











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La fragmentación de hábitat induce cambios importantes en el paisaje, lo cual a su vez genera cambios en la composición de las poblaciones faunísticas, y en los procesos ecológicos en los que estos están involucrados (Püttker et al., 2020). Como resultado, muchas especies tienden a disminuir su tamaño poblacional, pierden gran parte de la diversidad genética, y reducen su distribución geográfica, facilitando la extinción local (Lino et al., 2019). Además, el efecto de borde tiende a ser muy frecuente, lo que favorece el aumento en la depredación de nidos, el parasitismo, y la competencia (Seaman & Sculze, 2010).
Una manera de contrarrestar los efectos de la fragmentación de hábitat ha sido la creación de corredores biológicos, los cuales cumplen funciones como servir de hábitat para varias especies, facilitar el desplazamiento de elementos (animales, plantas, nutrientes, semillas), además, de permitir la expansión y distribución de las especies en matrices anexas y dar refugio a elementos o especies particulares (Calvo, 2009). Por otro lado, los corredores asociados a cuerpos de agua, ayudan a conservar y purificar este recurso (Cerra & Crain, 2016), mientras que aquellos que se establecen en zonas como calles arboladas o vías de tren, amortiguan los efectos de barrera causados por los sistemas de transporte, además de que regulan condiciones ambientales en microclimas y purifican el aire (Yueguang, 2003). Entre las modalidades que se han creado de corredores biológicos se encuentran los corredores interurbanos, los cuales conectan tramos verdes dentro de las ciudades con otros ecosistemas, incluyendo dimensiones culturales, socioeconómicas y políticas (Decreto Ejecutivo N°40043-MINAE, 2016).
Las aves por su parte, han sido utilizadas en muchas investigaciones como organismos bioindicadores, es decir, que mediante su presencia se indica el nivel de preservación en el que se encuentra un hábitat o un ecosistema (Pimenta et al., 2009). Esto hace posible que dichos organismos, sean correlacionados con variables del ecosistema que permiten su sobrevivencia a través de la determinación de la riqueza y la abundancia (Hidalgo, 2020). Además, los patrones de abundancia en aves, son indispensables en los procesos ecológicos, ya que contribuyen a servicios ecosistémicos como dispersión de semillas, polinización, control de plagas, sanidad ambiental, regulación en el ciclado de nutrientes y transmisión de enfermedades, así como también proveen de alimento a otras especies (Sica, 2016).
La avifauna ha sido un objeto de estudio esencial en el manejo, implementación y monitoreo de los corredores biológicos a nivel nacional. Por ejemplo, se ha estudiado como se afecta la riqueza de aves a partir del manejo de cercas vivas ubicadas en Turrialba, Cartago, dentro del Corredor Biológico Volcánica Central-Talamanca (Garbach et al., 2010). Además, uno de los principales motivos por el que se crea el Corredor Biológico San Juan la Selva, es para proteger a la lapa verde (Ara ambiguus) la cual se encuentra bajo amenaza, y también se favorece la migración de especies de aves y mariposas (Villate et al., 2009). De la misma manera, se ha estudiado la riqueza y abundancia avifaunísitca asociada a la microcuenca urbana del río Bermúdez en Heredia, caracterizando a las especies por su grado de dependencia al bosque y gremios tróficos (McQueen-Blanco et al., 2019).
Dentro de los trazados que comprende el Corredor Biológico Interurbano el Achiote, se encuentran distintos tipos de coberturas de uso del suelo como asentamientos urbanos, cultivos de café y caña, así como también bosques secundarios y ribereños. Sin embargo, actualmente no se tienen datos respecto a la avifauna del sitio, ni de como esto puede llegar a variar dentro de los distintos ecosistemas y su contribución al funcionamiento del corredor. Es por esto que el objetivo de esta investigación se basa en caracterizar el ensamblaje de aves asociada a los distintos tipos de coberturas de uso del suelo, a partir de sus requerimientos ecológicos, para el mejoramiento de la conectividad funcional en el corredor biológico.
