











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Las fluctuaciones climáticas alteran el volumen de agua y consecuentemente la disponibilidad de hábitats, tanto bentónicos como ribereños; lo que, a su vez, afecta la presencia y dinámica poblacional de distintos grupos de macroinvertebrados acuáticos, entre ellos los quironómidos (Rivera-Usme et al., 2013). En zonas tropicales, las diferencias en la composición de las comunidades, entre épocas climáticas contrastantes, están relacionadas con las estrategias de vida de los organismos, que definen la presencia o ausencia de algunos taxones en sequía, transición, o alta precipitación (Serna-Macias et al., 2023). Los quironómidos (Diptera: Chironomidae) son invertebrados bentónicos altamente diversos, abundantes y claves en la dinámica de nutrientes de los ecosistemas dulciacuícolas del mundo (Ferrington, 2008; Pero et al., 2023; Silva et al., 2018). Algunos géneros son considerados indicadores de condiciones ambientales particulares; se presentan y distribuyen en zonas con niveles variables de perturbación, responden a gradientes latitudinales y altitudinales en ambientes extremos y con requerimientos de hábitat altamente específicos (Oviedo-Machado & Reinoso-Flórez, 2018; Villamarín et al., 2012). En su fase larval colonizan diferentes sustratos presentes en la red de drenaje, y son comunes en sistemas acuáticos de alta montaña del neotrópico (Prat et al., 2014; Rieradevall et al., 2007; Villamarín et al., 2021), con aparente dominancia en lenticos (lagos, lagunas, humedales, embalses, y pantanos) versus lóticos (ríos o arroyos, Andersen et al., 2017).
La Familia Chironomidae es amplia y diversa, con cerca de 400 géneros (> 6 200 spp.) y con una proyección de entre 10 000 a 15 000 especies en el mundo (Silva et al., 2018); sin embargo, el conocimiento taxonómico y otros rasgos de historia de vida son aún desconocidos, particularmente en el neotrópico (Molineri et al., 2020; Prat & Castro-López, 2023; Silva et al., 2018). Su distribución y abundancia en arroyos y ríos es altamente influenciada por la altitud, temperatura, caudal, disponibilidad de alimento, tipo de sustrato (Armitage et al., 1995; Lindegaard & Brodersen, 1995; Rae, 2004) o aspectos biogeográficos (Scheibler et al., 2008). En ecosistemas altoandinos, se ha encontrado que algunos arroyos localizados a mayor altitud comparten taxones tanto eurioicos como estenoicos (Tejerina & Molineri, 2007).
A pesar de algunos avances realizados tanto en estudios taxonómicos como ecológicos sobre quironómidos, en varios sistemas dulceacuícolas altoandinos de Colombia (Caleño-Ruiz et al., 2018; Hoyos-Jaramillo & Gomes-Dias, 2020; Ospina-Torres et al., 1999; Ruiz-Moreno et al., 2000a; Ruiz-Moreno et al., 2000b; Tamaris-Turizo et al., 2023), aún persisten vacíos de conocimiento a nivel de género o especie (Oviedo-Machado & Reinoso-Flórez, 2018). Este panorama es contrastante con los avances logrados en las últimas décadas en sistemas acuáticos similares de la región Neotropical (e.g., Acosta & Prat, 2010; Acosta & Prat, 2018; Prat et al., 2011; Prat et al., 2014; Prat & Castro-López, 2023; Villamarín et al., 2021). En el departamento del Tolima, zona central de Colombia, el análisis de quironómidos tradicionalmente ocurre como parte del componente biológico, en estudios con macroinvertebrados acuáticos (Forero-Céspedes et al., 2013). No obstante, algunas de las investigaciones recientes (Oviedo-Machado & Reinoso-Flórez, 2018; Rojas-Céspedes et al., 2018; Rojas-Sandino et al., 2018), se han realizado con énfasis en el Orden Diptera y la Familia Chironomidae en zonas bajas del departamento (cuencas con bosque seco tropical).
