (13) se registró durante el periodo de sequía, mientras que en la de lluvias se obtuvieron 10 en toda el área. Una mayor riqueza en sequía se observó en cada uno de los sitios, excepto en el sitio de vegetación alterada, que tuvo dos especies en cada período (Cuadro 2).
En relación con la riqueza específica en cada uno de los sitios de muestreo, el mezquital ribereño registró el mayor valor con 14, le siguieron el mezquital no fragmentado y el mezquital de terrazas con 10 cada uno, las cactáceas columnares con ocho, y la menor riqueza se encontró en la vegetación alterada con solo tres. El mezquital de terrazas y ribereño fueron los únicos con especies exclusivas, además la fauna de estos dos lugares en conjunto agrupó las 15 especies registradas en este estudio. Apocellus sp., Belonuchus rufipennis (Fabricius, 1801) y Coproporus sp. Se capturaron únicamente en el mezquital ribereño y Ontophagus sp. en el mezquital de terrazas; Anotylus sp., Belonuchus sp. y Philonthus sp. se encontraron en todos los sitios. Por su parte, Belonuchus ephippiatus (Say, 1830), Canthon leechi (Martínez et al., 1964), Nicrophorus mexicanus Matthews, 1888 y Thanatophilus truncatus (Say, 1823) únicamente estuvieron ausentes en la vegetación alterada. Creophilus maxillosus (Linnaeus, 1758) y Omorgus mictlensis Deloya, 1995 exclusivamente se encontraron en el mezquital no fragmentado y el ribereño, Phloeonomus centralis Blackwelder, 1944 en los tres mezquitales, y Omorgus suberosus Fabricius, 1775 en las cactáceas columnares, el mezquital de terrazas y ribereño (Cuadro 2).
El mezquital de terrazas presentó la mayor riqueza de especies, las cuales estuvieron presentes todo el año con excepción de abril; tuvieron dos incrementos, el primero en septiembre, donde coexistieron siete de las diez especies registradas en este sitio, y el segundo en diciembre, donde coincidieron nueve especies. Ocho de éstas estuvieron presentes tanto en lluvias como en sequía, las restantes dos fueron N. mexicanus y B. ephippiatus, encontradas exclusivamente en enero y de diciembre a marzo respectivamente; la primera tuvo la misma fenología en todos los sitios donde se le capturó presentándose en enero o febrero. Le siguió el mezquital ribereño que albergó cinco de catorce especies tanto en lluvias como en sequía, y las otras nueve restringidas a alguno de estos periodos. Mientras que en las cactáceas columnares, la vegetación alterada y el mezquital no fragmentado, las especies mostraron preferencia por alguna estación del año; con excepción de Belonuchus sp. en la vegetación alterada y el mezquital no fragmentado, y Anotylus sp. en las cactáceas columnares, que estuvieron activas en ambos periodos.
En las cactáceas columnares y la vegetación alterada, la riqueza mensual varió entre una y dos, por lo que no se observaron valores significativos. Algo similar ocurrió en el mezquital no fragmentado, donde sus diez especies se distribuyeron a lo largo del muestreo entre una y cuatro especies por mes, presentes de junio a marzo, con excepción en febrero. Por último, en el mezquital ribereño las especies estuvieron presentes todo el año con excepción de abril, y su número por mes osciló de una a cinco (este valor máximo se registró en septiembre y octubre).
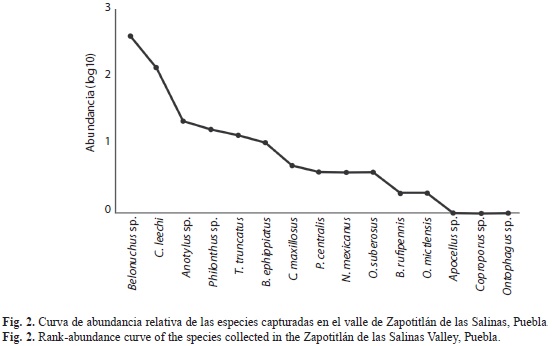
La familia con mayor abundancia relativa fue Staphylinidae con 74.2%, seguida por Scarabaeidae con 21.9%, Silphidae con 2.9% y Trogidae con 1%. Belonuchus sp. y C. leechi agruparon el 85.6% de la abundancia total, y las 13 especies restantes el 14.4%, todas con menos de 22 individuos (Fig. 2).
La abundancia presentó diferencias significativas entre los sitios de muestreo (ANOVA, p=0.0493). La mayor abundancia se obtuvo en el mezquital ribereño con 39.1%, seguido por la vegetación alterada con 30.1%, el mezquital de terrazas con 17.5%, el mezquital no fragmentado con 9.6% y las cactáceas columnares, donde se registró la menor abundancia con 3.6% individuos (Cuadro 2).
Se observó una alta dominancia de Belonuchus sp. en cuatro de los sitios, con más del 40% y hasta el 93% (vegetación alterada) de la abundancia de cada sitio; esta especie solo fue superada en el mezquital ribereño por C. leechi con 53.3%, valor que contrasta fuertemente con uno a tres individuos capturados de esta especie en los otros sitios, con excepción de la vegetación alterada donde estuvo ausente. El resto de las especies estuvieron representadas por menos de 12 individuos en todos los sitios (Cuadro 2).
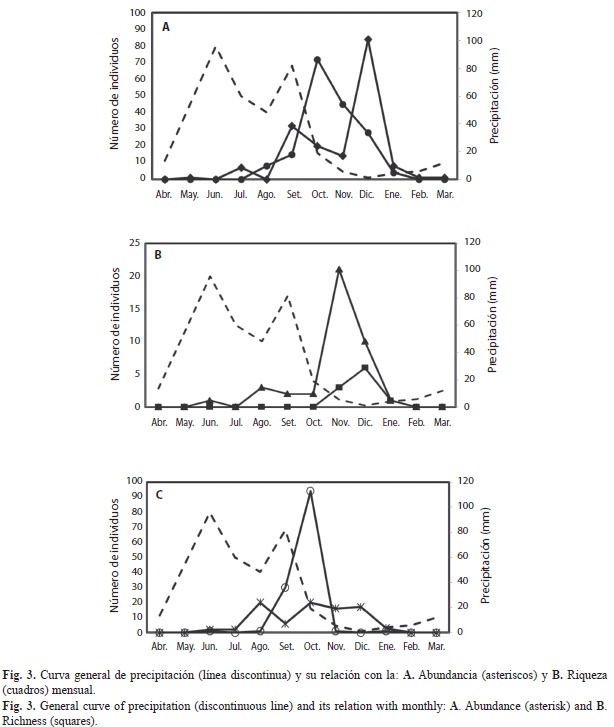
No se observó una relación directa entre la abundancia y la precipitación (ANOVA, p=0.2189), pero sí entre los meses evaluados a lo largo del año (ANOVA, p=0.001). La curva general de la abundancia mensual tuvo un incremento máximo en octubre, desfasados de las máximas precipitaciones (Fig. 3A), al igual que la riqueza mensual de especies, que mostró el valor más alto en octubre (nueve especies) y otro en enero (ocho) (Fig. 3B).
En el periodo de sequía se capturó la mayor abundancia (78.8%). Este patrón general se repitió en cada uno de los sitios, donde más del 70% de los individuos se capturaron durante el período de la sequía, y solo entre un 15% y 29% durante las lluvias (Cuadro 2).
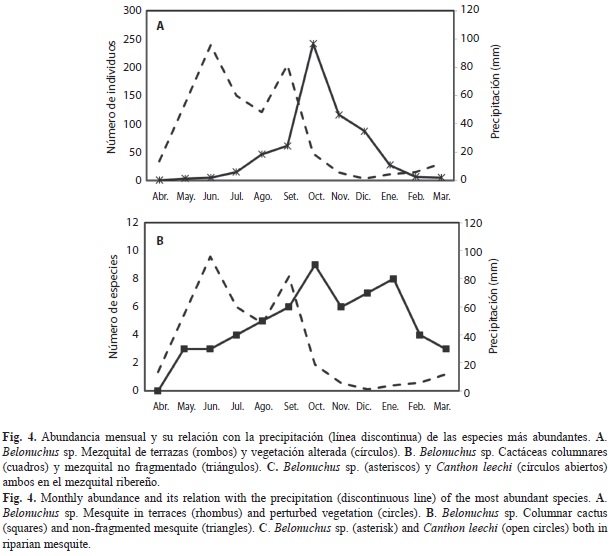
Staphylinidae estuvo presente 11 meses de mayo a marzo, siendo la más activa de las cuatro familias; mostró dos valores máximos de abundancia uno en agosto y uno mayor en octubre (Fig. 4A); Scarabaeidae estuvo en seis meses y su mayor abundancia fue en octubre (Fig. 4B). Silphidae y Trogidae tuvieron de uno a seis individuos por mes y por lo tanto no mostraron valores de abundancia importantes (4C).
