











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La pérdida de hábitat y la fragmentación constituyen dos de las principales amenazas a la biodiversidad (Wilkinson et al., 2018; Horváth et al., 2019). Estos procesos han provocado la pérdida de bosques prístinos, incluidas grandes áreas de los bosques tropicales de la región centroamericana (Hoang & Kanemoto, 2021). La pérdida de un 90% de la extensión de un hábitat particular implica que también cerca del 50% de las especies desaparezcan (García, 2002). El cantón de Abangares, Guanacaste, en el noroeste de Costa Rica ha sufrido pérdida de sus hábitats naturales debido a cambios en el uso de la tierra a causa de la expansión agrícola (Sierra et al., 2016).
La mayoría de los esfuerzos de conservación de la biodiversidad se centralizan en los sistemas nacionales de áreas silvestres protegidas (Pimm et al., 2018). No obstante, en la mayoría de los casos, las áreas protegidas estatales no son suficientes para contener un alto porcentaje de la biodiversidad a nivel local ni a nivel global (Pringle, 2017). Por ejemplo, la mayoría de las áreas no tienen el tamaño suficiente para contener poblaciones viables de mamíferos, lo que obedece a principios e hipótesis ecológicas básicas (Chase et al., 2020). Además, por lo general estas áreas protegidas se encuentran aisladas y son susceptibles a presiones de origen antrópico (Jones et al., 2018). Ante este panorama, las reservas naturales privadas surgen como herramienta complementaria a los esfuerzos gubernamentales y son de valioso aporte para la conservación de la biodiversidad (Cruz-Bazán et al., 2017). Estas iniciativas privadas y voluntarias protegen remanentes boscosos y en algunos casos forman parte de corredores biológicos que aportan conectividad ecológica (Cruz-Bazán et al., 2017). Bajo este principio la Reserva Sierra Zapote (RSZ) fue establecida con el objetivo de permitir la restauración ecológica natural, un proceso que comenzó en el año 2000 (Marín et al., 2021). En la actualidad la reserva protege 70ha de bosque secundario avanzado, bosque secundario en regeneración y bosque primario de galería (Marín et al., 2021).
Los mamíferos son un grupo de vertebrados que cumplen papeles ecológicos primordiales para el equilibrio y funcionamiento de los ecosistemas (Lacher Jr. et al., 2019). Estos animales contribuyen con la dispersión y depredación de semillas, la polinización de flores, la regulación de plagas y contribuyen con el ciclo de nutrientes en el ecosistema (Lacher Jr. et al., 2019). Algunos mamíferos son los depredadores más altos de las pirámides tróficas de varios ecosistemas del mundo y muchos tienen papeles ecológicos fundamentales en la dinámica de los suelos (Beca et al., 2022). Debido a los factores anteriores, así como sus requerimientos específicos de subsistencia, los mamíferos son bioindicadores del estado de sanidad del bosque (Serna-Lagunes et al., 2019). La ausencia o presencia de ciertas especies son indicativas del estado de conservación del ecosistema (Caro, 2003).
Los mamíferos medianos y grandes (llamados simplemente mamíferos en el resto del artículo) son grupos de especies que están entre los más amenazados por la pérdida y fragmentación del hábitat, debido a sus requerimientos de espacio, alimentación y refugio (de la Torre et al., 2012). Por este motivo los mamíferos requieren de grandes esfuerzos de conservación para mantener poblaciones viables (Ceballos et al., 2005). En este proceso es trascendental recuperar áreas deforestadas mediante estrategias de restauración ecológica (Haddad et al., 2015). Además, se requiere de conectividad entre los remanentes boscosos, protegidos o no, donde habitan los mamíferos para aumentar la dispersión y el flujo genético entre las especies (García, 2002). Desarrollamos este trabajo para conocer la riqueza y la abundancia relativa de estos mamíferos en la restauración vegetal de la Reserva Sierra Zapote en Costa Rica.
Materiales y métodos
Sitio de estudio
La Reserva Sierra Zapote está ubicada en el distrito de las Juntas del cantón de Abangares, Guanacaste, en el Bosque Húmedo Tropical de bajura del noroeste de Costa Rica (10°19'0.4"N, 84°59'21"W y 10°18'58"N, 84°59'8.9"W; Fig. 1.). El sitio se compone de 70ha de bosque secundario, bosque primario de galería y bosque en regeneración. Contiguo a la reserva prevalecen agropaisajes de pastos y fragmentos de bosque secundario (Marín et al., 2021).