Dada la importancia de los quironómidos en las redes tróficas, al igual que el potencial bioindicador de algunos géneros frente a la contaminación o cambios en su entorno, y su dinámica espacial y temporal (Molineri et al., 2020; Nicacio & Juen, 2015; Rodríguez-Rodríguez et al., 2021; Tamaris-Turizo et al., 2023). En el presente estudio se evaluó la dinámica espaciotemporal de la Familia Chironomidae a nivel de género, en tres quebradas altoandinas (Las Perlas, La Plata, y Cay), tributarias del Río Combeima (zona central de Colombia), las cuales reflejan gradientes altitudinales y de perturbación (Rojas-Céspedes et al., 2022; Váquiro-Capera et al., 2020). Los objetivos fueron: (i) identificar los géneros de la Familia Chironomidae presentes en cuencas altoandinas del departamento del Tolima y su asociación con aspectos ecológicos, (ii) determinar la riqueza, abundancia, diversidad y distribución espacial, temporal y altitudinal de los taxones registrados. El presente estudio amplia la información sobre los quironómidos en el país; además, constituye el primer estudio en sistemas acuáticos altoandinos en el departamento del Tolima.
Materiales y métodos
Área de estudio: La investigación se realizó en tres zonas de muestreo (parte alta [1], media [2], y baja [3]) de las quebradas Las Perlas (PE), La Plata (PL), y Cay (CA), tributarias de la cuenca del Río Combeima (área de 27 421 ha), que a su vez desemboca la cuenca del Río Coello, en el departamento del Tolima, zona central de Colombia. El Río Combeima se origina en el Nevado del Tolima a 5 225 m.s.n.m. Recibe aguas de diez quebradas principales, que representan el 35 % del área total de la cuenca. A su vez, presta múltiples servicios ambientales a nivel local y regional, ya que suministra el 80 % del agua para el acueducto de la ciudad de Ibagué, beneficia a más de 542 046 habitantes, y genera una importante oferta hídrica para el desarrollo de diferentes actividades agropecuarias de la región (Consejo Nacional de Política Económica y Social [CONPES], 2009).
Las tres microcuencas seleccionadas (Fig. 1), se ubican dentro de las zonas de vida Premontano, Montano bajo y Montano (1 000-3 000 m.s.n.m.) con una temperatura que oscila entre 6-24 °C (clima: templado cálido y templado frío). Se asocian con la vertiente oriental de la Cordillera Central (04º19’30’’-04º39’57’’ N & 75º10’11’’-75º23’23’’ W). Su rango altitudinal incluye desde los 5 225 m en su nacimiento en el nevado del Tolima hasta 648 m.s.n.m. en su desembocadura en el Río Coello (Váquiro-Capera et al., 2020). La zona representa uno de los lugares de mayor afluencia turística del municipio de Ibagué (Leonel & Páez, 2000). Las microcuencas, en general, presentan efectos de degradación antropogénica por ganadería, agricultura y urbanización (Rojas-Céspedes et al., 2023). La precipitación media anual es de 1 816 mm3, con dos épocas de sequía y dos de lluvia (distribución bimodal): las primeras de diciembre a marzo y de julio a agosto, y las segundas de abril a junio y de septiembre a noviembre, con una temperatura media anual de 17 ºC (Corporación Autónoma Regional del Tolima [CORTOLIMA], 2006). Los muestreos (M1-M6), se realizaron en agosto (M1, bajas lluvias), octubre (M2, altas lluvias) y noviembre (M3, transición bajas lluvias) de 2015 y febrero (M4, transición bajas lluvias), abril (M5, altas lluvias) y mayo (M6, transición bajas lluvias) de 2016.
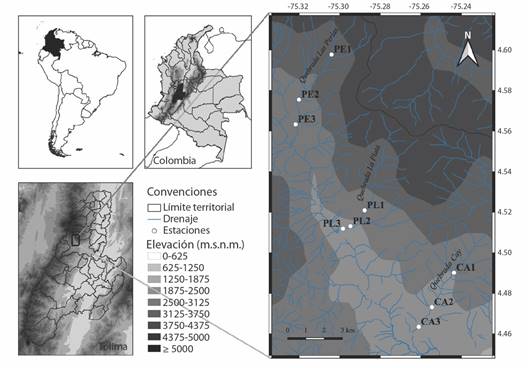
Fig. 1 Ubicación de las tres microcuencas tributarias de la cuenca del Río Combeima (Tolima, Colombia). En cada quebrada se indican los puntos de muestreo en la parte alta (1), media (2), y baja (3), respectivamente. / Fig. 1. Location of the three tributary microbasins in the Combeima River Basin (Tolima, Colombia). In each stream, the sampling points are indicated in the upper (1), middle (2), and lower (3) parts.