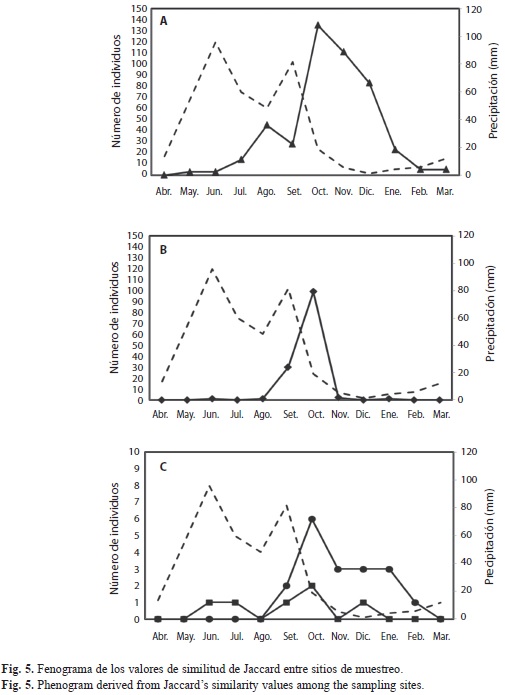
Belonuchus sp. y C. leechi fueron las especies más abundantes de agosto a diciembre y de septiembre a octubre, respectivamente, ambas mostraron el mismo patrón que el de sus familias correspondientes Staphylinidae y Scarabaeidae; sin embargo, el número de valores máximos de la abundancia de Belonuchus sp. fue diferente en cada sitio: en el mezquital de terrazas tuvo un valor máximo en septiembre y otro en diciembre, en la vegetación alterada y en el mezquital no fragmentado se observó un valor máximo en cada sitio, en octubre y noviembre, respectivamente (Fig. 5A y B); en las cactáceas columnares fue poco abundante y solo se presentó de noviembre a enero y en el mezquital ribereño se observaron cuatro valores máximos similares en agosto y de octubre a diciembre (Fig. 5B y C). Por su parte, el 96% de la abundancia de C. leechi se recolectó en el mezquital ribereño con un valor máximo en octubre (5C).
La abundancia por especie también mostró diferencias significativas a lo largo del año (ANOVA, p<0.0001). De manera general, la especie dominante Belonuchus sp.l estuvo activa durante 11 meses, le siguió Philonthus sp. con siete, C. leechi y Anotylus sp. con seis cada una, el resto de las especies estuvieron activas en menos de cinco meses. Sin embargo, en cada sitio el periodo de actividad fue menor, por ejemplo la actividad de Belonuchus sp. fue de seis a nueve meses en todos los sitios, excepto en las cactáceas columnares donde solo estuvo en tres meses, C. leechi fue la segunda más activa pero solo en el mezquital ribereño con seis meses. Para las demás especies la actividad fue inferior a cuatro meses en todos los sitios.
No hubo diferencias significativas entre el valor de diversidad obtenido para el periodo de lluvias (H’=1.39) y el período de sequía (H’=1.09) (prueba de student, p=0.001). Al comparar entre estaciones del año por sitio, se encontró que la vegetación alterada fue la única que mostró diferencias significativas (prueba de student, p=0.001). Por el contrario, al contrastar la diversidad entre los sitios, usando las posibles combinaciones entre trampas individuales, ninguna tuvo diferencias significativas (prueba de student, p=0.001).
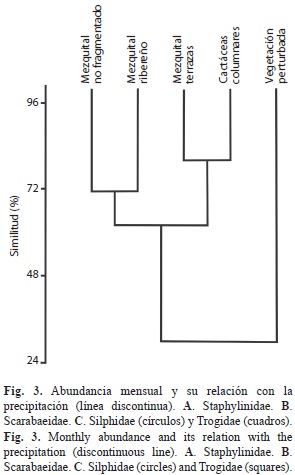
El índice de similitud de Jaccard produjo un fenograma con dos agrupamientos (Fig. 6). Uno de ellos incluyó a cuatro sitios con un porcentaje de similitud de 63%, es decir que entre ellos compartieron a más de la mitad de las especies entre siete y diez, a su vez dentro de éste hubo dos subconjuntos: uno formado por el mezquital de terrazas y las cactáceas columnares con 80% de similitud, donde este último tipo de vegetación compartió todas sus especies con el mezquital de terrazas. El otro subconjunto que incluyó al mezquital no fragmentado y al ribereño, con 72% y diez especies en común, la vegetación alterada solo tuvo tres especies, todas ellas distribuidas en todos los sitios, lo que representó un valor de 30%.
Discusión
El valle de Zapotitlán de las Salinas presentó una fauna necrófila inferior a la observada en otras zonas de México. La riqueza obtenida de Staphylinidae en el matorral xerófilo de Zapotitlán, fue considerablemente menor a la observada en estudios llevados a cabo en otros tipos de vegetación, como los bosques tropical caducifolio y subcaducifolio, pino-encino, pino y mesófilo de montaña con entre 22 y 53 especies (Ruíz-Lizárraga 1993, Jiménez-Sánchez et al. 2000, 2001, Caballero 2003, Márquez 2003, Jiménez-Sánchez et al. 2011). La riqueza obtenida es comparable solamente con las 12 especies obtenidas por Cejudo-Espinosa & Deloya (2005) en un bosque de pino a 3 600m de altitud y las 14 registradas por Morales et al. (1998) en vegetación costera, las cuales representan los puntos extremos de distribución altitudinal.
La riqueza de especies de Scarabaeidae es por ahora la más baja registrada en el país con el uso de la NTP-80, valores bajos habían sido registrados en áreas alteradas con registros entre cuatro y cinco especies (Morón 1987, Morales et al. 1998, Rivera-Cervantes & García-Real 1998, Trevilla-Rebollar et al. 2010). En otros estudios, la mayor riqueza se ha registrado en el bosque tropical caducifolio con hasta 25 especies y la menor en el bosque de pino con nueve, pasando por los bosques mesófilo de montaña, tropical perennifolio y subcaducifolio, y pino-encino (Morón et al. 1986, Deloya et al. 1987, Morón 1987, Terrón et al. 1991, Deloya 1992, Deloya & Morón 1998, Navarrete-Heredia & Quiroz-Rocha 2000, Reyes 2001, Cerecero 2002, Méndez-Castellanos 2002, Quiroz-Rocha et al. 2008, Mora-Aguilar & Montes de Oca 2009, Trevilla-Rebollar et al. 2010).
Para el caso de Silphidae frecuentemente son atraídas entre dos y cuatro especies, el valor mayor se ha obtenido en bosques de pino, pino-encino y mesófilo de montaña (Terrón et al. 1991, Arellano 1998, Rivera-Cervantes & García-Real 1998, Navarrete-Heredia & Quiroz-Rocha 2000, Reyes 2001, Méndez-Castellanos 2002, Cejudo-Espinosa & Deloya 2005, Quiroz-Rocha et al. 2008, Trevilla-Rebollar et al. 2010).
Con relación a Trogidae, son atraídas de una a cuatro especies, la mayor riqueza ha sido registrada en el bosque tropical caducifolio y la menor en bosques de pino, pino-encino y vegetación costera (Deloya et al. 1987, Terrón et al. 1991, Deloya 1992, Deloya & Morón 1998, Morales et al. 1998, Reyes 2001, Méndez-Castellanos 2002, Mora-Aguilar & Montes de Oca 2009, Trevilla-Rebollar et al. 2010).
En cuanto a la abundancia, las cuatro familias estudiadas han sido ampliamente recolectadas en la trampa NTP-80. Es Staphylinidae el grupo más representativo en la mayoría de los casos, seguido de Scarabaeidae, Silphidae y Trogidae (Morón & Terrón 1984, Morón & López-Méndez 1985, Terrón et al. 1991, Morales et al. 1998, Jiménez-Sánchez et al. 2000, Méndez-Castellanos 2002, Acuña 2004). Aunque, la presencia de algunas de estas familias en los estudios puede estar limitada por su distribución geográfica restringida y por la altitud (Morón et al. 1986, Cejudo-Espinosa & Deloya 2005).