El área ocupada por la reserva fue destinada para fines ganaderos hasta 1985 cuando fueron establecidos cultivos maderables con la siembra de teca (Tectona grandis Linn. F), melina (Gmelina arborea Roxb.) y pochote (Pachira quinata (Jacq.) W.S. Alverson). En el 2000 la propiedad fue dedicada a la protección de la naturaleza y se permitió que de ahí en adelante se diera la regeneración natural y asistida (Marín et al., 2021).
Se encuentra dentro del Bosque Húmedo Tropical (bh-T) según la clasificación del sistema de zonas de vida de Holdridge (Holdridge, 1967). Esta reserva tiene un ámbito altitudinal entre los 260 y los 350msnm (Ortiz-Malavasi, 2014). La precipitación promedio anual es de 2 500mm con cinco meses secos al año y una temperatura promedio de 22°C (Ortiz-Malavasi, 2014).
Métodos
El estudio lo ejecutamos desde mayo de 2020 hasta julio de 2021. Colocamos cinco cámaras trampa (dos Bushnell, dos Browning y una Alpha cam) a distancias variables (Fig. 1). La idea fue colocar las trampas en sitios estratégicos donde previamente visualizamos rastros de mamíferos como huellas, excretas, restos óseos y senderos entre otros (Fig. 1) según la metodología de estudios similares (e.g. Cruz-Jácome et al., 2015). Programamos las cámaras para que estuvieran activas durante las 24 horas del día y con un intervalo de 0,3 segundos entre cada disparo. Para cada fotografía o video registramos la fecha, hora, temperatura en ºC y la fase lunar. Seleccionamos el uso de cámaras trampa por ser una herramienta no invasiva y eficaz para registrar mamíferos de difícil detección o esquivos ante la presencia humana (Maffei et al., 2002; Monroy-Vilchis et al., 2009).
Revisamos las cámaras una vez cada uno o dos meses para recolectar los datos desde la memoria del dispositivo. Utilizamos el producto Obsession de Calvin Klein como atrayente dado que existe evidencia de ser un método útil para obtener más y mejores fotografías de los individuos (Mills et al. 2019).

Análisis de datos
Para estimar la riqueza específica de estos mamíferos en la reserva consideramos únicamente las especies claramente identificadas. La riqueza de mamíferos es la suma del total de especies registradas en la reserva (Moreno, 2001). Evaluamos la diversidad de los mamíferos de la reserva mediante el índice de diversidad de Shannon-Wiener (H'), para comparación con estudios similares en Costa Rica, el índice de dominancia de Berger-Parker y el estimador Chao-1 (Magurran, 1988). El índice de Berger-Parker describe sintéticamente la distribución de la abundancia de las especies de comunidades perturbadas (Caruso et al., 2007) y el índice de Chao indica el número de especies esperadas en el sitio basado en la abundancia de las especies detectadas (Magurran, 1988).
Determinamos la abundancia relativa con el número de registros fotográficos independientes (Monroy-Vilchis et al., 2011). Se consideran registros independientes a) fotografías consecutivas de diferentes individuos, b) fotografías consecutivas de individuos de la misma especie separados por un lapso de 24 horas, c) las fotografías no consecutivas de individuos de la misma especie (Monroy-Vilchis et al., 2011; Hernández et al., 2018). Para obtener el índice de abundancia relativa (IAR) de cada especie utilizamos la ecuación: IAR = (R / EM x 100), donde R= son los registros independientes, EM = es la intensidad de muestreo (número de cámaras/ días de muestreo) y 100 son días trampa como factor de corrección estándar para comparación con estudios similares (Monroy-Vilchis et al., 2011; Cusack et al., 2015).