Materiales y métodos
Área de Estudio
El Corredor Biológico Interurbano el Achiote, se ubica en la provincia de Alajuela, y comprende algunos distritos del cantón de Grecia como Puente de Piedra, San José, San Roque, San Isidro, Tacares y Grecia. Además, incluye una pequeña parte del distrito de San Juan, perteneciente al cantón de Poás. El corredor biológico cuenta con una superficie de 55,23km2, en donde se seleccionaron diez puntos de muestreo, dos por cada tipo de cobertura de uso del suelo, los cuales fueron: cultivo de café, cultivo de caña de azúcar, zonas urbanas, bosque secundario y bosque de ribera.

Muestreo de ensambles de aves
Los muestreos se realizaron dos veces por mes, durante seis meses (tres en época seca y tres en época lluviosa), para un total de 120 muestreos, lo que incluye las épocas reproductivas y migratorias (latitudinales y altitudinales) de la avifauna (Sandoval, 2016). Los muestreos se realizaron en dos períodos del día: el primero entre las 5am y las 10am, y el segundo entre las 3pm y las 6pm, lo que coincide con los picos de actividad de las aves diurnas (González-García, 2011).
Para el registro de las especies de aves, utilizamos la técnica de puntos de conteo de radio variable, con cinco puntos por cada sitio de muestreo, con una separación mínima de 150m entre cada punto (Rivero et al., 2015). En cada punto de conteo, durante períodos de 10min registramos la riqueza y abundancia de aves de manera visual y auditiva. Incluimos, también, todas las especies de aves que fueron observadas o escuchadas entre los puntos de muestreo, esto con el fin de abarcar la mayor riqueza posible dentro del área.
Caracterización del ensamblaje de aves
Realizamos la caracterización con base en la determinación de la Riqueza y la Abundancia relativa calculada la cual se expresa en la siguiente fórmula:

Por otra parte, también investigamos sobre el hábitat de preferencia para cada especie (Stiles & Skutch, 2007): hábitats urbanos (zonas grises, zonas verdes urbanas, zonas de ribera), hábitats abiertos o alterados (charrales, potreros y cultivos, arboledas abiertas), y hábitats boscosos (bosque de ribera, sotobosque, dosel y bordes de bosque). Además, se determinó el grado de dependencia al bosque basado en la siguiente clasificación (Sandoval & Barrantes, 2009):
Especies que requieren amplia cobertura de bosque secundario avanzado con muy poca o nula intervención.
Especies que toleran bosques secundarios intervenidos, con una cobertura boscosa mínima del 50%.
Especies capaces de sobrevivir en bosques fragmentados y alterados, que poseen menos del 50% de cobertura boscosa.
Especies que requieren de la presencia de árboles asociados a las áreas abiertas, como zonas de cultivos o potreros con sombra.
Especies que no requieren áreas boscosas, y pueden sobrevivir en charrales, cercas vivas, zonas de cultivos, o áreas urbanas.
De la misma forma determinamos el gremio trófico de las especies de aves, de acuerdo a los ítems alimenticios que conforman su dieta (Marateo & Arturi,2013), los cuales son: insectívoro, granívoro, frugívoro, nectarívoro, omnívoro, piscívoro, carnívoro, carroñero y folívoro. Finalmente, tomamos en cuenta las categorías de amenaza para cada especie, con base en los datos suministrados por la Unión Internacional para la Conservación de la Naturaleza (Unión Internacional para la Conservación de la Naturaleza (UICN), 2012), clasificándolas según lo estipulado en los criterios de la Lista Roja de la siguiente manera: no evaluada, en peligro crítico, en peligro, vulnerable, casi amenazada y preocupación menor.
Análisis estadístico
Mediante el software de R (versión 4.2), en primer lugar, realizamos un análisis de Chi-cuadrado, para comparar la abundancia relativa teórica, y también el tipo de hábitat de preferencia. Seguidamente, mediante un Análisis de Varianza (ANOVA) diseñado por bloques, se evaluó si existen diferencias significativas para la cantidad de especies de aves clasificadas entre los tipos de cobertura de uso del suelo (factor bloque), con respecto a: los comportamientos que exhiben las especies (uso de hábitat), y los tipos de dependencia al bosque.