Trabajo de campo: Los quironómidos evaluados se obtuvieron de recolecciones en un macroproyecto con macroinvertebrados del Grupo de Investigación en Zoología (GIZ), Facultad de Ciencias, Universidad del Tolima (Ibagué, Colombia). La recolecta se realizó siguiendo los protocolos y técnicas propuestas por Wantzen y Rueda (2009), a través de métodos cuantitativos como red Surber (0.09 m2 de área y poro de malla de 250 μm). Los organismos se almacenaron en frascos plásticos debidamente marcados con la información de campo, y se preservaron en alcohol al 70 %. A nivel temporal, se utilizó la información de los registros pluviométricos históricos (~ 10 años) suministrados por el Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia (IDEAM), con referencia a los periodos de altas y bajas precipitaciones.
Trabajo de laboratorio: Se realizó la separación e identificación a género, mediante micro-preparados (montaje y fijación de placas con piezas bucales, antenas y parápodos posteriores); para esta actividad, se utilizó el 10 % de los individuos en cada muestra con base en los registros espaciales y temporales. Se siguieron las claves de Roldán (1996), Ospina-Torres et al. (1999), Ruiz-Moreno et al. (2000a), Ruiz-Moreno et al. (2000b), Epler (2001), Courtney y Merritt (2008), Ferrington (2008), Merritt et al. (2008), Domínguez y Fernández (2009), Prat et al. (2011), Trivinho-Strixino (2011) y Andersen et al. (2013). Los especímenes restantes fueron preservados en tubos Vacutainer® con alcohol al 70 % e ingresados a la Colección Zoológica de la Universidad del Tolima (CZUT, Ibagué, Colombia). Se registró in situ oxígeno disuelto, porcentaje de saturación de oxígeno, conductividad, caudal y temperatura del agua. Paralelamente se tomaron muestras de agua para análisis ex situ de pH, turbiedad, DQO, alcalinidad, cloruros, dureza, sólidos totales, sólidos suspendidos, fosfatos, nitratos, y coliformes totales y fecales, en el laboratorio de análisis químico (LASEREX) de la Universidad del Tolima (Ibagué, Colombia; ver Rojas-Céspedes et al., 2023).
Análisis de datos: Se organizó una matriz de datos por quebrada, con información de los tres puntos (parte alta, media, y baja), y frecuencia de muestreo (M1-M6). Con los datos obtenidos durante todo el estudio, se calculó la abundancia relativa por género, ordenación del ensamblaje, diversidad con números de Hill, y se estimó una curva de acumulación de taxones (= géneros). Para el análisis del ensamblaje de quironómidos, se calculó la matriz de similitud de Bray-Curtis y se graficó en un espacio de ordenación reducido mediante escalamiento multidimensional no métrico (NMDS; función metaMDS) con el paquete “vegan” (Oksanen et al., 2020). Asimismo, se realizó un PERMANOVA para evaluar diferencias en los patrones de distribución espacial y temporal de los géneros: (i) entre quebradas (efecto fijo con tres niveles; Las Perlas, La Plata, y Cay), (ii) en el tiempo (efecto aleatorio con seis niveles; M1-M6). Este procedimiento se realizó con la función Adonis (999 permutaciones; Oksanen et al., 2020), y se complementó con pruebas post-hoc por pares (Martínez, 2017), en RStudio versión 3.2.5 (R Development Core Team, 2016). Para identificar los agrupamientos y detectar diferencias en la composición taxonómica de los géneros con mayor contribución entre sitios (quebradas/puntos de muestreo) y temporalidad, se utilizó el procedimiento SIMPER (Clarke & Warwick, 2001).
La diversidad se calculó con números de Hill o número efectivo de taxones con base en la riqueza y abundancia relativa de los géneros identificados (Chao et al., 2014; Jost, 2006). Se realizó un análisis de rarefacción con 100 aleatorizaciones y extrapolación al doble del número de individuos para los órdenes q0, q1 y q2, con el fin de identificar diferencias en la diversidad entre quebradas a nivel temporal (Chao et al., 2014), mediante RStudio versión 3.2.5 y las librerías Vegan e iNEXT (Chao et al., 2014; R Development Core Team, 2016). Adicionalmente, se realizó una evaluación de la riqueza taxonómica a través de una curva de acumulación entre los géneros observados y las unidades de muestreo. La representatividad del muestreo fue evaluada con los géneros esperados, mediante los estimadores Chao 1, ACE y Cole calculados con EstimateS 9.1 (Colwell, 2024).
