











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Los bosques templados de México se caracterizan por su alta diversidad biológica y especies endémicas (1). Una de estas especies endémicas es Abies hickelii (Flous & Gaussen), que habita en las montañas centrales de Oaxaca, Mezcalapa y Copainalá (Chiapas), y en la región de Carabinas de Xico, Veracruz (2),(3). Actualmente, la especie está restringida a los estados de Veracruz, Oaxaca, Guerrero y Chiapas (4),(5),(3). Cabe destacar que A. hickelii está catalogada como especie en peligro de extinción en la Norma Oficial Mexicana (NOM-059-SEMARNAT-2010) y por la Unión Internacional para la Conservación de la Naturaleza (UICN) (6).
La comprensión de los mecanismos y factores que influyen en la distribución y abundancia de las especies ha sido una preocupación durante mucho tiempo para los ecólogos (7). Se han propuesto dos enfoques teóricos principales para explicar los patrones de distribución y abundancia de las especies: las interacciones entre individuos, incluida la competencia, y las características ambientales, asumiendo que cada especie responde de forma independiente a estos factores (8).
En el caso de A. hickelii (Flous & Gaussen), la disposición espacial de su población se rige por múltiples factores, como la disponibilidad de recursos, las interacciones con otros individuos y los factores ambientales (9),(10). Sin embargo, hace falta fortalecer los conocimientos sobre los factores geomorfológicos y la orientación espacial. Estos factores son cruciales para comprender cómo las características geográficas y la configuración del terreno pueden influir en la distribución y el comportamiento de las plantas a escala local.
Dada la presión sobre las especies en categorías de riesgo debido al cambio en el uso del suelo, el cambio climático y otras actividades humanas (3), estudiar el patrón de distribución y la densidad de las poblaciones en diferentes etapas de crecimiento es esencial para comprender la dinámica de una especie a una edad temprana y su respuesta a los factores geomorfológicos locales. El objetivo de este estudio fue determinar si la regeneración natural de Abies hickelii (Flous & Gaussen) está asociada a una dirección específica, utilizando el norte magnético como referencia. Además, se establecieron dos objetivos secundarios: identificar las especies más comunes que coexisten con A. hickelii en el área de estudio y describir la topografía local. Para ello, se dividió el contorno de los árboles de referencia en cuatro cuadrantes, y se comparó la proporción de individuos presentes en cada cuadrante, partiendo de la hipótesis de que la regeneración natural se distribuye uniformemente en todas las direcciones.
Materiales y métodos
Área de estudio. El estudio se realizó en el predio de Santo Domingo Xagacía, situado en las cadenas montañosas de la Sierra Norte de Oaxaca, México, entre los paralelos 17° 09' de latitud norte y los meridianos 96° 16' de longitud oeste (ver Figura 1). La población investigada abarca aproximadamente 112.27 hectáreas y presenta una topografía irregular, con pendientes que oscilan entre el 6% y el 50%. Las exposiciones varían en distancias cortas, siendo las más comunes las orientaciones Norte, Noreste, Este y Noroeste. La elevación sobre el nivel del mar fluctúa entre 2767 y 3095 metros.

Figure 1. Location of the study area.
Los tipos de clima presentes en el área de estudio son: templado semifrío húmedo, templado semicálido subhúmedo y templado húmedo, según la clasificación de Köppen, modificada por García (11). La temperatura media anual en el área de muestreo varía entre 5 y 18 °C, mientras que la precipitación media anual oscila entre 1301 y 1398 mm. Estas condiciones climáticas favorecen la presencia predominante de bosques de pino y vegetación secundaria arbórea de pino-encino. El suelo característico de la región es el Cambisol húmico, conocido por su riqueza en materia orgánica (12).