Analizamos el patrón de actividad de los mamíferos de la reserva según las horas en que cada especie estuvo activa. Establecimos cuatro categorías de horario: diurno (0700-1659h), nocturno (1900-0459h), crepuscular matutino (0500-0659h) y crepuscular vespertino (1700-1859h) (Lira-Torres & Briones-Salas, 2012). Los horarios fueron clasificados por el programa Sun Times versión 7.1 (Kay & Du Croz, 2008). Clasificamos las especies como a) diurnas: cuando el 90% o más de sus registros fueron durante el día; b) mayormente diurnas: cuando de 70% a 89% de sus registros fueron durante el día; c) nocturnas: cuando el 90% o más de sus registros fueron durante la noche; d) mayormente nocturnas: cuando de 70% a 89% de sus registros fueron durante la noche; e) crepusculares: cuando 50% o más de sus registros fueron durante el periodo crepúsculo; y f) catemerales: sus registros fueron durante el día y la noche (Tattersall, 2006).
Para la evaluación de la actividad de los mamíferos según el ciclo lunar tomamos en cuenta a las especies categorizadas como nocturnas y mayormente nocturnas (Albanesi et al., 2016). Establecimos tres categorías de actividad, a) lunarfóbicas: especies cuyos registros fueron durante las fases de luna nueva, luna creciente y luna menguante cuando la intensidad lumínica es menor; b) lunarfílicas: especies con registros durante las fases de luna llena, gibosa creciente y gibosa menguante, cuando la intensidad lumínica es mayor; c) indiferentes: especies con un patrón disperso de actividad (Albanesi et al., 2016). Para ubicar una especie dentro de una categoría, debe tener al menos tres registros más que la otra categoría; con menos de tres registros de diferencia se considera indiferente (Albanesi et al., 2016).
Resultados
Obtuvimos 758 registros independientes de mamíferos con un esfuerzo de 2 135 días- trampa. Esos registros representan una riqueza de 19 especies de mamíferos pertenecientes a 14 familias en siete órdenes (Tabla 1). El orden más representado fue Carnívora (nueve especies), seguido de Rodentia (tres especies). Los órdenes menos representados fueron Cetartiodactyla, Pilosa y Primates con el registro de solo una especie cada uno. Es importante destacar la presencia común de perros domésticos (Canis lupus familiaris) en el área de estudio (16 registros, IAR = 0,75). Todas las 19 especies fueron identificadas sin ninguna duda y destacan tres felinos: el león breñero (Herpailurus yagouaroundi), el manigordo (Leopardus pardalis) y el puma (Puma concolor), el saíno (Pecari tajacu) y el tepezcuinte (Cuniculus paca) (Fig. 2). Estas dos últimas especies son de gran valor cinegético y muy apetecidas por los cazadores. Otras dos especies de mamíferos detectadas en la reserva tienen valor cinegético: el armadillo (Dasypus novemcinctus) y el zorro pelón (Didelphis marsupialis). El armadillo de zopilote (Cabassous centralis), la guatusa (Dasyprocta punctata) y la ardilla común (Sciurus variegatoides) también son cazadas por la gente para consumo, aunque sólo muy ocasionalmente. La diversidad de los mamíferos de la reserva fue de H' = 1,96, un valor del índice de Berger-Parker = 0,41 y el estimador de Chao-1 = 20.
Las especies más abundantes fueron Nasua narica (IAR = 14,6, N= 312) y Didelphis marsupialis (IAR = 6,1, N= 130). Las especies menos frecuentes en fotocapturas fueron Herpailurus yagouaroundi y Procyon lotor (IAR = 0,05, N= 1) (Tabla 1). Estos dos extremos muestran la gran diferencia que encontramos en el número de fotocapturas para las 19 especies de mamíferos en la reserva (Fig. 3).
Los patrones de actividad de los mamíferos de la reserva fueron los esperados. La mayoría de especies son de actividad nocturna y solo el saíno (Pecari tajacu), el pizote (Nasua narica) y la guatusa (Dasyprocta punctata) son de actividad mayoritariamente diurna (Fig. 4).