Para los datos de los gremios tróficos, se realizó la prueba de Brown-Forsythe, la cual es un ANOVA más robusto, seguido de una prueba de Kruskal-Wallis para comparar el agrupamiento de los gremios entre tipos de cobertura.
Por otra parte, realizamos análisis multivariados para evaluar la caracterización de los sitios de muestreo con base en los ensambles de aves, a través de una matriz biológica transformada según Hellinger. En primer lugar, realizamos una comparación entre los diez sitios de muestreo para analizar el grado de similitud entre estos con respecto al ensamblaje de aves, mediante una técnica de ordenación por Clusters evaluada mediante un índice de correlación cofenética, a partir de las reglas de similitud de Bray-Curtis. Seguidamente, mediante un diagrama de Venn, representamos las especies de aves que se distribuyen entre los grupos de sitios obtenidos mediante el Cluster. En segundo lugar, determinamos si existen diferencias significativas en la composición biológica de las especies de aves, con respecto a los tipos de cobertura de uso del suelo, mediante un Análisis de Permutación de la Varianza Multivariado (PERMANOVA).
Resultados
Riqueza y abundancia
Para el corredor biológico registramos un total de 194 especies de aves silvestres, agrupadas en 42 familias y 17 órdenes (Tabla 1 en Apéndice). El orden Passeriformes fue el más abundante con 123 especies. Las familias más representativas en cuanto a riqueza fueron Tyrannidae con 21 especies, seguida de Parulidae con 18 especies, Trochilidae con 14 especies y Thraupidae con 13 especies. En cuanto a la abundancia relativa calculada, el perico barbinaranja (Brotogeris jugularis), fue la especie más abundante con el 20,9% de los registros, seguida de la golondrina azul y blanco (Pygochelidon cyanoleuca) representando un 8,04%, y el loro frentirrojo (Psittacara finschii) con 7,41% (Fig. 2).

Hábitat de preferencia
La distribución de las especies de aves con respecto al tipo de hábitat presentó diferencias significativas (x2= 198,91, gl= 9, pval< 0,05). La mayoría de las especies se agruparon en hábitats abiertos (Fig. 3). Por otro lado, en los hábitats con mayor cobertura boscosa, las aves teóricamente presentan preferencia hacia las zonas de borde. Por su parte, en las zonas más urbanizadas las aves prefieren más los hábitats asociados a las áreas verdes y en los márgenes de ríos y quebradas.

Grado de dependencia al bosque
Las categorías de dependencia al bosque más abundantes entre las especies de aves son las menos dependientes (tipos cuatro y cinco), en contraposición con las aves más dependientes (tipos uno y dos) (F=9,15, gl=4, p< 0,05) (Fig. 4). Sin embargo, en las categorías altamente dependientes, las especies se concentran en los bosques secundarios, los cuales presentaron un total de 36 especies (tipo uno y dos), mientras que, en el uso del suelo urbano estuvieron ausentes, y solamente 5 especies fueron registradas para los cañales y los cafetales. Algunas de las especies poco comunes, y altamente dependientes del bosque para el corredor, son: pava negra (Chamaepetes unicolor), paloma perdiz (Zentrygon costaricensis), colibrí montañés gorgimorado (Lampornis calolaemus), lechuza serranera (Megascops clarkii), codorniz moteada (Odontophorus guttatus), saltón cabecicastaño (Arremon brunneinucha) y el quetzal (Pharomachrus mocinno).

Gremios tróficos
La composición de los gremios muestra una tendencia similar entre los tipos de cobertura de uso del suelo (KW=0,72, gl=4, p> 0,05), aunque entre los tipos de gremios si se encontraron diferencias significativas (F=33,73, gl=8, p< 0,05) (Fig. 5). Los gremios predominantes fueron los insectívoros (Tyrannidae, Parulidae, Vireonidae, Picidae, Dendrocolaptidae, Troglodytidae) y frugívoros (Turdidae, Cracidae, Trogonidae, Pstittacidae, Thraupidae e Icteridae), en contraste con grupos más reducidos como carroñeros (Cathartidae), nectarívoros (Trochilidae), omnívoros (Ramphastidae, Fringillidae, Thraupidae, Icteridae, Momotidae), granívoros (Columbidae, Passerellidae, y Cardinalidae) y carnívoros (Falconidae, Accipitridae y Strigidae), los cuales registraron menos especies. Para el gremio trófico piscívoro, únicamente se registraron dos especies dentro del bosque de ribera, las cuales fueron el martín pescador verde (Chloroceryle americana), y el martín pescador collarejo (Megaceryle torquata), mientras que los folívoros fueron representados por los sinsontes (Saltator coerulescens y Saltator maximus), así como la codorniz crestada (Colinus cristatus), junto con otras 6 especies (Psittacidae y Columbidae).