Muestreo y obtención de variables estudiadas. En la población estudiada se establecieron sistemáticamente 10 parcelas circulares de 56.42 m de radio con una superficie de 10,000 m2 cada una. Entre sus límites se establecieron a la vez, cuatro unidades de muestreo secundarias, conocidas como “sitios”, cada una con una superficie de 400 m2 de superficie, distribuidos en forma de “Y” invertida con orientación al Norte, emulando el diseño de muestreo de la Comisión Nacional Forestal (13), para el Inventario Nacional Forestal de México. La disposición de las parcelas principales se determinó considerando las condiciones locales, como la exposición, la pendiente, la cercanía de un arroyo, el tipo de vegetación y la densidad. Se excluyeron lugares con perturbaciones antropogénicas significativas, principalmente una zona de extracción reciente de árboles del género Pinus afectados por insectos descortezadores y defoliadores.
Definición del árbol central y del área de muestreo de regeneración natural En este estudio se denomina árbol central o árbol madre, a los individuos de A. hickelii que cumplieron con lo establecido por Ávila-Bello et al. 14). y García-García et al. (15). Para dicho propósito se tomaron en cuenta los siguientes criterios: (1) el individuo seleccionado debía estar entre los límites de la parcela de 56.42 m de radio; (2)contar con un diámetro normal ≥ a 7.5, asumiendo que A. hickelii inicia su etapa reproductiva cuando alcanza el diámetro normal de 7.5 cm (14), (16); (3) no debían presentar plagas ni enfermedades ni daños mecánicos o físicos; y (4) sus copas no debían traslaparse con las copas de otros individuos adultos de la misma especie.
Después de identificar los individuos que se seleccionaron como “árboles madres”, se definió el área de muestreo para cuantificar la regeneración natural alrededor de cada uno de ellos. Para definir esta área, se midió la distancia desde el extremo de la copa de cada árbol madre y se añadió un margen de 1.3 metros adicionales (Figura 2). Este valor de 1.3 metros se obtuvo calculando la desviación estándar del radio de la copa de los árboles adultos, lo que se hizo mediante un muestreo piloto en las parcelas de 400 metros cuadrados. Como resultado, el tamaño del área de muestreo para la regeneración varió de un árbol madre a otro.
Para identificar las especies vecinas, se recolectaron ejemplares botánicos y se enviaron al herbario del Instituto de Estudios Ambientales de la Universidad de la Sierra Juárez para su determinación taxonómica.
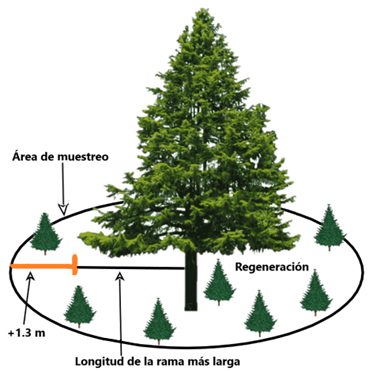
Figura 2 Esquema que muestra la distribución espacial de la regeneración natural alrededor de un árbol central o árbol madre.
Figure 2. Diagram showing the spatial distribution of natural regeneration around a central or mother tree.
Por otro lado, en las parcelas de 400 m2, se registraron tres variables topográficas: la pendiente (%), la exposición predominante de la ladera (%) y la elevación sobre el nivel del mar (m). La elevación sobre el nivel del mar se registró con un geoposicionador marca Garmin eTrex® 20x con sistema de posicionamiento global (GPS). La pendiente se midió en porcentaje utilizando un clinómetro de la marca Suunto®, y la exposición de la ladera se determinó con la brújula del geoposicionador GPS.
En esta investigación, se define como regeneración natural a los individuos de la especie estudiada con un diámetro normal inferior a 7.5 cm. Se realizaron mediciones de la altura total de estos individuos, desde la base del suelo hasta el ápice principal, utilizando una regla graduada o una cinta métrica Truper®, según la altura.