Resultados
Composición general y aspectos ecológicos: Se registraron 15 594 larvas de la Familia Chironomidae, pertenecientes a tres subfamilias (Chironominae, Orthocladiinae, Tanypodinae) y 20 géneros (Tabla 1). La subfamilia más abundante fue Orthocladiinae (13 697 larvas, 87.84 %) con los géneros Cardiocladius, Corynoneura, Cricotopus, Onconeura, Parametriocnemus, Rheocricotopus y Thienemanniella, seguida de Chironominae (1 367 individuos, 8.77 %) con Chironomus, Cladotanytarsus, Cryptochironomus, Dicrotendipes, Endotribelos, Paratanytarsus, Polypedilum, Riethia y Saetheria. Tanypodinae registró la menor abundancia relativa acumulada (530 organismos, 3.40 %). El género más abundante en Orthocladiinae fue Cricotopus (66.19 %), seguido de Cardiocladius (6.73 %), Parametriocnemus (5.81 %) y Onconeura (5.65 %), que se corresponden con los valores más altos reportados durante el estudio (> 5.5 %). En Chironominae fue Polypedilum (5.18 %) seguido de Paratanytarsus (2.58 %), y en Tanypodinae fue Pentaneura (2.91 % de todo el estudio, Tabla 1).
Tabla 1 Composición y abundancia total de larvas de la Familia Chironomidae en las quebradas Las Perlas, La Plata y Cay en el Río Combeima (Tolima, Colombia), recolectadas en nueve puntos de muestreo durante agosto (M1), octubre (M2), noviembre 2015 (M3) y, febrero (M4), abril (M5) y mayo 2016 (M6). / Table 1. Composition and total abundance of larvae of the Family Chironomidae in the Las Perlas, La Plata and Cay streams in the Combeima River (Tolima, Colombia), collected at nine sampling points during August (M1), October (M2), and November 2015 (M3), and February (M4), April (M5), and May 2016 (M6).
| Subfamilia | Género | Las Perlas (PE) | La Plata (PL) | Cay (CA) | Total | ||||||||||||||||||
| M1 | M2 | M3 | M4 | M5 | M6 | M1 | M2 | M3 | M4 | M5 | M6 | M1 | M2 | M3 | M4 | M5 | M6 | ||||||
| Chironominae | Cryptochironomus | 1 | 1 | 1544 | |||||||||||||||||||
| Cladotanytarsus | 1 | 3 | 2 | 5 | 11 | ||||||||||||||||||
| Dicrotendipes | 1 | 16 | 17 | ||||||||||||||||||||
| Chironomus | 1 | 13 | 7 | 4 | 25 | ||||||||||||||||||
| Riethia | 1 | 3 | 8 | 11 | 5 | 4 | 32 | ||||||||||||||||
| Saetheria | 1 | 13 | 8 | 3 | 7 | 32 | |||||||||||||||||
| Endotribelos | 5 | 2 | 6 | 24 | 3 | 40 | |||||||||||||||||
| Paratanytarsus | 46 | 17 | 23 | 44 | 6 | 2 | 45 | 19 | 171 | 122 | 28 | 17 | 540 | ||||||||||
| Polypedilum | 14 | 14 | 7 | 3 | 1 | 150 | 57 | 157 | 283 | 128 | 32 | 846 | |||||||||||
| Orthocladiinae | Thienemanniella | 2 | 1 | 2 | 5 | 13662 | |||||||||||||||||
| Corynoneura | 10 | 2 | 11 | 110 | 37 | 40 | 3 | 5 | 218 | ||||||||||||||
| Rheocricotopus | 4 | 1 | 3 | 9 | 2 | 173 | 3 | 25 | 2 | 58 | 1 | 281 | |||||||||||