Estado de amenaza
De las 194 especies de aves registradas para el corredor, 184 se encuentran categorizadas bajo preocupación menor. Se registraron seis especies casi amenazadas: pava negra (Chamaepetes unicolor), pibí boreal (Contopus cooperi), pinzón cafetalero (Melozone cabanisi), quetzal (Pharomachrus mocinno), el zacatero común (Sturnella magna), y la reinita alidorada (Vermivora chrysoptera). El perico frentinaranja (Eupsittula canicularis) se encuentra en estado vulnerable, mientras que la lora de nuca amarilla (Amazona auropalliata) se categoriza bajo peligro, siendo esta la especie más amenazada dentro del corredor biológico. Además, para el colibrí orejivioláceo verde (Colibri cyanotus), y el carpintero lineado (Dryocopus lineatus), no se encontraron datos sobre su condición.
Análisis espacial
los ensambles de aves varían de acuerdo con los tipos de cobertura de uso del suelo dentro del corredor biológico (F=3,24, gl=4, r2=0,72, p< 0,05). Mediante el análisis del Cluster, con una correlación cofenética del 0,906, señala que los sitios con más similitud en cuanto a la comunidad de especies de aves que albergan, se clasifican en 3 grupos (Fig. 6): Grupo A con los sitios uno y dos (bosque secundario), el grupo B con los sitios más alterados que fueron el sitio cinco (cafetal con poca sombra), sitio siete (cañal sin sombra), y los sitios seis y ocho (urbanos), y finalmente el grupo C con los sitios más arbolados, que fueron los sitios cuatro y nueve (bosque de ribera), junto con el sitio diez (cafetal sombreado) y el sitio tres (cañales y potreros sombreados).
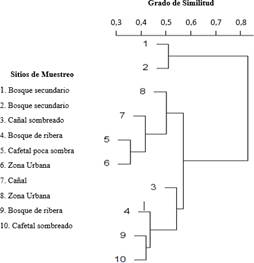
Fig. 6 Agrupamiento por ligamiento promedio de los sitios de muestreo según la comunidad avifaunística dentro del Corredor Biológico Interurbano el Achiote, Grecia, Alajuela, Costa Rica.
El grupo A contiene 94 especies de aves, de las cuales 18 comparte con el grupo B y 37 con el grupo C. Por su parte, el grupo B cuenta con 88 especies de aves, de las cuales comparte 57 con el grupo C, dejando únicamente 3 especies exclusivas de los sitios más alterados del corredor. Finalmente, el grupo C cuenta con la mayor cantidad de especies, con un total de 144, de las cuales 40 son exclusivas y no se comparten (Fig. 7). Únicamente 10 especies se traslapan entre todos los grupos.

Discusión
Con respecto a la abundancia relativa calculada, el perico barbinaranja (Brotogeris jugularis), junto con el loro frentirroja (Psittacara finschii), fueron registradas como las especies de psitácidos más abundantes dentro del corredor biológico. Lo anterior se debe a que son especies gregarias que prefieren generalmente las áreas abiertas (Stiles & Skutch, 2007), pero que también pueden presentarse dentro de ecosistemas urbanos en altas densidades junto con otras especies como la paloma de Castilla (Columba livia), el zanate (Quiscalus mexicanus), la paloma aliblanca (Zenaida asiatica) y la viudita (Thraupis episcopus) (Fallas, 2016).