Para evaluar la distribución de la regeneración alrededor de los árboles de referencia, se segmentó el área circular en torno a cada árbol madre en cuatro cuadrantes, tomando como referencia el norte magnético. El cuadrante norte (N) abarca el sector entre 315° y 45° de azimut, con una desviación de ± 45°. El cuadrante este (E) cubre el intervalo de 45° a 135°, el cuadrante sur (S) de 135° a 225°, y el cuadrante oeste (O) de 225° a 315°. Una vez segmentado el área alrededor de cada árbol central, se registró la frecuencia absoluta de la regeneración natural en cada cuadrante y se calcularon las proporciones y los intervalos de confianza correspondientes.
Cálculo de la proporción y el intervalo de confianza de la regeneración natural. La proporción de la regeneración en cada cuadrante se determinó mediante la siguiente formula (38) eq 1.

donde: 𝑝 = es la proporción del i-ésimo cuadrante del árbol central; a = es el número de individuos en el i-ésimo cuadrante; n = es el número de individuos observados en los cuatro cuadrantes.
El intervalo de confianza para estas proporciones se calculó utilizando el método de Clopper-Pearson (17), usando el programa R, versión 4.3.0 (18). Además, se empleó la función de densidad de probabilidad de Weibull (19) para modelar la distribución de frecuencias de la exposición, la pendiente y la altitud sobre el nivel del mar, debido a la alta flexibilidad de esta función (20). Se asumió que la distribución de frecuencias de la regeneración natural es desconocida a priori (21).
El modelo de Weibull de dos parámetros se expresa de la siguiente manera (19),(22) eq 2 y 3:

y su función acumulada es:

Donde 𝑐>0, es el parámetro de forma, y 𝑏>0 el parámetro de escala.
Resultados
Las especies arbóreas más comunes en la población estudiada fueron Quercus laurina (Humb et Bonpl.), Pinus ayacahuite (Ehren.), Pinus pseudostrobus (Lindl. var pseudostrobus), Pinus hartwegii (Lindl.), Quercus glabrescens (Humb et Bonpl.), Quercus obtusata (Bonpl.), Salix paradoxa (Kunth.), Clethra galeottiana (Briquet.) y Litsea glaucescens (Kunth.). Las especies identificadas como los vecinos más próximos fueron Q. laurina, P. ayacahuite, y P. pseudostrobus.
El rango altitudinal de la población estudiada se extiende entre 2720 a 3100 m, siendo el rango óptimo de abundancia entre 2875 y 3067 metros, según la función de Weibull (Figura 3A). La pendiente predominante del área oscila entre 13.71% y 33.73%, aunque la mayor concentración de la especie se observó en terrenos con pendientes de 15% a 20% (Figura 3B). En contraste, se observaron bajos índices en terrenos con inclinaciones inferiores al 10% o superiores al 40% (Figura 3B). Se destaca que se observaron ondulaciones y concavidades del terreno a escasos cientos de metros con exposiciones geográficas y pendientes contrastantes.
Alrededor de los árboles centrales, se identificaron en total 185 individuos de regeneración natural. De ese total, el 52% se clasificó como latizales y el 48% como brinzales. La altura total de los 185 individuos varió entre 10 cm y 11 m, aunque en su mayoría tenían una altura inferior a un metro, representando poco más del 45% del total registrado (Figura 3C y 3D). Como se esperaba, debido a la competencia, a medida que la altura total aumentaba, la proporción de individuos disminuyó gradualmente (Figura 3D).
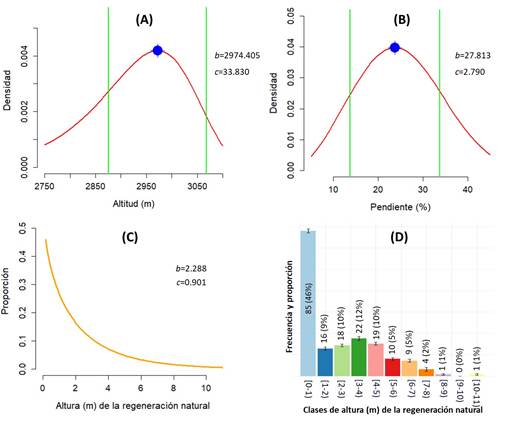
Figura 3 Curvas de densidad de probabilidad ajustadas para la altitud (A) la pendiente (B), la altura de la regeneración natural de A. hickellii (C) y la distribución de frecuencias de la altura total de la regeneración natural (D). En los paneles A, B y C, “b” representa el parámetro de escala y “c” representa el parámetro de forma del modelo de Weibull.
