











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkLa diversidad es una característica susceptible de ser medida en los ecosistemas marinos, como la productividad, densidad y biomasa (Alcolado, 1998). Los índices de diversidad reflejan la complejidad y estructura del sistema en estudio (Pecenko, 1982), aunque otros autores reconocen que éste es un tema principal a tratar en la ecología (Magurran, 2004; Rodríguez-Zaragoza et al., 2011). Así como el concepto ha generado discusión semántica, también es polémico el origen de los factores que determinan la diversidad (Alcolado, 1998). Se recomienda utilizarlo como un concepto genérico que englobe los conceptos específicos de riqueza, heterogeneidad, equidad, entre otros índices que se complementen entre sí, según los objetivos y diseño de la investigación (Peet, 1974; Rodríguez-Zaragoza et al., 2011).
Actualmente, muchos grupos de gestión y conservación de los ecosistemas utilizan el declive de la diversidad como medio para justificar las decisiones en políticas de manejo (Heller & Zavaleta, 2009; Rands et al., 2010; Wong, Jiang, Kinzig, Lee, & Ouyang, 2015). Las naciones caribeñas reconocen cada vez más la riqueza natural de los arrecifes coralinos (Hughes et al., 2003), donde los ecosistemas marinos con mayor diversidad tendrán mejor funcionamiento y resiliencia.
La implementación de áreas marinas protegidas (AMP) se ha convertido en una herramienta efectiva para la conservación de la biodiversidad en los arrecifes coralinos (Gell & Roberts, 2003; Russ & Alcala, 2004). Se ha demostrado la efectividad de las AMPs respecto al incremento de la riqueza de especies de peces en arrecifes coralinos (Gell & Roberts, 2002; Willis, 2003; Palumbi, 2004), también como en el aumento de la resiliencia en comunidades arrecifales (Mellin, MacNeil, Cheal, Emslie, & Caley, 2016); sin embargo, el sistema global actual de AMP es insuficiente para proteger la diversidad total de corales y peces (Mouillot et al., 2016).
Dentro de las causas de la pérdida de diversidadFiguran la destrucción del hábitat, la sobreexplotación, la contaminación, el cambio climático, y las especies invasoras (Santos & Tellería, 2006; Gutiérrez, 2006; Capote López, Mitrani Arenal, & Suárez, 2012). Las especies invasoras son aquellas que se establecen en ecosistemas naturales o seminaturales, y constituyen un agente de cambio y de amenaza a la diversidad biológica nativa (Mendoza, Luna, & Aguilera, 2015). Dentro de los principales impactos ecológicos que provocan destacan la degradación de los hábitats, el desequilibrio ecosistémico, el desplazamiento y la extinción de la flora y fauna nativa, la alteración de la estructura trófica, la facilitación de invasiones subsiguientes y la transmisión de enfermedades (Gutiérrez, 2006).
El pez león (Pterois volitans/miles) es reconocido como la mayor invasión marina en el Atlántico y el Caribe (Hixon, Green, Albins, Akins, & Morris, 2016), donde actúa como un competidor muy efectivo, que a largo plazo podría convertirse en una especie dominante que impacta la diversidad de los arrecifes coralinos (Côté, Green, & Hixon, 2013; Albins, 2015). Estudios experimentales controlados de campo demuestran que el pez león afecta la abundancia local de los peces nativos, lo que puede reflejarse en la diversidad de estos ecosistemas marinos (Green, Akins, Maljković, & Côté, 2012; Ballew, Bacheler, Kellison, & Schueller, 2016).
El presente estudio se realizó en dos parques nacionales -Guanahacabibes, Cuba (PNG) y Arrecifes de Xcalak, México (PNAX)- ubicados en el Mar Caribe. Ambos cuentan con información sobre la estructura, composición y diversidad de sus comunidades de peces nativos (Cobián & Chevalier Monteagudo, 2009; Cobián, Claro, Chevalier, Perera, & Caballero, 2011; Schmitter-Soto et al., 1998; Morales-Aranda, Schmitter-Soto, & Herrera Pavón, 2012). En el PNG y el PNAX se detectó el pez león desde 2009 (Chevalier et al., 2008; Sabido-Itzá, Gómez Poot, Medina, García Rivas, & Hadad López, 2012). Monitoreos realizados en ambas áreas demuestran que su población se incrementó de manera significativa (Cobián et al., 2016; Sabido-Itzá et al., 2012). El objetivo de este estudio fue analizar la diversidad de las comunidades de peces nativos (que pudieran ser afectadas por el pez león) en dos áreas marinas protegidas del Caribe y compararla con datos previos a la invasión del pez león.