Tabla 1 Número de registros independientes (RI) de mamíferos medianos y grandes (según orden y familia taxonómica) con su respectivo índice de abundancia relativa (IAR) en la Reserva Sierra Zapote, Abangares, Guanacaste, Costa Rica.
| Orden | Familia | Especie | Nombre común | RI | IAR |
| Carninora | Canidae | Canis latrans | Coyote | 8 | 0,37 |
| - | Mephitidae | Conepatus semistriatus | Zorrillo | 16 | 0,75 |
| - | - | Spilogale angustifrons | Zorrillo manchado | 9 | 0,42 |
| - | Mustelidae | Eira barbara | Tolomuco | 3 | 0,14 |
| - | Felidae | Leopardus pardalis | Manigordo | 25 | 1,17 |
| - | - | Puma concolor | Puma | 4 | 0,19 |
| - | - | Herpailurus | León breñero | 1 | 1,05 |
| - | - | yahouaroundi | - | - | - |
| - | Procyonidae | Procyon lotor | Mapache | 1 | 0,05 |
| - | - | Nasua narica | Pizote | 312 | 14,61 |
| Primates | Cebidae | Cebus imitator | Mono carablanca | 6 | 0,28 |
| Cingulata | Dasypodidae | Dasypus novemcinctus | Cusuco | 74 | 3,47 |
| - | Chlamyphoridae | Cabassous centrali | Armadillo zopilote | 5 | 0,23 |
| Cetart. | Tayassuidae | Pecari tajacu | Saíno | 19 | 0,89 |
| Pilosa | Myrmecophagidae | Tamandua mexicana | Oso chaleco | 26 | 1,22 |
| Rodentia | Cuniculidae | Cuniculus paca | Tepezcuite | 6 | 0,28 |
| - | Dasyproctidae | Dasyprocta punctata | Guatusa | 79 | 3,7 |
| - | Sciuridae | Sciurus variegatoides | Ardilla | 5 | 0,23 |
| Didelph. | Didelphidae | Didelphis marsupialis | Zorro pelón | 130 | 6,09 |
| - | - | Philander vossi | Zorro cuatrojos | 26 | 1,22 |
| Total | 14 | 19 | 755 | - | - |

Fig. 2 Algunas de las especies fotocapturadas en la Reserva Sierra Zapote, Abangares, Guanacaste, Costa Rica: A) Conepatus semistriatus, B) Leopardus pardalis, C) Puma concolor, D) Eira barbara, E) Pecari tajacu, F) Cuniculus paca; G) Canis latrans, H) Herpailurus yagouaroundi
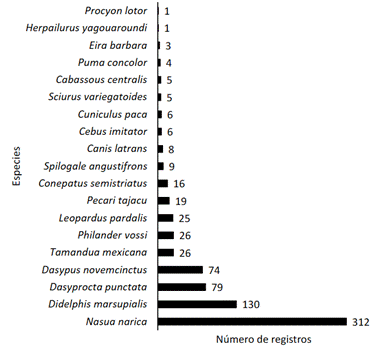
Fig. 3 Número de registros por cada una de las 19 especies de mamíferos medianos y grandes fotocapturados en la Reserva Sierra Zapote, Abangares, Guanacaste, Costa Rica
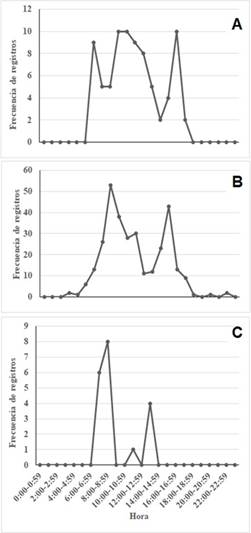
Fig. 4 Patrones de actividad diaria de las tres especies diurnas detectadas en la Reserva Sierra Zapote: A) guatusa (Dasyprocta punctata), B) pizote (Nasua narica), C) saíno (Pecari tajacu). Abangares, Guanacaste, Costa Rica
En las seis especies de hábitos nocturnos o mayormente nocturnos que detectamos en la reserva hay una cierta tendencia a más actividad cuando no hay luna en cuatro de ellas, mientras que en dos casos la tendencia a más actividad cuando hay luna (Fig. 5). Sin embargo, estas tendencias no tienen significancia estadística (X2 = 8,4, g.l. = 5, p = 0,13). Dasypus novemcinctus y Tamandua mexicana tuvieron mayor actividad cuando la intensidad lumínica fue menor (lunarfóbicas). Didelphis marsupialis y Philander vossi estuvieron más activos cuando la intensidad lumínica fue mayor (lunarfílicas). Por su lado, Leopardus pardalis y Conepatus semistriatus presentaron hábitos catamerales de actividad (Fig. 5)

Discusión
La riqueza de mamíferos medianos y grandes de 19 especies en la RSZ es comparativamente alta. En la Reserva Biológica Alberto Manuel Brenes (ReBAMB) en San Ramón, más de 100 veces más grande, se encontraron 20 especies con la misma técnica de fototrampeo (Cartín-Núñez, 2021), aunque con menos días trampa (2 135 días-trampa vs 1 281 días-trampa). No obstante, con un esfuerzo de muestreo de 1 393 días-trampa, se registraron 17 especies de mamíferos en un solo sector de esa reserva (Sánchez et al., 2019). De igual manera, el índice de diversidad H'= 1,96 para la RSZ parece mayor al de la ReBAMB de H' = 1,79 (Sánchez et