Figure 3. Fitted probability density curves for altitude (A), slope (B), height of natural regeneration of A. hickelii (C), and frequency distribution of the total height of natural regeneration (D). In panels A, B, and C, "b" represents the scale parameter and "c" represents the shape parameter of the Weibull model.
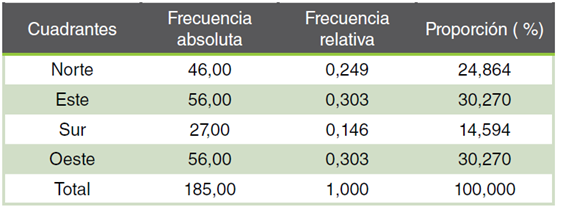
Al contrastar los intervalos de confianza del 95% en función del azimut se observó que, independientemente de la topografía, al menos uno de los cuadrantes presentaba una proporción significativamente diferente a la del resto. La Figura 4 ilustra que las proporciones difieren en los cuatro cuadrantes. El intervalo de confianza del cuadrante sur no se superpone con los de los cuadrantes este y oeste. Además, se observa una superposición parcial entre el cuadrante sur y el cuadrante norte. Tomando en cuenta lo anterior, la regeneración en la población estudiada no se distribuye de manera uniforme alrededor de los árboles de referencia: más del 80% se localizó en los cuadrantes norte, oeste y este, mientras que solo el 14.6% se registró en el cuadrante sur (Tabla 1).
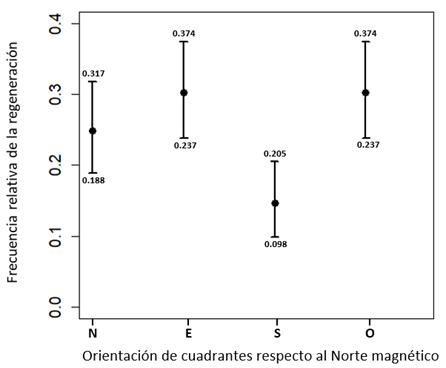
Figura 4 Intervalo de confianza de la proporción de la regeneración natural de Abies hickelii (Flous & Gaussen) en los cuadrantes norte (N), este (E), sur (S), oeste (O), alrededor de cada árbol madre.
Figure 4. Confidence interval of the proportion of natural regeneration of Abies hickelii (Flous & Gaussen) in the northern (N), eastern (E), southern (S), and western (W) quadrants around each mother tree.
Cuadro 1 Proporción de la regeneración natural de Abies hickelii (Flous & Gaussen) en los cuatro cuadrantes.
Se observó que la topografía local influye en la densidad y distribución de los individuos estudiados. En particular, no se encontraron árboles adultos que cumplieran con los criterios para ser considerados árboles madre en laderas orientadas al sur, oeste, sureste o suroeste. En cambio, estos árboles se encontraron únicamente en terrenos orientados hacia el norte, noreste, noroeste y este (Cuadro 2). Bajo el dosel de los árboles adultos, la regeneración natural parecía seguir un patrón similar, con la mayor concentración de regeneración en terrenos orientados hacia el noreste (69%), este (22%), norte (6%) y noroeste (3%).
Al analizar la exposición del terreno, es decir, la orientación de las laderas donde se encuentran los árboles de referencia, se observaron ligeras variaciones en la proporción de regeneración en los cuatro cuadrantes. Por ejemplo, cuando los árboles madre estaban situados en laderas expuestas al noroeste, el cuadrante con mayor número de registros de la regeneración fue el oeste, seguido del norte y el este. En contraste, en terrenos expuestos al este, el 50% de la regeneración se encontró en el cuadrante este, el 25% en el sur, el 17.5% en el norte y el 7% en el cuadrante oeste. Se concluye que los atributos topográficos como la exposición de la ladera, la pendiente y la altitud desempeñan un papel crucial en la disposición espacial de los individuos estudiados, según estos resultados.