| Onconeura | 2 | 1 | 14 | 24 | 63 | 11 | 27 | 5 | 5 | 12 | 2 | 5 | 53 | 23 | 413 | 65 | 89 | 67 | 881 | ||||
| Parametriocnemus | 1 | 3 | 61 | 21 | 2 | 2 | 282 | 8 | 22 | 11 | 1 | 180 | 4 | 90 | 141 | 8 | 69 | 906 | |||||
| Cardiocladius | 5 | 3 | 4 | 105 | 1 | 45 | 59 | 5 | 19 | 107 | 40 | 468 | 100 | 47 | 42 | 1050 | |||||||
| Cricotopus | 15 | 45 | 126 | 50 | 48 | 93 | 544 | 129 | 425 | 1950 | 36 | 807 | 824 | 206 | 1002 | 922 | 1902 | 1197 | 10321 | ||||
| Tanypodinae | Thienemannimyia | 2 | 1 | 1 | 4 | 388 | |||||||||||||||||
| Alotanypus | 6 | 8 | 14 | ||||||||||||||||||||
| Larsia | 6 | 4 | 5 | 17 | 18 | 4 | 54 | ||||||||||||||||
| Pentaneura | 1 | 13 | 32 | 2 | 1 | 86 | 24 | 44 | 88 | 14 | 11 | 316 | |||||||||||
| TOTAL | 22 | 51 | 238 | 145 | 117 | 124 | 1128 | 174 | 533 | 2116 | 50 | 834 | 1666 | 389 | 2440 | 1782 | 2319 | 1466 | |||||
| 697 | 4835 | 10062 | |||||||||||||||||||||
| 15594 | |||||||||||||||||||||||
Representatividad del muestreo: Los estimadores de riqueza incluyeron los 20 géneros observados (Sobs) entre las tres subfamilias, en los nueve puntos evaluados, lo que correspondió al 100 % del valor esperado para cada estimador (S Mean, Chao 1, ACE, Cole Rarefacción) para la quebrada Cay, mientras que, para Las Perlas y La Plata, se encontró que los géneros identificados corresponden al 78.55 y 97.09 % de los taxones esperados (Fig. 2).
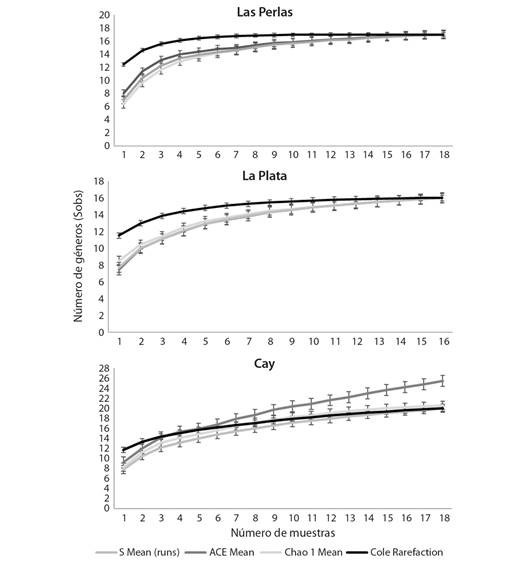
Fig. 2 Estimación de la riqueza de la Familia Chironomidae en nueve puntos de muestreo durante un ciclo hidrológico en tres quebradas de la cuenca del Río Combeima (Tolima, Colombia). / Fig. 2. Estimation of the richness of the Chironomidae Family at nine sampling points during a hydrological cycle in three streams of the Combeima River Basin (Tolima, Colombia).
Números de Hill: A partir de la extrapolación del número de taxones (q0) por muestreos se observó que la mayor riqueza taxonómica ocurrió en la quebrada Cay (20 géneros), seguida de La Plata y Las Perlas (Fig. 3). Durante los muestreos M4 y M6 se registró la mayor riqueza en Cay con 16 géneros, mientras que en La Plata ocurrió en M3, y en Las Perlas en M3 y M4. En relación con los géneros comunes (q1) y taxones dominantes la respuesta fue similar a la riqueza, donde la quebrada Cay obtuvo los mayores registros y el valor más bajo ocurrió en la quebrada Las Perlas, con variaciones temporales (Fig. 3). A partir del número de géneros típicos o comunes (q1) se evidenció que las subfamilias Chironominae y Tanypodinae mostraron abundancias similares con Chironomus, Endotribelos, Riethia, Saetheria, y Larsia. Respecto al número de géneros dominantes (q2), se observó que Cricotopus (10 321), Cardiocladius (1 050), Parametriocnemus (906), Onconeura (881) y Polypedilum (846) registraron los valores más altos.