Materiales y métodos
Zona de estudio: El Parque Nacional Guanahacabibes (PNG) se ubica en la zona más occidental de Cuba con un área total de 39 830 ha. La zona de estudio se extendió desde Cabo Corrientes (21°45’ N; 84°30’W) hasta las cercanías del Verraco (21° 91’ N, 84° 61’ W) (Figura 1). Los arrecifes estudiados son tipo costero y su perfil característico generalmente es el de una terraza única (González-Ferrer et al., 2007) que culmina en un escarpe profundo de estructura variada y gran arquitectura. El Parque Nacional Arrecifes de Xcalak (PNAX), en México, abarca una superficie total de 17 949 456 ha, ubicado al sur de Quintana Roo. El área de muestreo se extendió desde Xahuayxol (18° 30’ N; 87°45’W) hasta Bacalar chico (18°12’N; 87°49’W) (Figura 1). Está área presenta una barrera arrecifal semi-continua a lo largo de la costa, que divide la laguna arrecifal (dominada por pastos marinos y parches aislados de corales escleractinios) del frente arrecifal (elevada diversidad y densidad de corales y gorgonias) (Núñez-Lara, González-Salas, Ruiz-Zárate, Hernández-Landa, & Arias-González, 2003).
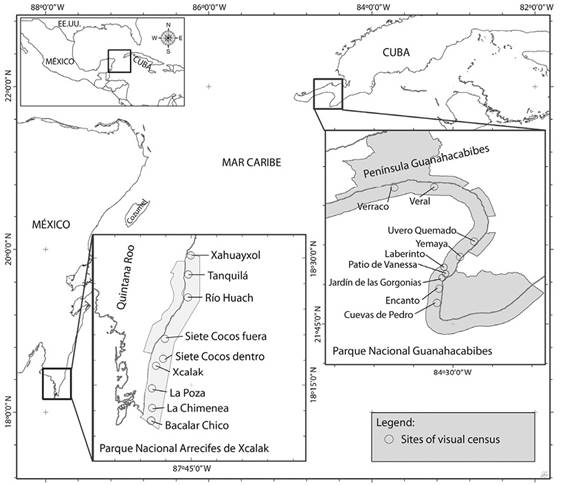
Figura 1 Sitios de muestreo en el Parque Nacional Guanahacabibes (Cuba) y Parque Nacional Arrecifes Xcalak (México).
Comunidades de peces: Se realizaron conteos visuales estacionarios (Bohnsack & Bannerot, 1986) en nueve sitios del PNG, y otros nueve del PNAX (Figura 1), durante las épocas de secas (PNG: febrero 2014 y 2015; PNAX: abril 2014 y 2015) y lluvias (PNG: junio 2013 y 2014; PNAX: septiembre 2014 y 2015). Ambos arrecifes fueron muestreados cuatro veces, y en cada campaña se realizaron 20 censos por sitio. En cada unidad de muestreo se contabilizó el número total de individuos por especie, dentro de un cilindro imaginario de 5 m de radio con el observador en el centro, rotando 360° durante cinco minutos. Los datos fueron registrados por dos ictiólogos de experiencia. En el PNG se trabajó en el arrecife frontal entre 12-15 m de profundidad (cuatro sitios umbral, tres sitios veril, dos sitios camellones y cangilones), mientras que en el PNAX se muestreó en el arrecife frontal en la misma profundidad que en Cuba (cinco sitios camellones y cangilones, dos sitios en el borde de la pared arrecifal y un sitio grieta) y en el interior de la laguna arrecifal (3-5 m) (un sitio: cabezos y parches).