Por otro lado, Belonuchus sp. (Staphylinidae) y C. leechi (Scarabaeidae) agruparon el 85.6% de la abundancia total. La dominancia de pocas especies es un patrón ampliamente documentado para estas familias (Jiménez-Sánchez et al. 2000, 2001, Navarrete-Heredia & Quiroz-Rocha 2000, Márquez 2003, Caballero et al. 2009, Trevilla-Rebollar et al. 2010). Así, éste comportamiento sugiere que estas especies son altamente tolerantes a las condiciones de clima y vegetación de la zona. Rivera-Cervantes & Halffter (1999) señalan que C. leechi se encuentra en áreas con insolación, lo cual es una característica del área de estudio.
La época de lluvias en el valle de Zapotitlán, no fue un factor determinante para una mayor abundancia de la fauna necrófila como podría esperarse. En los trópicos la variación de la precipitación, especialmente en el periodo seco puede tener un efecto considerable sobre la abundancia de los insectos provocando un bajo número de adultos activos (Wolda 1978) lo que afecta directamente en la disminución de las poblaciones de las familias necrófilas (Ruíz-Lizárraga 1993, Jiménez-Sánchez et al. 2000, Reyes 2001, Méndez-Castellanos 2002, Márquez 2003, Trevilla-Rebollar et al. 2010, Jiménez-Sánchez et al. 2011).
Las especies no se encuentran limitadas por las lluvias, éstas estuvieron presentes tanto en la época húmeda como en la seca, además no hubo diferencias entre la diversidad de ambos periodos. Lobo et al. (2006), hicieron un estudio en una zona árida en la región central de España, donde encontraron una afinidad de la fauna de Scarabaeidae hacia los recursos y no hacia la fluctuación de humedad. Caballero et al. (2009) observaron cambios estacionales marcados en la diversidad de la comunidad de Staphylinidae necrófilos entre la estación seca y húmeda, sin embargo, no ocurrió lo mismo para la comunidad de coprófilos, debido a la disponibilidad continua de excremento de ganado en los sistemas de agro-pastoreo de una región de Chiapas, México.
Las variaciones locales de la abundancia, la diversidad y la riqueza de estos escarabajos necrófilos, posiblemente hayan sido determinadas por las características del tipo de vegetación, como los periodos de floración y fructificación de algunos de los elementos del matorral xerófilo que durante las lluvias sólo almacenan nutrientes suficientes para la reproducción la cual ocurre al final de dicho período (Arias et al. 2000). Por ejemplo, la floración de las cactáceas columnares N. tetetzo ocurre de marzo a mayo (Valiente-Banuet et al. 1996). Numa et al. (2012) señalan que la vegetación es uno de los principales factores que propician la variabilidad dentro de la abundancia, debido a que ésta determina que la fauna pueda encontrar más recursos a lo largo del año. En este sentido, las zonas áridas están caracterizadas como sitios de baja productividad (Hadley & Szarek 1981), donde los organismos deben buscar de manera activa y continua los recursos presentes en los diferentes tipos de vegetación.
Las diferentes asociaciones vegetales presentes en cada zona mostraron una riqueza de especies y una abundancia diferente en cada sitio. La distribución observada, indica que los escarabajos prefieren los mezquitales y matorrales establecidos en las zonas de depósito aluvial donde se desarrolla la principal actividad productiva humana. En particular el mezquital de terrazas y el ribereño fueron los lugares que albergaron en conjunto a las 15 especies registradas en este estudio, varias de ellas exclusivas de estos sitios. Estrada et al. (1998) menciona que las zonas de transición son las que presentan una diversidad más grande. De estos dos lugares, las terrazas mantuvieron mayor riqueza durante todo el año, mientras que el ubicado en las inmediaciones del río perenne, constituyó un punto de interacción de las faunas de las distintas áreas muestreadas, lo cual podría indicar la importancia de la humedad para el desarrollo de estos grupos de coleópteros. Aunque con grupos de escarabajos diferentes a los aquí tratados, Zaragoza-Caballero et al. (2003) y Jiménez-Sánchez et al. (2009) observaron que la presencia constante de un cuerpo de agua artificial (presa) influyó en el mantenimiento de las poblaciones de algunas especies de Lampyridae y Staphylinidae durante diez u 11 meses en un bosque tropical caducifolio.
En el bosque de cactáceas columnares se encontraron especies que también estuvieron presentes en los sitios con mezquital y matorral, así como, aquellas ampliamente distribuidas. Esto coincide con el hecho de que más de la mitad de las especies se comparten entre cuatro de los sitios ubicados en el mezquital y las cactáceas columnares debido a la cercanía de los mismos, mientras que el sitio con la vegetación alterada fue el lugar más disímil, donde solo se encontraron a las especies más tolerantes por estar en todos los sitios que fueron: Anotylus sp., Belonuchus sp. y Philonthus sp.
En cuanto a algunos aspectos biológicos de las especies encontradas, se sabe que C. maxillosus acude a carroña de varios tipos donde se alimenta de larvas, se encuentra en hábitats naturales variados, así como, en situciones de sinantropía (Newton 2001), en Zapotitlán sólo fue capturada en el mezquital no fragmentado y el ribereño que correspondieron con los lugares más húmedos y donde se observó la mayor actividad humana. Además, su abundancia fue baja con sólo cinco individuos, lo cual concuerda con lo observado en otros estudios sobre estafilínidos necrófilos donde se registran menos de tres individuos en bosque tropical caducifolio y bosque mesófilo de montaña (Huacuja 1982, Márquez 2001, Jiménez-Sánchez et al. 2001), por lo que aparentemente sus poblaciones suelen ser bajas.
Por su parte, B. ephippiatus se encontró en todos los sitios con excepción de la vegetación alterada, se sabe que tiene preferencia por las zonas semiáridas, donde es comúnmente encontrada en varios tipos de cactáceas grandes y otras plantas en descomposición (Smetana 1995). En la misma área de estudio Jiménez-Sánchez & Padilla (1999) confirmaron esta aseveración al registrarla dentro de Neobuxbaumia tetetzo en descomposición buscando a sus presas y parece ser que la carroña es un recurso alternativo. Destacan también B. rufipennis y P. centralis, por haber tenido una abundancia muy baja con respecto a recolectas en localidades con otros tipos de vegetación y promedios de precipitación mayor donde han sido muy abundantes en la NTP-80 (Navarrete-Heredia 1996, Jiménez-Sánchez et al. 2000, Márquez 2003, Jiménez-Sánchez et al. 2011).
Canthon leechi acude tanto a coprotrampas como a trampas con carroña, se ha observado que tiene preferencia por la selva caducifolia, bordes de selva perennifolia y sitios abiertos (Rivera-Cervantes & Halffter 1999), esto último es una condición común en el área de estudio, aunque su captura se concentró en el mezquital ribereño.
Nicrophorus mexicanus y T. truncatus, coexistieron en el tiempo y estuvieron presentes en todos los sitios con excepción del sitio con vegetación alterada. Navarrete-Heredia & Fierros-López (2000) señalan que N. mexicanus explota cadáveres relativamente grandes y T. truncatus cadáveres de tallas pequeñas.
De acuerdo con su distribución estatal, N. mexicanus se registra por primera vez para Puebla y su presencia en el matorral xerófilo es poco común, pues se ha observado que tiene preferencia por las zonas montañosas en bosque de encino, encino-pino y pino, y mesófilo de montaña (Navarrete-Heredia & Fierros-López 2000); sin embargo, también han sido registrados algunos organismos en bosque tropical caducifolio (Gómez-Jaimes et al. 2004) lo cual sugiere que si bien tiene mayor afinidad por zonas montanas, la especie puede habitar otros ecosistemas. Por su parte, T. truncatus ha sido recolectada en una amplia variedad de ecosistemas desde bosque tropical caducifolio hasta el bosque de encino-pino, pino y mesófilo de montaña (Navarrete-Heredia & Fierros-López 2000) y al igual que N. mexicanus se registran por primera vez para un matorral xerófilo.
En el caso de Trogidae, O. suberosus es la más ampliamente distribuida en México, en América se distribuye desde Estados Unidos hasta Sudamérica (Deloya 2000, 2003). Por su parte, O. mictlensis sólo era conocida de Tepeyehualco, Puebla que es la localidad tipo (Deloya 1995, 2000), por lo que Zapotitlán de las Salinas es una nueva localidad de distribución, Deloya (2000) señala que posiblemente la especie sea endémica del área xérica del valle de Tehuacán.