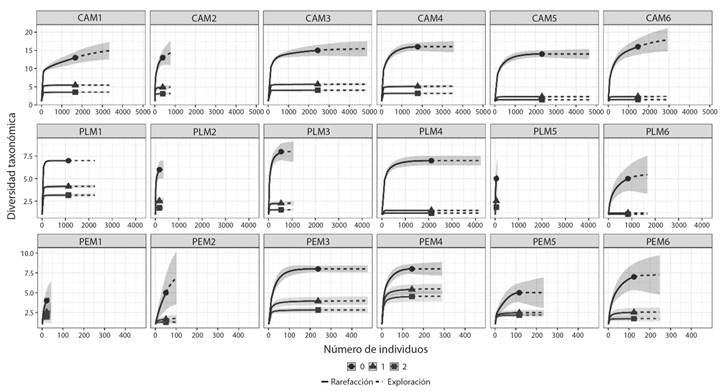
Fig. 3 Rarefacción/extrapolación del número efectivo de especies del orden q (q0, q1 y q2) en seis muestreos (M1-M6) realizados en la cuenca del Río Combeima (Tolima, Colombia). Quebradas: Las Perlas (PE), La Plata (PL), y Cay (CA). / Fig. 3. Rarefaction/extrapolation curves of the effective number of species of the order q (q0, q1, and q2) in six samplings (M1-M6) in the Combeima River Basin (Tolima, Colombia). Streams: Las Perlas (PE), La Plata (PL), and Cay (CA).
Ensamblaje de los géneros registrados: La ordenación NMDS mostró a nivel espacial, solapamiento parcial de las quebradas Las Perlas y La Plata, y una clara separación de la quebrada Cay (NMDS estrés = 0.12). La separación de las quebradas evaluadas se asocia principalmente con las bajas abundancias de Chironominae y Tanypodinae registradas en Las Perlas y La Plata (Fig. 4). Sin embargo, no se encontraron diferencias significativas en la abundancia de quironómidos de las tres quebradas evaluadas (PERMANOVA, Pseudo-F = 1.30; p > 0.05). Los géneros Cryptochironomus, Endotribelos, Thienemanniella, Alotanypus, y Thienemannimyia presentaron la menor distribución altitudinal, mientras que la mayoría se registraron entre 1 419-2 586 m.s.n.m. (Fig. 5).
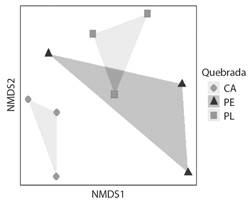
Fig. 4 Diagrama de ordenación NMDS con base en las abundancias en los géneros de la Familia Chironomidae registrados en seis muestreos en las quebradas Las Perlas (PE), La Plata (PL), y Cay (CA). / Fig. 4. Ordering diagram produced by NMDS analysis based on the abundance of the genera of the Chironomidae Family recorded in six samplings in the Las Perlas (PE), La Plata (PL), and Cay (CA) streams.

Discusión
La composición de los géneros registrados coincide con lo reportado por otros autores en quebradas altoandinas similares, donde los quironómidos presentaron alta variabilidad, correlacionada principalmente con la temperatura. La subfamilia Orthocladiinae presentó la mayor diversidad y abundancia en los puntos de muestreo con bajas temperaturas del agua (partes altas de cada quebrada; Rojas-Céspedes et al., 2023), característicos de ambientes lóticos, donde el género que refleja mayor dominancia es comúnmente Cricotopus (Butakka et al., 2014a; Butakka et al., 2014b; Castella et al., 2001; Lods-Crozet et al., 2001a; Lods-Crozet et al., 2001b; Maiolini & Lencioni, 2001; Tejerina & Molineri, 2007; Prat et al., 2014).