Antes-después del pez león: En Cuba, se tienen registros de las comunidades de peces de cuatro sitios (Cuevas de Pedro, Encanto, Yemayá y Jardín de las Gorgonias) de los años 2003, 2004, 2007, 2008 y 2010, y en México de tres sitios (Xahuayxol: 1997, 1998, 2006, 2010; Río Huach: 1997, 1998, 2006; y Xcalak: 2006, 2007, 2010). Para la comparación, se realizaron muestreos en los mismos sitios teniendo en cuenta los métodos empleados históricamente. En el PNG se ejecutaron censos visuales en seis transectos lineales de 50 m de largo por 2 m de ancho (Brock, 1954) desde el 2003-2015. En el PNAX se realizaron 20 censos estacionarios (Bohnsack & Bannerot, 1986) en cada sitio en los años 2014-15, que se compararon con 10 censos realizados en Rio Huach y 27 censos en Xcalak en 2006-2007. En caso de esfuerzo desigual, los conteos en cada sitio fueron estandarizados por rarefacción (Colwell, 2009) para poder establecer comparaciones.
A partir de la abundancia total, en cada sitio se calculó el índice de riqueza de Margalef (D’n) (Margalef, 1951) definido como: D’= S-1/Ln N (S= número de especies de la muestra, N= número total de individuos en la muestra); el índice de diversidad de Shannon (H’n) (Shannon & Weaver, 1949) definido como H’n= Ss i=1 p i Ln p i (pi= probabilidad de presencia de la especie i (número de individuos de la especie i en la muestra dividida entre el número total de individuos en la muestra); y el índice de equidad de Pielou (J’n) (Pielou, 1966), definido como J’= H’/LnS.
Los datos fueron sometidos a prueba de normalidad (Shapiro-Wilk) y homoscedasticidad (Levene) y presentaron una distribución normal y homogeneidad de varianza, por lo que no fue necesaria ninguna transformación. Se realizaron ANOVAs anidados para determinar las diferencias de los tres índices mencionados, así como del número de especies, entre el PNG y PNAX, entre sitios dentro de una misma AMP, y con respecto a la época climática. No fue necesario aplicar técnicas de rarefacción o acumulación de especies debido a que el esfuerzo fue el mismo.
Se realizó una prueba de t de Student para comparar las medias de los índices de diversidad (H’ y J’) por sitio (PNG: Cuevas de Pedro, Encanto, Jardín de las Gorgonias y Yemayá; PNAX: Xahuayxol, Río Huach y Xcalak) teniendo en cuenta como factor la ausencia (PNG: 2003, 2004, 2007 y 2008; PNAX: 1997-1998, 2006 y 2007) y la presencia de pez león (PNG: 2013, 2014 y 2015; PNAX: 2010, 2014 y 2015). En el caso del PNAX se usaron curvas de rarefacción para analizar la diversidad debido a que el esfuerzo de muestreo fue diferente cada año en cada sitio (Xahuayxol, Río Huach y Xcalak). Para el análisis de los datos se utilizó el programa Statistica 8.0 (Statsoft, Inc.) y EstimateS (Colwell, 2009).
Resultados
Comunidades de peces: Hubo diferencias significativas entre AMPs para el número de especies de peces (F1= 16.74, p<0.001), pero no entre sitios de cada AMP (Figura 2A). El PNG mostró mayores valores de riqueza que el PNAX (43.47±5.14 y 40.22±4.96, respectivamente). El sitio de menor riqueza de las dos AMP fue Xcalak con 35 especies.
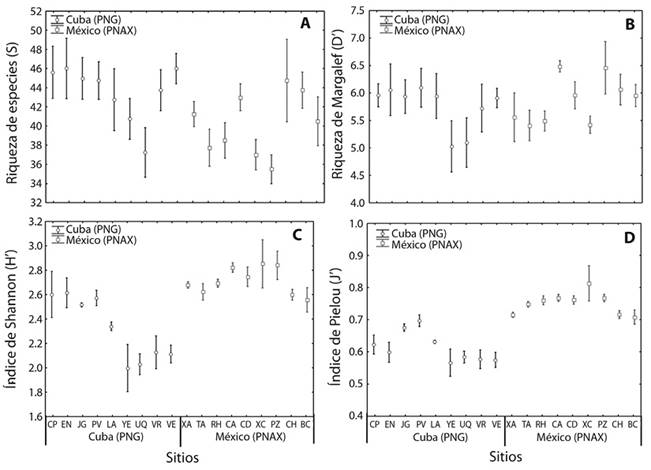
Figura 2 Variación de índices de diversidad en los sitios del Parque Nacional Guanahacabibes (Cuba, PNG) y del Parque Nacional Arrecifes de Xcalak (México, PNAX). Número de especies (A), Índice de Margalef (B), Índice de diversidad de Shannon (C), Índice de equidad de Pielou (D). Las barras muestran la media ±EE.