Finalmente, en este tipo de ambientes como en muchos otros, los insectos constituyen una importante fuente de alimento para varios grupos de vertebrados, por ejemplo las aves y murciélagos que para el valle de Tehuacán han sido registradas 90 y 34 especies respectivamente, de las cuales casi el 50% de las especies son insectívoras (Arizamendi & Espinosa de los Monteros 1996, Rojas-Martínez & Valiente-Banuet 1996). Por lo tanto, es fundamental el mantenimiento del mezquital y el matorral espinoso tanto el establecido en las terrazas aluviales como el localizado en las inmediaciones de los arroyos, debido a que corresponde con los sitios más importantes para el desarrollo de las poblaciones de escarabajos necrófilos y posiblemente de otros grupos de artrópodos de los cuales hacen falta estudios. Sin embargo, estos lugares están sujetos a un proceso de erosión natural y tienen una fuerte presión de la población humana por corresponder a las áreas donde se desarrollan las actividades productivas de la región.
Agradecimientos
Los autores agradecen por la corroboración taxonómica de las especies a Cuauhtémoc Deloya y a José Luis Navarrete-Heredia. Este estudio fue financiado parcialmente por el Proyecto Zoología de la División de Investigación y Posgrado de la FES-Iztacala, UNAM.
Referencias
Acuña, J.A. 2004. Coleópteros necrófilos (Scarabaeidae, Silphidae, Staphylinidae e Histeridae) de la Sierra Norte de Puebla, México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Arellano, G.L. 1998. Distribución de Silphidae (Coleoptera: Insecta) en la región central del estado de Veracruz. Dugesiana 5: 1-16. [ Links ]
Arias, T.A.A., M.T. Valverde & J. Reyes S. 2000. Las plantas de la región de Zapotitlán de las Salinas, Puebla. INE, Red para el Desarrollo Sostenible, A. C. Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Arizmendi, M.C. & A. Espinosa de los Monteros. 1996. Avifauna de los bosques de cactáceas columnares del Valle de Tehuacán, Puebla. Acta Zool. Mex. (n.s.) 67: 25-46. [ Links ]
Beatley, J.C. 1974. Phenological Events and Their Environmental Triggers in Mojave Desert Ecosystems. Ecology 55: 856-863. [ Links ]
Caballero, U. 2003. Staphylinidae necrófilos (Insecta: Coleoptera) de la Sierra de Huautla, Morelos. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Caballero, U., J.L. León-Cortés & A. Morón-Ríos. 2009. Response of rove beetles (Staphylinidae) to various habitat types and change in Southern Mexico. J. Anim. Ecol. 264: 67-75. [ Links ]
Camargo-Ricalde, S.L., S.S. Dhillion & R. Grether. 2002. Community structure of endemic Mimosa species and environmental heterogeneity in a semi-arid Mexican valley. J. Veg. Sci. 13: 697-704. [ Links ]
Cejudo-Espinosa, E. & C. Deloya. 2005. Coleoptera necrófilos del bosque de Pinus hartwegii del Nevado de Toluca, México. Folia Entomol. Mex. 44: 67-73. [ Links ]
Cerecero, M.A. 2002. Contribución al conocimiento de los Scarabaeidae (Insecta: Coleoptera) necrófilos de la región de Los Tuxtlas Veracruz, México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Colwell, R.K. 2006. Estimates: Statistical estimation of species richness and shared species from samples. Version 8. (Consultado: 1 marzo 2012, purl.oclc.org/ estimates). [ Links ]
Dávila, P., J.L. Villaseñor, L. Medina, A. Ramírez, A. Salinas, J. Sánchez-Ken & P. Tenorio. 1993. Flora del Valle Tehuacán-Cuicatlán. Listados Florísticos de México X. México: Instituto de Biología, Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Deloya, C. 1992. Necrophilous Scarabaeidae and Trogidae beetles of tropical deciduous forest in Tepexco, Puebla, México. Acta Zool. Mex. (n.s.) 52: 1-13. [ Links ]
Deloya, C. 1995. A new species of Omorgus (Omorgus) Erichson from México (Coleoptera: Trogidae). Coleopt. Bull. 49: 153-156. [ Links ]
Deloya, C. 2000. Distribución de la familia Trogidae en México (Coleoptera: Lamellicornia). Acta Zool. Mex. (n.s.) 81: 63-76. [ Links ]
Deloya, C. 2003. Capítulo 6. Familia Trogidae, p.125-133. In M.A. Morón (ed.). Atlas de los escarabajos de México. Coleoptera: Lamellicornia, Vol. II Familias Scarabaeidae, Trogidae, Passalidae y Lucanidae. Argania, Barcelona, España. [ Links ]
Deloya, C. & M.A. Morón. 1998. Scarabaeoidea (Insecta: Coleoptera) necrófagos de “Los Tuxtlas”, Veracruz y Puerto Ángel, Oaxaca, México. Dugesiana 5: 17-28. [ Links ]
Deloya, C., G. Ruíz-Lizárraga & M.A. Morón. 1987. Análisis de la entomofauna necrófila de la Región de Jojutla, Morelos, México. Folia Entomol. Mex. 73: 157-171. [ Links ]
Estrada, A., R. Coates-Estrada, A. Anzures & P. Cammarano. 1998. Dung and Carrion Beetles in Tropical Rain Forest Fragments and Agricultural Habitats at Los Tuxtlas, Mexico. J. Trop. Ecol. 14: 577-593. [ Links ]
Fowler, N. 1986. The role of competition in plant communities in arid and semiarid regions. Ann. Rev. Ecol. Syst. 17: 89-110. [ Links ]
García, E. 1988. Modificaciones al sistema de clasificación climática de Koppen: para adaptarlo a las condiciones de la República Mexicana. Editado por el Autor, México. [ Links ]
Giller, P.S. & B.M. Doube. 1989. Experimental analysis of inter and intraspecific competition in dung beetle communities. J. Anim. Ecol. 58: 129-142. [ Links ]
Gómez-Jaimes, G., E. Jiménez-Sánchez & J. Padilla-Ramírez. 2004. Macrocoleópteros necrófilos (Sacarabaeidae, Trogidae y Silphidae) de la Reserva de la Biosfera Sierra de Huautla, Morelos, México, p. 273-277. In A. Morales, M. Ibarra, A.P. Rivera & S. Stanford (eds.). Entomología Mexicana, Vol. 3. Sociedad Mexicana de Entomología, Estado de México, México. [ Links ]
Hadley, N.F & S.R. Szarek. 1981. Productivity of desert ecosystems. Bioscience 31: 747-753. [ Links ]
Hammer, Ø., D.A.T. Harper & P.D. Ryan. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Paleo. Electr. 4: 10-50. [ Links ]
Hanski, I. 1983. Distributional ecology and abundance of dung and carrion-feeding beetles (Scarabaeidae) in tropical rain forests in Sarawak, Borneo. Acta Zool. Fenn. 167: 1-45. [ Links ]
Hanski, I. 1987. Nutritional Ecology of Dung and Carrion Feeding Insects, p. 836-884. In F. Slansky & J.G. Rodríguez (eds.). Off prints from nutritional ecology of insects, mites and spider. Wiley, Nueva York, EE.UU. [ Links ]
Halvorson, W.L. & T.P. Duncan. 1975. Productivity and Flowering of Winter Ephemerals in Relation to Sonoran Desert Shrubs. Am. Mid. Nat. 93: 311-319. [ Links ]
Hortal-Muñoz, J., F. Martín-Piera & M. Lobo. 2000. Dung Beetle Geographic Diversity Variation Along a Western Iberian Latitudinal Transect (Coleoptera: Scarabaeidae). Ann. Entomol. Soc. Am. 93: 235-243. [ Links ]