Los 20 géneros registrados están distribuidos en tres de las 11 subfamilias existentes en Colombia (Hoyos-Jaramillo & Gomes-Dias, 2020). Orthocladiinae fue el taxón dominante en todos los puntos de muestreo, tanto en los que evidenciaron mayor impacto por intervención antropogénica, como en los más conservados. La alta abundancia de esta subfamilia (10 veces > Chironominae; ≈ 40 veces > Tanypodinae) se asocia con su amplia colonización y tolerancia en todos los tipos de hábitats acuáticos (lóticos, lenticos e incluso salobres). Orthocladiinae se considera dominante a menor temperatura, y ampliamente adaptada a ambientes lóticos de aguas frías con predominio en quebradas andinas (Paggi, 2009; Pero et al., 2023; Scheibler et al., 2008). Lo anterior, coincide con lo reportado por otros autores en ríos altoandinos (Acosta & Prat, 2010; Scheibler et al., 2008; Villamarín, 2012), donde esta subfamilia muestra mayor abundancia y frecuencia relativa, asociadas con la estenotermia (Lencioni et al., 2015), particularmente en zonas frías, correspondiente a las cabeceras y tramos intermedios de los arroyos (Paggi, 2009). De esta subfamilia, Cricotopus registró la mayor abundancia en PL3, zona cercana al acueducto comunitario procedente de la quebrada Las Perlas. No obstante, en cercanías a este punto de muestreo, se presenta alta intervención antropogénica por agricultura, ganadería, turismo, urbanización y presencia de coliformes (251 652.5 NMP/100mL; Rojas-Céspedes et al., 2023). Los valores anteriores son coincidentes con los estudios reportados por García y Añón (2007) y Tejerina y Molineri (2007) en arroyos argentinos, quienes resaltan que Cricotopus es el género más abundante y común de Orthocladiinae, al ser cosmopolita y de fácil adaptación a diversas variables ambientales. Además, posee densidades elevadas en la mayoría de los estudios bentónicos (Zanotto-Arpellino et al., 2015).
Chironominae fue la segunda subfamilia más abundante, y la de mayor riqueza taxonómica, con géneros que constituyen nuevos registros para zonas altas del departamento del Tolima. Esta subfamilia comprende aproximadamente el 50 % de las especies de quironómidos que han sido descritas en la región Neotropical, que es más del doble de la proporción de cualquier otra subfamilia y se encuentran en una amplia variedad de hábitats acuáticos y semiacuáticos, aunque son relativamente más abundantes en aguas bajas estancadas y de flujo lento, que en sitios lóticos en áreas montañosas (Andersen et al., 2017). No obstante, en las quebradas evaluadas registró una baja abundancia (11.3 % con respecto a Orthocladiinae). En el presente estudio, Polypedilum fue el género más abundante, con el mayor valor en CA1 que corresponde a la zona ubicada en la parte alta de la quebrada Cay, con presencia de zonas de rápidos resultantes de un terreno encañonado, rodeado por relictos de vegetación ribereña, con incidencia de algunos asentamientos humanos (Rojas-Céspedes et al., 2023). No obstante, la distribución de este género no estuvo relacionada con un patrón determinado o alguna característica particular entre las zonas evaluadas, ya que regularmente es de amplia distribución, común en todo tipo de hábitats acuáticos, tanto en sistemas lénticos como lóticos (Epler, 2001; Rojas-Céspedes et al., 2018; Trivinho-Strixino, 2011). En Tanypodinae, Pentaneura fue el más representativo a mayor altitud de la quebrada Las Perlas (PL1). Allí se encuentra una cascada que descarga gran cantidad de agua, presenta cobertura boscosa densa, a pesar de un alto grado de intervención antropogénica por agricultura y turismo (Rojas-Céspedes et al., 2023). Sin embargo, este resultado coincide con lo reportado por Prat et al. (2011), quienes indican que es un género ampliamente distribuido y es propio de los ríos altoandinos, con mayores abundancias en las partes más altas.
A nivel temporal, la mayor abundancia registrada en las tres quebradas en M3 y M4 (transición a bajas lluvias), puede estar asociada con la baja alcalinidad y turbiedad, alta conductividad eléctrica y concentración de sólidos totales, y bajas velocidades (Rojas-Céspedes et al., 2023). Por otra parte, la altitud es un factor que influye en la composición y distribución de las larvas de Chironomidae en los arroyos y ríos (Acosta, 2009; Jacobsen et al., 2003; Pero et al., 2023; Villamarín, 2012). En el presente estudio, Orthocladiinae y Chironominae, reflejaron una tendencia a incrementar su abundancia a mayor altitud, mientras que Tanypodinae reflejó un patrón inverso, lo cual puede indicar que el gradiente altitudinal influye en la composición, distribución y diversidad de la Familia Chironomidae.