Discusión
El estudio de los patrones espaciales requiere de esfuerzos de muestreo y de estrategias de gestión apoyadas en el conocimiento científico (23). Bajo este contexto, comprender la distribución espacial de una población en riesgo como Abies hickelii (Flous & Gaussen) es de vital importancia para fortalecer los mecanismos y estrategias de conservación.
Se ha definido que la dispersión de las plantas jóvenes alrededor de un árbol semillero resulta de una compleja interacción entre la dispersión de semillas, la competencia por recursos, las condiciones ambientales y las interacciones biológicas. Los patrones observados, como parches o manchas circulares, son el resultado de, cómo estas fuerzas se combinan y afectan la distribución de las plántulas en el paisaje. Parte de este fenómeno es explicado por la teoría del equilibrio entre competencia y dispersión (24), (25).
No obstante, en un entorno de alta competencia intraespecífica como el caso de A. hickelii y, sobre todo, donde la competencia es marcadamente asimétrica como fue observada por Antúnez et al. (26), se esperaría una distribución más o menos uniforme sin discrepancias significativas entre un cuadrante y otro, para optimizar los espacios alrededor del árbol madre. Y en ese sentido, el azimut por sí solo, no debería corresponderse con la disposición de la regeneración, si se asume que el árbol madre ocupa el centro de dispersión de las semillas. Sin embargo, los resultados evidenciaron que la proporción de la regeneración de al menos un cuadrante era significativamente diferente que el resto.
La proporción de individuos en el cuadrante sur fue significativamente inferior que los cuadrantes este y oeste (Figura 4). Debe notarse que la comparación de intervalos de confianza de las proporciones (1-95%) en este contexto es más riguroso que la comparación de medias, al proporcionar un rango de valores posibles para el parámetro estimado (proporción). En este caso, se consideró que había diferencia significativa si los extremos (límites confidenciales) no se traslapan.
A escala local, la dispersión depende en gran medida de los recursos disponibles (e. g. agua, nutrientes, luz). Cuando estos recursos son limitados, de acuerdo con la teoría de la competencia y la dispersión de semillas, los nuevos individuos tienden a ser agregada en lugar de uniforme (27). De manera similar, las características del terreno, incluyendo la exposición, pendiente; la variabilidad en la disponibilidad de recursos, y la influencia de factores bióticos y abióticos suelen hacer que la distribución sea más irregular (28). ¿Pero exactamente de qué manera se asocian estos factores con el azimut?
Si la densidad de la regeneración natural es alta cerca del árbol madre, los recursos pueden agotarse rápidamente, reduciendo el crecimiento de nuevas plantas cerca del árbol semillero. Esto puede llevar a una mayor densidad de plantas jóvenes en áreas donde los recursos son relativamente más abundantes. Sin embargo, con la información recabada, no se logra explicar si la disponibilidad de nutrientes se asocia con el rumbo o el azimut en la población estudiada. Otra posible explicación a lo observado podría asociarse con la disponibilidad de la luz solar. En ambientes densamente vegetados como se constató en campo, la competencia por luz puede influir en la distribución de las plántulas. Las plantas jóvenes (regeneración natural) que germinan cerca de la sombra del árbol madre pueden estar en desventaja y, por lo tanto, sería más probable que las plántulas se distribuyan hacia alguna orientación. De ser esta la explicación, el género Abies no debe considerarse tolerante a la sombra como lo sugiere Bautista-Sampayo (29) si no intolerante como lo menciona López-Mata, (30) o al menos medianamente tolerante. Para entender mejor este fenómeno, hace falta más estudios de individuos en fases iniciales de crecimiento de esta población, incluidos la competencia en el sistema radicular y buscar una explicación sobre el papel del azimut en la distribución de la regeneración natural de A. hickelii.