Huacuja, Z.A.H. 1982. Análisis de la fauna de coleópteros Staphyinidae saprófilos de Zacualtipan, Hidalgo. Tesis de licenciatura. Facultad de Ciencias, Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Jiménez-Sánchez, E. & J. Padilla-Ramírez. 1999. Estudio preliminar de Staphylinidae (Insecta: Coleoptera) de una región árida en Zapotitlán de las Salinas, Puebla, México, p. 107-111. In Memorias del XXXIV Con-greso Nacional de Entomología, Sociedad Mexicana de Entomología, Aguascalientes, Aguascalientes, México. [ Links ]
Jiménez-Sánchez, E., J.L. Navarrete-Heredia & J.R. Padilla-Ramírez. 2000. Estafilínidos (Coleoptera: Staphylinidae) necrófilos de la Sierra de Nanchititla, Estado de México, México. Folia Entomol. Mex. 108: 53-78. [ Links ]
Jiménez-Sánchez, E., J.R. Padilla-Ramírez, S. Stanford-Camargo & R. Quezada-García. 2001. Staphylinidae (Insecta: Coleoptera) necrófilos de “El Salto de las Granadas”, Guerrero, México, p. 55-68. In J.L. Naverrete-Heredia, H.E. Fierros- López & A. Burgos-Solorio (eds.). Tópicos sobre Coleoptera de México. Universidad de Guadalajara-Universidad Autónoma del estado de Morelos, Guadalajara, México. [ Links ]
Jiménez-Sánchez, E., O.M. Juárez-Gaytán & J.R. Padilla-Ramírez. 2011. Estafilínidos (Coleoptera : Staphyli-nidae ) necrófilos de Malinalco, Estado de México. Dugesiana 18: 73-84. [ Links ]
Jiménez-Sánchez, E., S. Zaragoza-Caballero & F.A. Noguera. 2009. Variación temporal de la diversidad de estafilínidos (Coleoptera: Staphylinidae) nocturnos en un bosque tropical caducifolio de México. Rev. Mex. Biodiv. 80: 157-168. [ Links ]
Kremen, C., R.K. Colwell, T.L. Erwin, D.D. Murphy, R.F. Noss & M.A. Sanjayan. 1993. Terrestrial Arthropod Assemblages: Their Use in Conservation Planning. Conserv. Biol. 7: 796-808. [ Links ]
Lobo, J.M., J. Hortal & F.J. Cabrero-Sañudo. 2006. Regional and local influence of grazing activity on the diversity of a semi-arid dung beetle community. Diversity Distrib. 12: 111-123. [ Links ]
Magurran, A.E. 1989. Diversidad ecológica y su medición. Vedra, Barcelona, España. [ Links ]
Márquez, J. 2001. Especies necrófilas de Staphylinidae (Insecta: Coleoptera) del Municipio de Tlayacapán, Morelos, México. Folia Entomol. Mex. 40: 93-131. [ Links ]
Márquez, J., J. Asiain & Q.J. Santiago-Jiménez. 2003. Especies de Staphylinidae (Coleoptera: Staphylinidae) de “El Mirador” Veracruz, México. Dugesiana 10: 21-46. [ Links ]
Márquez, J. 2003. Ecological patterns in necrophilous Staphylinidae (Insecta: Coleoptera) from Tlayacapan, Morelos, México. Acta Zool. Mex. (n.s.) 89: 69-83. [ Links ]
McAuliffe, J.R. 1986. Hervibore limited establishment of a Sonorian desert tree, Cercidium microphyllum. Ecology 67: 276-280. [ Links ]
Méndez-Castellanos, R. 2002. Macro-coleópteros necrófilos (Silphidae, Trogidae, Geotrupidae y Scarabaeidae) de la Sierra de Nanchititla, Estado de México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Méndez, I., J.L. Villaseñor, R. Lira, J.J. Morrone, P. Dávila & E. Ortiz. 2005. Toward the identification of a core zone in the Tehuacán-Cuicatlán Biosphere Reserve, Mexico, Based on Parsimony analysis of endemicity of flowering plant species. Interciencia 30: 267-274. [ Links ]
Montaña, C. & A. Valiente-Banuet. 1998. Floristic and Life-Form Diversity along an Altitudinal Gradient in an Intertropical Semiarid Mexican Region. Southwest Nat. 43: 25-39. [ Links ]
Mora-Aguilar, E.F. & E. Montes de Oca. 2009. Escarabajos necrófagos (Coleoptera : Scarabaeidae y Trogidae) de la región central baja de Veracruz, México. Acta Zool. Mex. (n.s.) 25: 569-588. [ Links ]
Morales, A., S. Cházaro & J.R. Padilla-Ramírez. 1998. Análisis de la comunidad de Coleoptera necrófilos de “Las Escolleras”, Alvarado, Veracruz, México. Dugesiana 5: 23-40. [ Links ]
Morón, M.A. & J.A. López-Méndez. 1985. Análisis de la entomofauna necrófila de un cafetal en el Soconusco, Chiapas, México. Folia Entomol. Mex. 63: 47-59. [ Links ]
Morón, M.A. & R.A. Terrón. 1984. Distribución altitudinal y estacional de los insectos necrófilos en la Sierra Norte de Hidalgo, México. Acta Zool. Mex. (n.s.) 3: 1-47. [ Links ]
Morón, M.A., J.F. Camal & O. Canul. 1986. Análisis de la entomofauna necrófila del área norte de la Reserva de la Biosfera “Sian Ka’an”, Quintana Roo, México. Folia Entomol. Mex. 69: 83-98. [ Links ]
Morón, M.A. 1987. The necrophagous Scarabaeinae beteles (Coleoptera: Scarabaeidae) from a coffee plantation in Chiapas, México: habits and phenology. Coleopt. Bull. 41: 225-232. [ Links ]
Muller, C.H. 1953. The Association of Desert Annuals with Shrubs. Am. J. Bot. 40: 53-60. [ Links ]
Navarrete-Heredia, J.L. & H.E. Fierros-López. 2000. 20. Silphidae (Coleoptera), p. 401-412. In B.J. Llorente, E. González & N. Papavero (eds.). Biodiversidad, taxonomía y biogeografía de artrópodos de México. Hacia una síntesis de su conocimiento. Vol. II. CONABIO-UNAM-Bayer, D.F., México. [ Links ]
Navarrete-Heredia, J.L. 1996. Coleópteros micetócolos de Basidiomycetes de San José de Los Laureles, Morelos, México. Tesis de Maestría, Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Navarrete-Heredia, J.L. 2009. Silphidae (Coleoptera) de México: Diversidad y distribución. Universidad de Guadalajara, Guadalajara, Jalisco, México. [ Links ]
Navarrete-Heredia, J.L., A.F. Newton, M.K. Thayer, J.S. Ashe & D.S. Chandler. 2002. Guía Ilustrada para los géneros de Staphylinidae (Coleoptera) de México. Illustrated guide to the genera of Staphylinidae (Coleoptera) of México. Universidad de Guadalajara-CONABIO, D.F., México. [ Links ]
Navarrete-Heredia, J.L. & G.A. Quiroz-Rocha. 2000. Macro-coleópteros necrófilos de San José de los Laureles, Morelos, México (Coleoptera: Scarabaeidae y Silphidae). Folia Entomol. Mex. 110: 1-13. [ Links ]
Numa, C., J.M. Lobo & J.R. Verdú. 2012. Scaling local abundance determinants in Mediterranean dung beetles. Insect Conserv. Diver. 5: 106-117. [ Links ]
Newton, A.F., M.K. Thayer, J.S. Ashe & D.S. Chandler. 2001. 22. Staphylinidae Latreille, 1802, p. 272-418. In R.H. Arnett, Jr. & M.C. Thomas (eds.). American beteles. Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. Vol. 1 CRS Press LLC, Boca Raton, Florida, EE.UU. [ Links ]
Oliveros, O. 2000. Descripción estructural de las comunidades vegetales en las terrazas fluviales del río El Salado, en el Valle de Zapotitlán de las Salinas, Puebla, México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Quiroz-Rocha, G.A., J.L. Navarrete-Heredia & P.A. Mar-tínez. 2008. Especies de Scarabaeinae (Coleoptera: Sacarabaeide) y Silphidae (Coeloptera) necrófilas de bosque de pino-encino y bosque mesófilo de montaña en el municipio de Mascota, Jalisco, México. Dugesiana 15: 27-37. [ Links ]
Reyes, G. 2001. Los coleópteros saprófagos (Scarabaeidae, Silphidae y Trogidae) del Salto de las Granadas Guerreo, México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Rivera-Cervantes, L.E. & E. García-Real. 1998. Análisis preliminar sobre la composición de los escarabajos necrófilos (Coleoptera: Silphidae y Scarabaeidae) presentes en dos bosques de pino (uno dañado por fuego), en la estación científica Las Joyas, Sierra de Manantlan, Jalisco, México. Dugesiana 5: 11-22. [ Links ]