La mayoría de especies de aves presentan una preferencia por los hábitats abiertos, seguidos por los boscosos y por último los urbanos. Quiñonez &Hernández (2017), indican que dicha preferencia se atribuye a las características que cada hábitat ofrece (fuentes de alimento y refugio), junto con la distribución que presenta cada especie. Agudelo-Rendón et al., (2021), señala que las especies asociadas a las áreas abiertas presentan rasgos generalistas, con capacidades ventajosas de utilizar diferentes recursos alimenticios, y de desplazarse por zonas fragmentadas. Además, Guevara (2021) indica que las especies migratorias evitan los interiores del bosque, y prefieren las áreas alteradas en búsqueda de zonas próximas y accesibles de alimentación, refugio y descanso, lo cual aumenta la riqueza de aves en este tipo de hábitats.
Wang et al. (2014), señala que la diversidad de aves es grande en las zonas boscosas, debido a la heterogeneidad del hábitat por la presencia de plantas epífitas, y el sotobosque, aparte de que presentan protección contra ciertos depredadores (Melo et al., 2020). Sin embargo, en las zonas urbanas la riqueza de aves observada fue la más baja. Leveau (2018), indica que los factores que afectan la riqueza avifaunística en las zonas urbanas, son principalmente la poca disponibilidad de hábitat y alimento por la escasa vegetación, las variaciones en humedad, temperatura, y la cantidad de luz. La ausencia de vegetación nativa es algo que también influye sobre las aves en las zonas urbanas (Araya & Carvajal,2019), así como la intolerancia al ruido antropogénico (Romero, 2015). Lo anterior, caracteriza a las especies de aves en zonas urbanas por ser generalistas y altamente adaptables a los disturbios antropogénicos.
Con respecto al grado de dependencia al bosque, las especies de aves más abundantes en todos los tipos de cobertura, son las que presentan una dependencia intermedia. Estos resultados fueron similares al estudio realizado por Sandoval (2019), el cual también presentó un mayor número de especies de aves con dependencias intermedias al bosque, mientras que los tipos de dependencia baja y alta, fueron las que menos especies presentaron para la avifauna presente en Getsemaní, Heredia. Por otra parte, Biamonte et al. (2011), explican que las especies con alta dependencia al bosque son raras, dado que la expansión agropecuaria dentro del Valle Central, junto con el desarrollo urbanístico, causaron una reducción de manera significativa en las coberturas del bosque secundario. Por esta razón, la mayoría de especies altamente dependientes, se confinan para el caso del corredor biológico, en la Reserva Forestal de Grecia.
Considerando los gremios tróficos, resultados obtenidos por Zurita & Bellocq (2012), indican que los insectívoros son exitosos en diferentes áreas, ya que presentan bastante abundancia y disponibilidad de alimento, el cual se mantiene constante a lo largo del año. Por otro lado, la dominancia de especies frugívoras en varios tipos de cobertura, puede deberse a la presencia de especies de plantas ornitócoras, de las cuales muchas especies de aves, se logran aprovechar (Salazar-Ramírez et al., 2014). Sin embargo, en zonas urbanas, la composición de los gremios en el corredor biológico se comporta de una manera similar al resto de tipos de cobertura. En estudios realizados por Marigliano et al. (2009) y Coria et al. (2015), también son los insectívoros junto a los granívoros los gremios más abundantes en las ciudades, esto debido a la alta abundancia de especies como pinzones o palomas.
Por otro lado, los nectarívoros fueron constantes en todos los tipos de cobertura, lo cual se relaciona con la presencia de flores ornitófilas que contribuyen de forma significativa a su abundancia y distribución (Salazar-Ramírez et al., 2014). Los carroñeros por su parte fueron muy abundantes en todos los tipos de cobertura, representados por dos especies: el zopilote cabeza negra (Coragyps atratus), y cabeza roja (Cathartes aura). Fandiño et al. (2017), explica en sus resultados que estas 2 especies de aves son relativamente abundantes, al tener la capacidad de volar sobre varios tipos de cobertura (sobre todo en presencia de corrientes termales), y aprovecharse de la presencia de árboles y postes para perchar, de la misma manera que lo hacen las rapaces del gremio carnívoro. Sin embargo, en los cañales del corredor biológico fue más frecuente encontrar especies de rapaces. López (2020), explica en su estudio que es más fácil encontrar presas en campos agrícolas y pastizales, razón por la cual hay una mayor abundancia de este gremio en hábitats alterados, comparado con hábitats de bosque o inclusive urbanos.