Otro aspecto importante encontrado en este estudio fue el efecto de la inclinación de la ladera en la frecuencia de la regeneración. De acuerdo con Valverde-Padilla (31) y López-Gómez et al. (32), en el hemisferio norte, las laderas con exposición Sur reciben casi seis veces más radiación solar que las laderas con orientación norte. Por consiguiente, las laderas con exposición norte son más húmedas en comparación con las orientadas al sur. En ese sentido, Ávila et al. (33) reportan que la distribución ecológica del género Abies se distribuye en terrenos con exposición N, NE, NO, SE y SO. Los mismos autores reportan una alta presencia de Abies en terrenos con exposición S, E y O. De manera similar, en un estudio realizado en el Pico de Orizaba por Ávila-Bello y Mata (3), se reporta que el bosque de A. hickelii (Flous & Gaussen) es mejor conservado en terrenos con exposiciones norte, y de manera esporádica en laderas con exposición sur; en tanto que, en las cumbres de Acultzingo, la mayor proporción se observó en exposiciones sureste y suroeste (3). De esa manera, dichos resultados contrastan con los hallazgos reportados en este trabajo. Este último atribuible a condiciones locales, a decir de los autores.
De acuerdo con Nieto de Pascual (34), en un estudio sinecológico del bosque de oyamel de la cañada de contreras (Ciudad de México) se observaron poblaciones más abundantes hacia la exposición suroeste, y en menor medida en sitios con otras exposiciones. Finalmente, un estudio realizado en el Cofre de Perote, estado de Veracruz, México, por Sánchez-Velásquez et al. (35) se reporta una distribución de A. religiosa en sitios con exposición noroeste (47% del total), aunque el mayor registro fue en lugares con exposiciones suroeste, oeste, noroeste y noreste.
Por otro lado, Madrigal-Sánchez (36) reporta registros nulos en las exposiciones sur y sureste, y registros de presencia en exposiciones noroeste y noreste, en el Valle de México. De esta manera, varios estudios relacionados con las preferencias topográficas de A. hickelii (Flous & Gaussen) coinciden parcialmente con los resultados reportados en esta investigación, como es el caso de Ávila et al. (33), Ávila-Bello et al. (3), Sánchez-Velásquez et al. (35) y Nieto de Pascual (34). Estas variaciones podrían atribuirse a diferentes factores incluyendo la presencia y dirección de vientos dominantes, el grado de inclinación del terreno, la presencia de la fauna dispersora de semillas, entre otros factores. Otro de los aspectos a considerar es la microtopografía o la topografía local, uno de los factores determinantes en la distribución espacial de las plantas con distribución restringida, tal como fue señalado por Antúnez et al., (37). Esta afirmación coincide con los resultados de este estudio (Tabla 2).
Conclusiones
Los resultados del estudio revelaron que la distribución de las plántulas no sigue un patrón uniforme alrededor del árbol madre, sino que presenta variaciones significativas en diferentes cuadrantes en función del azimut y la exposición de la ladera. Más del 80% de la regeneración natural de la población estudiada se encuentra concentrada en los cuadrantes norte, oeste y este, mientras que sólo el 14.6% se ubica en el cuadrante sur. Esto lleva a rechazar la hipótesis de igualdad de proporciones planteada al inicio de la investigación. Por otro lado, la topografía local y la exposición al sol también juegan un papel importante en la distribución de la regeneración natural. Si se considera la exposición geográfica de los árboles madres, la mayor concentración de la regeneración natural se produce en el noreste (69%), el este (22%) y el norte (6%). Estos hallazgos subrayan la necesidad de considerar estos factores en las estrategias de conservación para Abies hickelii, incluyendo posibles aclareos selectivos y la protección de áreas con condiciones óptimas para la regeneración. La investigación futura debería centrarse en comprender mejor cómo estos factores interactúan y afectan la dinámica de la población, especialmente en las etapas iniciales de crecimiento.