Rivera-Cervantes, L.E. & G. Halffter. 1999. Monografía de las especies mexicanas de Canthon del subgénero Glaphyrocanthon (Coleoptera: Scarabaeidae: Scarabaeinae). Acta Zool. Mex. (n.s.) 77: 23-150. [ Links ]
Rojas-Martínez, A.E. & A. Valiente-Banuet. 1996. Análisis comparativo de la quiropterofauna del Valle de Tehuacan-Cuicatlán, Puebla-Oaxaca. Acta Zool. Mex. (n.s.) 67: 1-23. [ Links ]
Ruíz-Lizárraga, G. 1993. Contribución al conocimiento de los Staphylinidae (Coleoptera) necrófilos de Acahuizotla, Guerrero. Tesis de licenciatura, Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa, D.F. México. [ Links ]
SAS Institute Inc. 1998. SAS user’s guide; statistics 1998. SAS Institute Cary North Carolina. EE.UU. [ Links ]
Servicio Meteorológico Nacional (SMN). 2012. Climatología. Normales climatológicas por estación. Comisión Nacional del Agua, México. (Consultado: 29 febrero 2012, http://smn.cna.gob.mx/index.php?option=com_content&view=article&id=42&Ite mid=28). [ Links ]
Sánchez-Ramos, G., J. Lobo, M. Lara & P. Reyes-Castillo. 1993. Distribución altitudinal y estacional de la entomofauna necrófila en la Reserva de la Biosfera “El Cielo”, Tamaulipas, México. BIOTAM 5: 13-24. [ Links ]
Smetana, A. 1995. Rove Beetles of the subtribe Philonthina of America North of Mexico (Coleoptera: Staphylinidae) classification, phylogeny and taxonomic revision. Memoirs on Entomology, International. Vol. 3 Associate publishers, Gainesville, Florida, EE.UU. [ Links ]
Terrón, R.A., S. Anduaga & M.A. Morón. 1991. Análisis de la coleopterofauna necrófila de la Reserva de la Biosfera «La Michilia», Durango, México. Folia Entomol. Mex. 81: 315-324. [ Links ]
Trevilla-Rebollar, A., C. Deloya & J.R. Padilla-Ramírez. 2010. Coleópteros necrófilos (Scarabaeidae, Silphidae y Trogidae) de Malinalco, Estado de México, México. Neotrop. Entomol. 39: 486-495. [ Links ]
Valiente-Banuet, A., A. Bolongaro-Crevenna, O. Briones, E. Ezcurra, M. Rosas, H. Nuñez, G. Barnard & E. Vázquez. 1991. Spatial relationships between cacti and nurse shrubs in a semi-arid environment in central Mexico. J. Veg. Sci. 2: 15-20. [ Links ]
Valiente-Banuet, A., A. Casas, A. Alcántara, P. Dávila, N. Flores-Hernández, M.C. Arizmendi, J.L. Villaseñor & J. Ortega R. 2000. La vegetación del valle de Tehuacan-Cuicatlán. Bol. Soc. Bot. México 67: 24-74. [ Links ]
Valiente-Banuet, A., M.C., Arizmendi, A. Rojas-Martínez & L. Domínguez-Canseco. 1996 Ecological relationships between columnar cacti and nectar-feeding bats in Mexico. J. Trop. Ecol. 12: 103-11. [ Links ]
Villaseñor, J.L., P. Dávila & F. Chiang. 1990. Fitogeografía del valle de Tehuacán-Cuicatlán. Bol. Soc. Bot. México 50: 135-149. [ Links ]
Whitford, W.G. 1978. Structure and seasonal activity of Chihuahua desert ant communities. Insectes Soc. 25: 79-88. [ Links ]
Wolda, H. 1978. Fluctuations in abundance of tropical insects. Am. Nat. 112: 1017-1045. [ Links ]
Zar, J.H. 1984. Biostatiscal Analysis. Prentice Hall, Eaglewood, EE.UU. [ Links ]
Zaragoza-Caballero, S., F.A. Noguera, J.A. Chemsak, E. González-Soriano, A. Rodríguez-Palafox, E. Ramírez-García & R. Ayala. 2003. Diversity of Lycidae, Phengodidae, Lampyridae, and Cantharidae (Coleoptera) in a tropical dry forest region in México: Sierra de Huautla, Morelos. Pan-Pac. Entomol. 7: 3-37. [ Links ]
Arellano, G.L. 1998. Distribución de Silphidae (Coleoptera: Insecta) en la región central del estado de Veracruz. Dugesiana 5: 1-16. [ Links ]
Arias, T.A.A., M.T. Valverde & J. Reyes S. 2000. Las plantas de la región de Zapotitlán de las Salinas, Puebla. INE, Red para el Desarrollo Sostenible, A. C. Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Arizmendi, M.C. & A. Espinosa de los Monteros. 1996. Avifauna de los bosques de cactáceas columnares del Valle de Tehuacán, Puebla. Acta Zool. Mex. (n.s.) 67: 25-46. [ Links ]
Beatley, J.C. 1974. Phenological Events and Their Environmental Triggers in Mojave Desert Ecosystems. Ecology 55: 856-863. [ Links ]
Caballero, U. 2003. Staphylinidae necrófilos (Insecta: Coleoptera) de la Sierra de Huautla, Morelos. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Caballero, U., J.L. León-Cortés & A. Morón-Ríos. 2009. Response of rove beetles (Staphylinidae) to various habitat types and change in Southern Mexico. J. Anim. Ecol. 264: 67-75. [ Links ]
Camargo-Ricalde, S.L., S.S. Dhillion & R. Grether. 2002. Community structure of endemic Mimosa species and environmental heterogeneity in a semi-arid Mexican valley. J. Veg. Sci. 13: 697-704. [ Links ]
Cejudo-Espinosa, E. & C. Deloya. 2005. Coleoptera necrófilos del bosque de Pinus hartwegii del Nevado de Toluca, México. Folia Entomol. Mex. 44: 67-73. [ Links ]
Cerecero, M.A. 2002. Contribución al conocimiento de los Scarabaeidae (Insecta: Coleoptera) necrófilos de la región de Los Tuxtlas Veracruz, México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Colwell, R.K. 2006. Estimates: Statistical estimation of species richness and shared species from samples. Version 8. (Consultado: 1 marzo 2012, purl.oclc.org/ estimates). [ Links ]
Dávila, P., J.L. Villaseñor, L. Medina, A. Ramírez, A. Salinas, J. Sánchez-Ken & P. Tenorio. 1993. Flora del Valle Tehuacán-Cuicatlán. Listados Florísticos de México X. México: Instituto de Biología, Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Deloya, C. 1992. Necrophilous Scarabaeidae and Trogidae beetles of tropical deciduous forest in Tepexco, Puebla, México. Acta Zool. Mex. (n.s.) 52: 1-13. [ Links ]
Deloya, C. 1995. A new species of Omorgus (Omorgus) Erichson from México (Coleoptera: Trogidae). Coleopt. Bull. 49: 153-156. [ Links ]
Deloya, C. 2000. Distribución de la familia Trogidae en México (Coleoptera: Lamellicornia). Acta Zool. Mex. (n.s.) 81: 63-76. [ Links ]
Deloya, C. 2003. Capítulo 6. Familia Trogidae, p.125-133. In M.A. Morón (ed.). Atlas de los escarabajos de México. Coleoptera: Lamellicornia, Vol. II Familias Scarabaeidae, Trogidae, Passalidae y Lucanidae. Argania, Barcelona, España. [ Links ]
Deloya, C. & M.A. Morón. 1998. Scarabaeoidea (Insecta: Coleoptera) necrófagos de “Los Tuxtlas”, Veracruz y Puerto Ángel, Oaxaca, México. Dugesiana 5: 17-28. [ Links ]
Deloya, C., G. Ruíz-Lizárraga & M.A. Morón. 1987. Análisis de la entomofauna necrófila de la Región de Jojutla, Morelos, México. Folia Entomol. Mex. 73: 157-171. [ Links ]
Estrada, A., R. Coates-Estrada, A. Anzures & P. Cammarano. 1998. Dung and Carrion Beetles in Tropical Rain Forest Fragments and Agricultural Habitats at Los Tuxtlas, Mexico. J. Trop. Ecol. 14: 577-593. [ Links ]