al., 2019), aunque para comprobarlo se requiere un análisis estadístico comparativo apropiado (no contamos con los datos crudos de ambos trabajos para realizarlo). Además, el mono aullador (Alouatta palliata) y el puerco espín (Coendou mexicanus) son dos de las especies de mamíferos que hemos observado en la RSZ, sin embargo, no las registramos con las cámaras trampa en el periodo de nuestro estudio. Estas dos especies son arborícolas por lo que la técnica de fototrampeo no las detectó. Es posible que de igual manera otras especies no fueran detectadas por las cámaras trampa durante el estudio, por lo que la riqueza de especies de mamíferos de la reserva es mayor a la que reportamos aquí. No obstante, el valor de Chao-1 = 20 indica que quedaron pocas especies por detectar en nuestro trabajo en la RSZ. Así, la diversidad de mamíferos en la reserva es comparativamente alta (H'= 1,96), lo cual se ratifica con un valor relativamente bajo del índice de Berger-Parker = 0,41 ya que al ser este índice un valor de dominancia un aumento indica una disminución de la diversidad (Magurran, 1988). Al ser el H' un índice de heterogeneidad disminuye con la alta abundancia de algunas especies. Los elevados registros de Nasua narica, Didelphis marsupialis, Dasypus novemcinctus y Dasyprocta punctata (Cuadro 1), podrían deberse a que son especies muy comunes y de hábitos generalistas (Mora, 2000). Estas especies ocupan también hábitats abiertos por lo que se mueven tanto dentro como fuera de la reserva por lo que su detección es fácil con la técnica de fototrampeo.
De las 19 especies detectadas en la reserva con el fototrampeo destacan los cinco registros de Cabassous centralis ya que es una especie poco común y de la que hay poca información documentada a lo largo de su distribución (Marín et al., 2021). Sin embargo, desde el punto de vista de funcionalidad ecológica más destacable aún es la presencia del puma, una especie que requiere de grandes extensiones de hábitat y suficiente disponibilidad de presas para sobrevivir (Crawshaw & Quigley, 2002). Esta especie es considerada un indicador de la buena salud de un ecosistema (Cortés-Marcial & Briones-Salas, 2014; Arroyo et al., 2016). Su registro en la reserva podría ser un indicativo de la buena marcha del proceso de restauración del bosque de la reserva. Hay que tomar en cuenta que además del puma, registramos dos felinos más, el manigordo y el león breñero, que junto a seis carnívoros más (Orden Carnivora, Tabla 1), forman un gremio de nueve carnívoros de gran impacto ecológico. Sostener un gremio como este implica que las condiciones ecológicas de la reserva permiten la existencia de suficientes presas para sostenerlo. La presencia común de Nasua narica, Didelphis marsupialis, Dasypus novemcinctus y Dasyprocta punctata es un indicador de este equilibrio ecológico. Además, como complemento, aparte de ser presas, estas especies juegan papeles ecológicos de gran importancia para la dinámica y equilibrio de los ecosistemas como aireación del suelo, dispersión de semillas y depredación de presas menores como invertebrados (Mora, 2000; Lacher Jr. et al., 2019). No obstante, la presencia de perros y otros animales domésticos puede influir de forma negativa en los ecosistemas naturales, dado que pueden interactuar como posibles depredadores, competidores y como vectores de enfermedades (Vanak & Gomper 2009; Almeida, 2019). Por ejemplo, la presencia de perros domésticos puede disminuir o desplazar a los carnívoros nativos por competencia (Zapata-Ríos & Branch, 2018), pero igual podrían influir en la ausencia o baja presencia de otras especies de mamíferos.