El índice de riqueza de Margalef mostró diferencias significativas entre ambas AMP (F1= 15.16, p<0.001), mientras que los sitios de cada AMP no mostraron diferencias significativas entre ellos (Figura 2B). El PNAX mostró mayores valores del índice de riqueza de Margalef (5.9±0.64) que el PNG (5.6±0.79). El índice de diversidad de Shannon manifestó diferencias significativas entre las dos AMP (F1= 16.07, p<0.001) y entre los sitios de cada AMP (F1= 31.70, p<0.001) (Figura 2C). Los valores de diversidad fueron mayores en el PNAX (2.71±0.19) que en el PNG (2.34±0.31). Los sitios de mayor diversidad en el PNAX fueron: La Poza, La Chimenea y Bacalar Chico, mientras, que en el PNG se destacó Cuevas de Pedro, Encanto y Patio de Vanesa. El índice de equidad de Pielou presentó un patrón similar, diferencias significativas entre las AMP (F1= 25.19, p<0.001) y entre los sitios de cada AMP (F1= 2.96, p = 0.001) (Figura 2D). El PNAX mostró mayores valores (0.75±0.05) que el PNG (0.62±0.06).
Los análisis de varianza aplicados al número de especies y a los índices de Margalef, de equidad y de Shannon no mostraron diferencias significativas con respecto a la época climática dentro de cada AMP ni entre AMP (Figura 3A-D). En los Cuadros 1 y 2 se muestran los valores de los índices por sitio en cada AMP, para lluvia y seca.
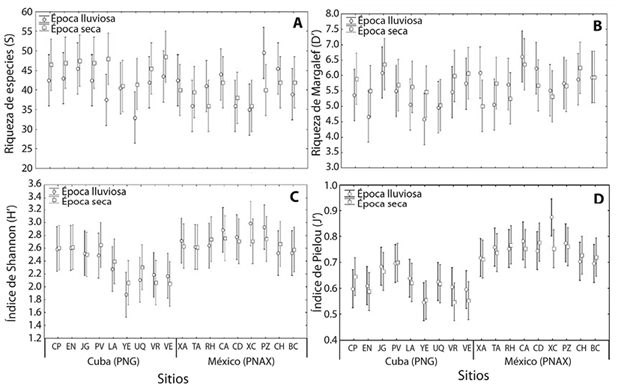
Figura 3 Variación en los índices de diversidad en épocas de seca y lluvia por año en sitios del Parque Nacional Guanahacabibes (Cuba, PNG) y del Parque Nacional Arrecifes de Xcalak (México, PNAX). Número de especies (A), Índice de Margalef (B), Índice de equidad de Pielou (C), Índice de diversidad de Shannon (D). Las barras muestran la media ±EE.
Cuadro 1: Especies más abundantes en el Parque Nacional Guanahacabibes (Cuba) y Parque Nacional Arrecifes de Xcalak (México). TABLE 1: Most abundant species in the Guanahacabibes National Park (Cuba) and Xcalak Reef National Park (Mexico)
| Especies | Abundancia % (PNG) | Abundancia % (PNAX) |
|---|---|---|
| Coryphopterus personatus/hyalinus | 27.49 | 2.14 |
| Clepticus parrae | 19.66 | 4.74 |
| Stegastes partitus | 10.18 | 11.8 |
| Chromis cyanea | 10.01 | 10.03 |
| Gramma loreto | 4.35 | 2.6 |
| Thalassoma bifasciatum | 4.32 | 14.23 |
| Chromis multilineata | 2.99 | 0.47 |
| Gramma melacara | 2.82 | 0 |
| Halichoeres garnoti | 1.39 | 4.35 |
| Ocyurus chrysurus | 1.15 | 1.04 |
| Canthigaster rostrata | 0.92 | 4.12 |
| Acanthurus coeruleus | 1.08 | 4.07 |
| Pterois volitans | 0.65 | 0.36 |
| Scarus taeniopterus | 0.61 | 3.52 |
| Abudefduf saxatilis | 0.04 | 3.3 |
| Scarus iseri | 0.42 | 2.99 |