Resultados de McQueen-Blanco et al., (2019), indican que las zonas urbanas favorecen la dominancia de aves generalistas. Sin embargo, para el corredor biológico las especies omnívoras fueron constantes entre los tipos de cobertura, posiblemente porque pueden expandir su dieta a otros ítems como granos, frutos, insectos, y néctar, cuando hay escasez. A su vez, las especies granívoras también fueron constantes, ya que las semillas, granos cultivados y material vegetal, puede ser encontrados en diferentes hábitats (Carpio et al., 2016). Además, los gremios menos frecuentes dentro del corredor biológico fueron los piscívoros y folívoros. Varios estudios concuerdan en que las aves piscívoras son más frecuentes en cercanías a cuerpos de agua, al proveer alimentación, estadía y reproducción (Gómez et al., 2016). Para el corredor biológico, los bosques de ribera constituyen el único hábitat principal para los martines pescadores. Por otra parte, Botero- Delgadillo & García (2011) mencionan que la folivoría en las aves es un hábito infrecuente y poco documentado, posiblemente por el alto contenido de fibra presentes en las hojas, las cuales son difíciles de metabolizar.
Para las categorías de amenaza registradas, las especies más amenazadas, dos son psitácidos (Eupsittula canicularis y Amazona auropalliata). En el mercado y tráfico ilegal de fauna, los psitácidos están entre las especies más vendidas y fáciles de obtener (Padilla & Meza, 2021). En Oaxaca, México, se contabilizó un total de 245 psitácidos obtenidos de forma ilegal, en donde se registró que Eupsittula canicularis fue la especie en cautiverio más común (Buenrostro-Silva et al., 2021). Además, la pérdida de hábitat constituye otra causa de amenaza para muchas especies de aves. El quetzal (Pharomachrus mocinno), y el pinzón cafetalero (Melozone cabanisi) una de las especies de aves endémicas para Costa Rica (Asociación Ornitológica de Costa Rica, 2020), presentan hábitats muy restringidos por la fragmentación y la expansión urbanística (Muñoz et al., 2021).
Con respecto a la comparación entre los sitios de muestreo con la comunidad avifaunística asociada, el mayor traslape de especies ocurre entre los sitios del grupo C (bosque de ribera y cafetal sombreado), con el grupo B (sitios urbanos y cañales). Este traslape, indica que la mayoría de estas especies de aves son generalistas, y presentan una alta capacidad de buscar diferentes opciones de recursos ecológicos en varios sitios (Londoño-Betancourth, 2011). Resultados obtenidos por Agudelo-Rendón et al. (2021), indican que las aves de los sitios urbanos, logran utilizar gran variedad de áreas abiertas para encontrar recursos, mientras que los bordes de bosque y las quebradas los utilizan principalmente para su desplazamiento a lo largo del paisaje. De la misma manera, el sitio con cafetal sombreado presenta bastante similitud con respecto a los bosques de ribera. Sánchez- Clavijo et al. (2020), explica en su estudio, que los cafetales con sombra actúan similar a un bosque, suplementando recursos para gran cantidad de especies de aves, al presentar una estructura vegetal compleja. Por último, los sitios del grupo B son los que presentan la menor riqueza de aves, con únicamente 3 especies: la paloma de castilla (Columba livia) y el gorrión (Passer domesticus), en zonas urbanas, junto con la codorniz moteada (Colinus cristatus), en los cañales. Valdiri (2021), menciona que la baja riqueza encontrada en los cañaverales, se debe a razones como: una baja estructura y diversidad vegetal, el uso de agroquímicos, y el aprovechamiento intensivo de la producción agrícola, lo cual también se cumple en los cañales del corredor biológico.
En conclusión, las aves difieren según el uso del suelo. La mayoría de especies parecen preferir áreas abiertas y alteradas, ya que muchas presentan rasgos generalistas (alimentación y comportamientos) y mediana dependencia del bosque; además, tienen la capacidad de desplazarse por zonas fragmentadas.