Es notoria la ausencia del venado (Odocoileus virginianus) en las fotocapturas de la RSZ. Esta especie es común en el noroeste de Costa Rica, en particular en áreas abiertas y bordes de bosques (Rodríguez-Ramírez & Mora, 2022). Aun así, si fotocapturamos al coyote (Tabla 1), una especie de hábitos similares a los del venado (Mora, 2000). Es posible que los puntos de ubicación de las cámaras influyeran en la no obtención de fotos de venados, sin embargo, tampoco observamos esta especie durante nuestras visitas a la RSZ. El venado es una de las presas favoritas de los cazadores humanos en Costa Rica y este factor podría ser lo que está incidiendo en la RSZ. Dos especies adicionales que merecen atención particular son el saíno y el tepezcuinte ya que son especies de gran valor cinegético y son muy apreciadas y perseguidas también por los cazadores (Mora, 2000). En este sentido su presencia en la reserva es de gran significado y quizás por sus hábitos estén siendo menos afectadas por la cacería; por ejemplo, los tepezcuintes son nocturnos, en general y en la reserva en particular según nuestras observaciones.
Los patrones de actividad de los mamíferos que registramos en la reserva se ajustan a lo esperado. La mayoría de especies son de actividad nocturna y solo tres especies son de actividad diurna. El pizote es de hábitos mayormente diurnos (Reid, 2009), pero nosotros obtuvimos cuatro registros de actividad nocturna (Fig. 4). No obstante, se sabe que los pizotes machos solitarios pueden estar activos de noche a diferencia de los pizotes gregarios (Valenzuela, 2002). Por su parte, la guatusa ha sido categorizada como catemeral y diurno-crepuscular (Albanesi et al., 2016; Arroyo- Arce et al., 2016). Con nuestros registros la catalogamos como mayormente diurna, coincidente con estudios similares (Vargas-Machuca et al., 2014; Encalada, 2018). El saíno presentó picos de actividad en la mañana en la reserva (de 0600h a 0900h, Fig. 5). El tamandúa es típicamente catemeral pues en la misma localidad hay individuos activos de día mientras otros están activos de noche (Reid 2009). En la RSZ el tamandúa fue mayormente nocturna, al igual que en otras localidades (Cortés-Marcial & Briones-Salas, 2014; Arroyo-Arce et al., 2017; Guzmán, 2017; Hernández et al., 2018; Sánchez et al., 2019), aunque acá detectamos un registro a las 0900h y la especie inició su actividad a las 1600h. Por su parte, el armadillo es una especie principalmente nocturna, aunque a veces se observa activa de día (Reid, 2009). Algunos estudios han mostrado su actividad nocturna (Vargas-Machuca et al., 2014; Buenrostro-Silva et al., 2015) o mayormente nocturna (Arroyo-Arce et al., 2016; Hernández et al., 2018; Almeida, 2019). En la reserva el armadillo estuvo activo de noche (desde las 1800h), pero con un registro de actividad entre las 0800 y las 0900h.
A pesar de su actividad nocturna, la actividad de algunas especies depende de la fase lunar y son catalogadas como lunarfílicas si están activas cuando hay luna o lunarfóbicas si están activas cuando no hay luna (Albanesi et al., 2016). La actividad de seis especies analizadas en la reserva fue muy similar en las diferentes fases lunares (Fig. 5). No obstante, la tendencia fue hacia un comportamiento lunarfóbico, aunque los dos marsupiales, Phillander vossi y, en particular, Didelphis marsupialis, tuvieron más registros lunarfílicos, aunque la diferencia no fue estadísticamente significativa. El comportamiento lunarfóbico ha sido relacionado a una posible estrategia para la evasión de depredadores (Guzmán, 2017). Al haber menor intensidad lumínica de la luna podría facilitar a algunas especies pasar desapercibidas y así, evitar ser depredados (Guzmán, 2017). Este dato podría tener, además, implicaciones para la conservación. Los cazadores humanos también utilizan las fases lunares para conseguir más presas (Briones et al., 2015; Mera et al., 2017).