Fowler, N. 1986. The role of competition in plant communities in arid and semiarid regions. Ann. Rev. Ecol. Syst. 17: 89-110. [ Links ]
García, E. 1988. Modificaciones al sistema de clasificación climática de Koppen: para adaptarlo a las condiciones de la República Mexicana. Editado por el Autor, México. [ Links ]
Giller, P.S. & B.M. Doube. 1989. Experimental analysis of inter and intraspecific competition in dung beetle communities. J. Anim. Ecol. 58: 129-142. [ Links ]
Gómez-Jaimes, G., E. Jiménez-Sánchez & J. Padilla-Ramírez. 2004. Macrocoleópteros necrófilos (Sacarabaeidae, Trogidae y Silphidae) de la Reserva de la Biosfera Sierra de Huautla, Morelos, México, p. 273-277. In A. Morales, M. Ibarra, A.P. Rivera & S. Stanford (eds.). Entomología Mexicana, Vol. 3. Sociedad Mexicana de Entomología, Estado de México, México. [ Links ]
Hadley, N.F & S.R. Szarek. 1981. Productivity of desert ecosystems. Bioscience 31: 747-753. [ Links ]
Hammer, Ø., D.A.T. Harper & P.D. Ryan. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Paleo. Electr. 4: 10-50. [ Links ]
Hanski, I. 1983. Distributional ecology and abundance of dung and carrion-feeding beetles (Scarabaeidae) in tropical rain forests in Sarawak, Borneo. Acta Zool. Fenn. 167: 1-45. [ Links ]
Hanski, I. 1987. Nutritional Ecology of Dung and Carrion Feeding Insects, p. 836-884. In F. Slansky & J.G. Rodríguez (eds.). Off prints from nutritional ecology of insects, mites and spider. Wiley, Nueva York, EE.UU. [ Links ]
Halvorson, W.L. & T.P. Duncan. 1975. Productivity and Flowering of Winter Ephemerals in Relation to Sonoran Desert Shrubs. Am. Mid. Nat. 93: 311-319. [ Links ]
Hortal-Muñoz, J., F. Martín-Piera & M. Lobo. 2000. Dung Beetle Geographic Diversity Variation Along a Western Iberian Latitudinal Transect (Coleoptera: Scarabaeidae). Ann. Entomol. Soc. Am. 93: 235-243. [ Links ]
Huacuja, Z.A.H. 1982. Análisis de la fauna de coleópteros Staphyinidae saprófilos de Zacualtipan, Hidalgo. Tesis de licenciatura. Facultad de Ciencias, Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Jiménez-Sánchez, E. & J. Padilla-Ramírez. 1999. Estudio preliminar de Staphylinidae (Insecta: Coleoptera) de una región árida en Zapotitlán de las Salinas, Puebla, México, p. 107-111. In Memorias del XXXIV Con-greso Nacional de Entomología, Sociedad Mexicana de Entomología, Aguascalientes, Aguascalientes, México. [ Links ]
Jiménez-Sánchez, E., J.L. Navarrete-Heredia & J.R. Padilla-Ramírez. 2000. Estafilínidos (Coleoptera: Staphylinidae) necrófilos de la Sierra de Nanchititla, Estado de México, México. Folia Entomol. Mex. 108: 53-78. [ Links ]
Jiménez-Sánchez, E., J.R. Padilla-Ramírez, S. Stanford-Camargo & R. Quezada-García. 2001. Staphylinidae (Insecta: Coleoptera) necrófilos de “El Salto de las Granadas”, Guerrero, México, p. 55-68. In J.L. Naverrete-Heredia, H.E. Fierros- López & A. Burgos-Solorio (eds.). Tópicos sobre Coleoptera de México. Universidad de Guadalajara-Universidad Autónoma del estado de Morelos, Guadalajara, México. [ Links ]
Jiménez-Sánchez, E., O.M. Juárez-Gaytán & J.R. Padilla-Ramírez. 2011. Estafilínidos (Coleoptera : Staphyli-nidae ) necrófilos de Malinalco, Estado de México. Dugesiana 18: 73-84. [ Links ]
Jiménez-Sánchez, E., S. Zaragoza-Caballero & F.A. Noguera. 2009. Variación temporal de la diversidad de estafilínidos (Coleoptera: Staphylinidae) nocturnos en un bosque tropical caducifolio de México. Rev. Mex. Biodiv. 80: 157-168. [ Links ]
Kremen, C., R.K. Colwell, T.L. Erwin, D.D. Murphy, R.F. Noss & M.A. Sanjayan. 1993. Terrestrial Arthropod Assemblages: Their Use in Conservation Planning. Conserv. Biol. 7: 796-808. [ Links ]
Lobo, J.M., J. Hortal & F.J. Cabrero-Sañudo. 2006. Regional and local influence of grazing activity on the diversity of a semi-arid dung beetle community. Diversity Distrib. 12: 111-123. [ Links ]
Magurran, A.E. 1989. Diversidad ecológica y su medición. Vedra, Barcelona, España. [ Links ]
Márquez, J. 2001. Especies necrófilas de Staphylinidae (Insecta: Coleoptera) del Municipio de Tlayacapán, Morelos, México. Folia Entomol. Mex. 40: 93-131. [ Links ]
Márquez, J., J. Asiain & Q.J. Santiago-Jiménez. 2003. Especies de Staphylinidae (Coleoptera: Staphylinidae) de “El Mirador” Veracruz, México. Dugesiana 10: 21-46. [ Links ]
Márquez, J. 2003. Ecological patterns in necrophilous Staphylinidae (Insecta: Coleoptera) from Tlayacapan, Morelos, México. Acta Zool. Mex. (n.s.) 89: 69-83. [ Links ]
McAuliffe, J.R. 1986. Hervibore limited establishment of a Sonorian desert tree, Cercidium microphyllum. Ecology 67: 276-280. [ Links ]
Méndez-Castellanos, R. 2002. Macro-coleópteros necrófilos (Silphidae, Trogidae, Geotrupidae y Scarabaeidae) de la Sierra de Nanchititla, Estado de México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Méndez, I., J.L. Villaseñor, R. Lira, J.J. Morrone, P. Dávila & E. Ortiz. 2005. Toward the identification of a core zone in the Tehuacán-Cuicatlán Biosphere Reserve, Mexico, Based on Parsimony analysis of endemicity of flowering plant species. Interciencia 30: 267-274. [ Links ]
Montaña, C. & A. Valiente-Banuet. 1998. Floristic and Life-Form Diversity along an Altitudinal Gradient in an Intertropical Semiarid Mexican Region. Southwest Nat. 43: 25-39. [ Links ]
Mora-Aguilar, E.F. & E. Montes de Oca. 2009. Escarabajos necrófagos (Coleoptera : Scarabaeidae y Trogidae) de la región central baja de Veracruz, México. Acta Zool. Mex. (n.s.) 25: 569-588. [ Links ]
Morales, A., S. Cházaro & J.R. Padilla-Ramírez. 1998. Análisis de la comunidad de Coleoptera necrófilos de “Las Escolleras”, Alvarado, Veracruz, México. Dugesiana 5: 23-40. [ Links ]
Morón, M.A. & J.A. López-Méndez. 1985. Análisis de la entomofauna necrófila de un cafetal en el Soconusco, Chiapas, México. Folia Entomol. Mex. 63: 47-59. [ Links ]
Morón, M.A. & R.A. Terrón. 1984. Distribución altitudinal y estacional de los insectos necrófilos en la Sierra Norte de Hidalgo, México. Acta Zool. Mex. (n.s.) 3: 1-47. [ Links ]
Morón, M.A., J.F. Camal & O. Canul. 1986. Análisis de la entomofauna necrófila del área norte de la Reserva de la Biosfera “Sian Ka’an”, Quintana Roo, México. Folia Entomol. Mex. 69: 83-98. [ Links ]
Morón, M.A. 1987. The necrophagous Scarabaeinae beteles (Coleoptera: Scarabaeidae) from a coffee plantation in Chiapas, México: habits and phenology. Coleopt. Bull. 41: 225-232. [ Links ]
Muller, C.H. 1953. The Association of Desert Annuals with Shrubs. Am. J. Bot. 40: 53-60. [ Links ]
Navarrete-Heredia, J.L. & H.E. Fierros-López. 2000. 20. Silphidae (Coleoptera), p. 401-412. In B.J. Llorente, E. González & N. Papavero (eds.). Biodiversidad, taxonomía y biogeografía de artrópodos de México. Hacia una síntesis de su conocimiento. Vol. II. CONABIO-UNAM-Bayer, D.F., México. [ Links ]