La diversidad con números de Hill resaltó la importancia de la quebrada Cay, que comparativamente presentó el menor caudal promedio durante el muestreo (13.71 m³/s versus 62.85 Las Perlas, 67.83 La Plata), y registró la mayor riqueza y dominancia tanto a nivel espacial como temporal, asociadas con Cricotopus y Cardiocladius. La variación de la diversidad (q0, q1, y q2; Fig. 3), también se puede relacionar con los cambios de caudal registrados en las quebradas en los distintos periodos climáticos. Por lo tanto, la composición y dinámica de la comunidad de quironómidos estuvo influenciada principalmente por las características locales de cada quebrada y punto de muestreo (Rojas-Céspedes et al., 2023), lo cual coincide con estudios realizados en zonas de alta montaña como los de Medina et al. (2008), Villamarín et al. (2021) y Rojas-Céspedes et al. (2022).
Las densidades de quironómidos pueden predominar, tanto en épocas de alta como de baja precipitación, gracias a los estrechos rangos de tolerancia a diferentes variables ambientales (López & Talero, 2015; Scheibler et al., 2008; Zanotto-Arpellino et al., 2015). Los puntos de muestreo seleccionados corresponden a lugares con intervención antropogénica diferencial (Las Perlas < La Plata y Cay; y en cada una, parte alta < media < baja), presencia de vegetación ribereña abundante y diferentes hábitats y sustratos en el cauce (Rojas-Céspedes et al., 2022), condiciones que favorecen el establecimiento y abundancia de estos organismos (Oviedo-Machado & Reinoso-Flórez, 2018; Rojas-Céspedes et al., 2018; Rojas-Céspedes et al., 2023).
Para este estudio la curva de acumulación indicó una buena representatividad y, por lo tanto, un esfuerzo muestreo adecuado según los estimadores evaluados. Se alcanzó una representatividad entre 78 y 100 %; sin embargo, es posible el registro de nuevos géneros (y potenciales especies), dado que la curva de acumulación se acerca parcialmente a la asíntota, principalmente en la quebrada Cay. Los taxones de Chironomidae compartidos entre las quebradas evaluadas, se corresponden con géneros comunes en zonas altoandinas, de manera similar a lo encontrado por otros autores (Lindegaard & Brodersen, 1995; Pero et al., 2023; Tejerina & Molineri, 2007; Thienemann, 1954). Las quebradas Las Perlas y La Plata, compartieron bajas abundancias de Dicrotendipes y Thienemannimyia. No obstante, se reporta que éstos son géneros de amplia distribución altitudinal, desde ecosistemas de zonas bajas hasta de alta montaña tropical (Butakka et al., 2014a; Butakka et al., 2014b; Prat et al., 2014; Caleño-Ruiz et al., 2018; Catalan et al., 2006; Henriques-Oliveira et al., 2003; Sanseverino & Nessimian, 2008; Silva et al., 2008).
Se amplía el área de distribución regional y altitudinal de la Familia Chironomidae, al registrarse por primera vez, 20 géneros en estado larval de las subfamilias Orthocladiinae, Chironominae, y Tanypodinae para quebradas de alta montaña del departamento del Tolima (zona central de Colombia). La proporción de estas subfamilias fue altamente variable entre puntos y periodos de muestreo, como ocurre en otros ecosistemas acuáticos altoandinos, donde Orthocladiinae puede dominar ampliamente frente a Chironominae y Tanypodinae. Cricotopus, Cardiocladius, Onconeura, Parametriocnemus, Paratanytarsus, Pentaneura, Polypedilum, Rheocricotopus y Riethia fueron los géneros más comunes tanto a nivel espacial como temporal, con altas densidades relativas con respecto a Alotanypus, Chironomus, Cladotanytarsus, Corynoneura, Cryptochironomus, Dicrotendipes, Endotribelos, Larsia, Saetheria y Thienemannimyia, que presentaron bajas densidades y distribución restringida, debido posiblemente a los atributos funcionales de cada género, tales como la morfología, comportamiento y preferencias de microhábitat. El presente estudió permitió evidenciar que las condiciones de calidad de las quebradas pueden evaluarse con organismos de la Familia Chironomidae; sin embargo, es necesario una mejor comprensión de las preferencias ecológicas de los taxones registrados. Por lo tanto, se debe continuar con la profundización del conocimiento taxonómico (idealmente a nivel de especie), biológico y ecológico que permita adecuar e implementar estrategias de manejo, conservación y restauración de los ecosistemas lóticos altoandinos de Colombia y los demás países del Neotrópico.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