La RSZ es una reserva pequeña en proceso de restauración ecológica que, sin embargo, alberga una alta diversidad de MGM. Los bosques secundarios del Trópico tienden a tener mayor densidad de producción de frutos (Chinchilla, 2009). Los frutos y las semillas son fuentes alimentarias importantes para varios mamíferos. Entre los mamíferos de la reserva es particularmente notorio un gremio de carnívoros (Carnivora) de nueve especies, aunque algunas de estas como el coyote y el pizote tienen hábitos alimentarios que incluyen materia vegetal (Mora, 2000). También sobresalen cuatro especies de valor cinegético y que comúnmente la gente suele cazar, incluidas el saíno y el tepezcuinte que son dos de las especies más buscadas por los cazadores en Costa Rica. Además, otras tres especies son cazadas ocasionalmente por la gente.
La fuerte presión de caza sobre el saíno y el tepezcuinte son un problema para su conservación en general (Rodríguez-H. et al., 2002) y podría serlo para el proceso de restauración ecológica de la RSZ en particular. Aunque no tenemos datos sobre cacería en la RSZ sabemos que si la hay en la finca Cerritos del SINAC que está a solo 2km aproximadamente de la RSZ. El saíno y el tepezcuinte son presas importantes para el puma, pero sobre todo para el jaguar (Panthera onca) que, aunque no se ha detectado en la reserva se ha recuperado poblacionalmente en varios sitios del país (Montalvo et al., 2015). La eliminación de tepezcuintes de un sitio podría producir efectos asociados como la disminución de la diversidad y supervivencia de especies vegetales ya que son dispersores claves (Martinez-Ceceñas et al., 2018). También el zorro pelón, el armadillo y la ardilla común son presas de los carnívoros presentes en la RSZ, por lo que su eliminación por cacería humana significa la disminución de presas disponibles. La falta de registros del venado en la RSZ durante nuestro estudio podría ser un indicativo de cacería, aunque hay argumentos ecológicos que podrían explicar su ausencia.
Las especies cinegéticas son parte de la biomasa total de la reserva por lo que son parte de las redes alimentarias cuya complejidad refleja el estado de conservación de un ecosistema. El remover esta biomasa sin un plan de manejo adecuado atenta contra el funcionamiento del ecosistema, en particular de áreas que están en proceso de restauración ecológica. Una baja riqueza de especies en paisajes fragmentados puede modificar más que solo la estructura del bosque y la diversidad de especies (Chinchilla, 2009). Varios mamíferos de valor cinegético son frugívoros y cumplen funciones ecológicas muy importante en la regeneración y restauración de los bosques (Wang et al., 2007; Wright et al., 2000). Una meta que podría plantearse para la RSZ es conservar a largo plazo una alta abundancia de presas para los carnívoros y que a la vez cumplen funciones ecológicas varias como la dispersión de semillas. Estas presas incluyen a las especies cinegéticas mencionadas, ya sea poco utilizadas como la guatusa o muy perseguidas como el tepezcuinte. La abundancia de estos mamíferos determinará la permanencia de los carnívoros, en especial el puma y el manigordo que están en la parte alta de la pirámide alimentaria. Esto se podría lograr, al menos en parte, con vigilancia y control en la RSZ y un programa de educación ambiental en la zona de influencia de la reserva. Cuando los recursos alimentarios son abundantes, es posible que haya mayor traslape entre especies, inclusive una interacción mayor entre los depredadores y las presas (Silva-Pereira et al., 2011). En un área en restauración estas interacciones son clave, al igual que lo son las interacciones planta-animal tales como la herbivoría, la frugivoría, la polinización, la dispersión de semillas, el pisoteo de plántulas y la depredación de semillas entre varias otras (Chinchilla, 2009). Depredadores y presas proveerán a la reserva de funciones y servicios ecosistémicos indispensables en el proceso de restauración ecológica.
Ética, discusión, confilcto de intereses y declaración en financiamiento
Declaramos haber cumplido con todos los requisitos éticos y legales pertinentes, tanto durante el estudio como en la preparación de este documento; que no hay conflictos de interés de ningún tipo, y que todas las fuentes financieras se detallan plena y claramente en la sección de agradecimientos. Asimismo, estamos de acuerdo con la versión editada final de esta publicación. El respectivo documento legal firmado se encuentra en los archivos de la revista.
La declaración de contribución de cada autor es la siguiente: P.M. y A.V.C.: Diseño del estudio, recolección y análisis de datos. L.I.L. y J.M.M.: Preparación de figuras y análisis de datos. P.M. y J.M.M.: preparación del manuscrito. Todos los coautores: edición y aprobación final del manuscrito.