Navarrete-Heredia, J.L. 1996. Coleópteros micetócolos de Basidiomycetes de San José de Los Laureles, Morelos, México. Tesis de Maestría, Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Navarrete-Heredia, J.L. 2009. Silphidae (Coleoptera) de México: Diversidad y distribución. Universidad de Guadalajara, Guadalajara, Jalisco, México. [ Links ]
Navarrete-Heredia, J.L., A.F. Newton, M.K. Thayer, J.S. Ashe & D.S. Chandler. 2002. Guía Ilustrada para los géneros de Staphylinidae (Coleoptera) de México. Illustrated guide to the genera of Staphylinidae (Coleoptera) of México. Universidad de Guadalajara-CONABIO, D.F., México. [ Links ]
Navarrete-Heredia, J.L. & G.A. Quiroz-Rocha. 2000. Macro-coleópteros necrófilos de San José de los Laureles, Morelos, México (Coleoptera: Scarabaeidae y Silphidae). Folia Entomol. Mex. 110: 1-13. [ Links ]
Numa, C., J.M. Lobo & J.R. Verdú. 2012. Scaling local abundance determinants in Mediterranean dung beetles. Insect Conserv. Diver. 5: 106-117. [ Links ]
Newton, A.F., M.K. Thayer, J.S. Ashe & D.S. Chandler. 2001. 22. Staphylinidae Latreille, 1802, p. 272-418. In R.H. Arnett, Jr. & M.C. Thomas (eds.). American beteles. Archostemata, Myxophaga, Adephaga, Polyphaga: Staphyliniformia. Vol. 1 CRS Press LLC, Boca Raton, Florida, EE.UU. [ Links ]
Oliveros, O. 2000. Descripción estructural de las comunidades vegetales en las terrazas fluviales del río El Salado, en el Valle de Zapotitlán de las Salinas, Puebla, México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Quiroz-Rocha, G.A., J.L. Navarrete-Heredia & P.A. Mar-tínez. 2008. Especies de Scarabaeinae (Coleoptera: Sacarabaeide) y Silphidae (Coeloptera) necrófilas de bosque de pino-encino y bosque mesófilo de montaña en el municipio de Mascota, Jalisco, México. Dugesiana 15: 27-37. [ Links ]
Reyes, G. 2001. Los coleópteros saprófagos (Scarabaeidae, Silphidae y Trogidae) del Salto de las Granadas Guerreo, México. Tesis de licenciatura, Universidad Nacional Autónoma de México, Estado de México, México. [ Links ]
Rivera-Cervantes, L.E. & E. García-Real. 1998. Análisis preliminar sobre la composición de los escarabajos necrófilos (Coleoptera: Silphidae y Scarabaeidae) presentes en dos bosques de pino (uno dañado por fuego), en la estación científica Las Joyas, Sierra de Manantlan, Jalisco, México. Dugesiana 5: 11-22. [ Links ]
Rivera-Cervantes, L.E. & G. Halffter. 1999. Monografía de las especies mexicanas de Canthon del subgénero Glaphyrocanthon (Coleoptera: Scarabaeidae: Scarabaeinae). Acta Zool. Mex. (n.s.) 77: 23-150. [ Links ]
Rojas-Martínez, A.E. & A. Valiente-Banuet. 1996. Análisis comparativo de la quiropterofauna del Valle de Tehuacan-Cuicatlán, Puebla-Oaxaca. Acta Zool. Mex. (n.s.) 67: 1-23. [ Links ]
Ruíz-Lizárraga, G. 1993. Contribución al conocimiento de los Staphylinidae (Coleoptera) necrófilos de Acahuizotla, Guerrero. Tesis de licenciatura, Universidad Nacional Autónoma de México, D.F., México. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa, D.F. México. [ Links ]
SAS Institute Inc. 1998. SAS user’s guide; statistics 1998. SAS Institute Cary North Carolina. EE.UU. [ Links ]
Servicio Meteorológico Nacional (SMN). 2012. Climatología. Normales climatológicas por estación. Comisión Nacional del Agua, México. (Consultado: 29 febrero 2012, http://smn.cna.gob.mx/index.php?option=com_content&view=article&id=42&Ite mid=28). [ Links ]
Sánchez-Ramos, G., J. Lobo, M. Lara & P. Reyes-Castillo. 1993. Distribución altitudinal y estacional de la entomofauna necrófila en la Reserva de la Biosfera “El Cielo”, Tamaulipas, México. BIOTAM 5: 13-24. [ Links ]
Smetana, A. 1995. Rove Beetles of the subtribe Philonthina of America North of Mexico (Coleoptera: Staphylinidae) classification, phylogeny and taxonomic revision. Memoirs on Entomology, International. Vol. 3 Associate publishers, Gainesville, Florida, EE.UU. [ Links ]
Terrón, R.A., S. Anduaga & M.A. Morón. 1991. Análisis de la coleopterofauna necrófila de la Reserva de la Biosfera «La Michilia», Durango, México. Folia Entomol. Mex. 81: 315-324. [ Links ]
Trevilla-Rebollar, A., C. Deloya & J.R. Padilla-Ramírez. 2010. Coleópteros necrófilos (Scarabaeidae, Silphidae y Trogidae) de Malinalco, Estado de México, México. Neotrop. Entomol. 39: 486-495. [ Links ]
Valiente-Banuet, A., A. Bolongaro-Crevenna, O. Briones, E. Ezcurra, M. Rosas, H. Nuñez, G. Barnard & E. Vázquez. 1991. Spatial relationships between cacti and nurse shrubs in a semi-arid environment in central Mexico. J. Veg. Sci. 2: 15-20. [ Links ]
Valiente-Banuet, A., A. Casas, A. Alcántara, P. Dávila, N. Flores-Hernández, M.C. Arizmendi, J.L. Villaseñor & J. Ortega R. 2000. La vegetación del valle de Tehuacan-Cuicatlán. Bol. Soc. Bot. México 67: 24-74. [ Links ]
Valiente-Banuet, A., M.C., Arizmendi, A. Rojas-Martínez & L. Domínguez-Canseco. 1996 Ecological relationships between columnar cacti and nectar-feeding bats in Mexico. J. Trop. Ecol. 12: 103-11. [ Links ]
Villaseñor, J.L., P. Dávila & F. Chiang. 1990. Fitogeografía del valle de Tehuacán-Cuicatlán. Bol. Soc. Bot. México 50: 135-149. [ Links ]
Whitford, W.G. 1978. Structure and seasonal activity of Chihuahua desert ant communities. Insectes Soc. 25: 79-88. [ Links ]
Wolda, H. 1978. Fluctuations in abundance of tropical insects. Am. Nat. 112: 1017-1045. [ Links ]
Zar, J.H. 1984. Biostatiscal Analysis. Prentice Hall, Eaglewood, EE.UU. [ Links ]
Zaragoza-Caballero, S., F.A. Noguera, J.A. Chemsak, E. González-Soriano, A. Rodríguez-Palafox, E. Ramírez-García & R. Ayala. 2003. Diversity of Lycidae, Phengodidae, Lampyridae, and Cantharidae (Coleoptera) in a tropical dry forest region in México: Sierra de Huautla, Morelos. Pan-Pac. Entomol. 7: 3-37. [ Links ]
*Correspondencia a:
Esteban Jiménez-Sánchez. Facultad de Estudios Superiores Iztacala, UNAM. Av. de los Barrios N° 1, CP. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de México, México; estjimsan@yahoo.com.mx
Roberto Quezada-García. Département des Sciences du Bois et de la Forêt, Faculté de Foresterie et de Géomatique, Université Laval, G1V 0A6, Québec, Canada; roberto_queza71@hotmail.com
Jorge Padilla-Ramírez. Facultad de Estudios Superiores Iztacala, UNAM. Av. de los Barrios N° 1, CP. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de México, México; jorgepr@servidor.unam.mx
1. Facultad de Estudios Superiores Iztacala, UNAM. Av. de los Barrios N° 1, CP. 54090, Los Reyes Iztacala, Tlalnepantla, Estado de México, México; estjimsan@yahoo.com.mx, jorgepr@servidor.unam.mx
2. Département des Sciences du Bois et de la Forêt, Faculté de Foresterie et de Géomatique, Université Laval, G1V 0A6, Québec, Canada; roberto_queza71@hotmail.com
Recibido 19-IV-2012. Corregido 09-X-2012. Aceptado 06-XI-2012.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}