Cuadro 2: Variación del índice de diversidad de Shannon (H’n, en nats por individuo), índice de equidad de Pielou (J’n) y abundancia (media del número de individuos total de pez león) en cuatro sitios del Parque Nacional Guanahacabibes (CP: cuevas de Pedro, EN: Encanto, JG: Jardín de las Gorgonias y YE: Yemayá) antes y después de la presencia de pez león TABLE 2: Variation of Shannon diversity index (in nats per individual), Pielou evenness index (J’n), and abundance (average number of lionfish) at four sites in Guanahacabibes National Park (CP: Cuevas de Pedro, EN: Encanto, JG: Jardín de las Gorgonias y YE: Yemayá) before and after the presence of lionfish
| Año | Épocas | Sitios | Ausencia-presencia de pez león | Abundancia (Media del número de individuos total de pez león) | H’n | T-Student | J’n | T-Student |
|---|---|---|---|---|---|---|---|---|
| 2003 | Lluvia | CP | Ausencia | 0.00 | 2.95 | NS | 0.62 | NS |
| 2004 | Seca | CP | Ausencia | 0.00 | 3.40 | 0.70 | ||
| 2007 | Lluvia | CP | Ausencia | 0.00 | 2.83 | 0.69 | ||
| 2008 | Seca | CP | Ausencia | 0.00 | 2.55 | 0.64 | ||
| 2013 | Lluvia | CP | Presencia | 2.05±1.35 | 2.85 | 0.56 | ||
| 2014 | Seca | CP | Presencia | 0.55±0.60 | 2.23 | 0.60 | ||
| 2014 | Lluvia | CP | Presencia | 0.65±0.74 | 2.33 | 0.64 | ||
| 2015 | Seca | CP | Presencia | 0.50±0.51 | 2.99 | 0.69 | ||
| 2003 | Lluvia | EN | Ausencia | 0.00 | 2.78 | NS | 0.59 | NS |
| 2004 | Seca | EN | Ausencia | 0.00 | 3.39 | 0.71 | ||
| 2007 | Lluvia | EN | Ausencia | 0.00 | 2.66 | 0.67 | ||
| 2008 | Seca | EN | Ausencia | 0.00 | 2.74 | 0.66 | ||
| 2013 | Lluvia | EN | Presencia | 0.40±0.58 | 2.88 | 0.57 | ||
| 2014 | Lluvia | EN | Presencia | 0.55±0.60 | 2.34 | 0.65 | ||
| 2015 | Seca | EN | Presencia | 0.65±0.81 | 2.48 | 0.65 | ||
| 2003 | Lluvia | JG | Ausencia | 0.00 | 3.14 | NS | 0.65 | NS |
| 2004 | Seca | JG | Ausencia | 0.00 | 3.58 | 0.73 | ||
| 2007 | Lluvia | JG | Ausencia | 0.00 | 2.65 | 0.64 | ||
| 2008 | Seca | JG | Ausencia | 0.00 | 2.68 | 0.68 | ||
| 2013 | Lluvia | JG | Presencia | 0.90±1.07 | 2.55 | 0.71 | ||
| 2014 | Seca | JG | Presencia | 0.80±1.05 | 2.48 | 0.68 | ||
| 2014 | Lluvia | JG | Presencia | 0.65±0.62 | 2.50 | 0.66 | ||
| 2015 | Seca | JG | Presencia | 0.50±0.60 | 2.54 | 0.65 | ||
| 2003 | Lluvia | YE | Ausencia | 0.00 | 2.79 | t=2.5326 p=0.0401 | 0.62 | NS |
| 2004 | Seca | YE | Ausencia | 0.00 | 3.25 | 0.67 | ||
| 2007 | Lluvia | YE | Ausencia | 0.00 | 2.56 | 0.58 | ||
| 2008 | Seca | YE | Ausencia | 0.00 | 2.23 | 0.54 | ||
| 2013 | Lluvia | YE | Presencia | 1.55±1.90 | 1.74 | 0.52 | ||
| 2014 | Seca | YE | Presencia | 0.70±0.75 | 1.63 | 0.56 | ||
| 2014 | Lluvia | YE | Presencia | 0.70±0.80 | 2.03 | 0.58 | ||
| 2015 | Seca | YE | Presencia | 0.50±0.60 | 2.50 | 0.65 |
En el PNG se registraron 152 especies de peces y en el PNAX 139, donde el PNG tuvo una media por sitio de 45 especies y el PNAX de 40 especies (Figura 4 y 5). Las especies dominantes (Cuadro 1) fueron más abundantes en el PNG (Coryphopterus personatus/hyalinus, Clepticus parrae, Gramma loreto, G. melacara y Chromis multilineata) que, en el PNAX, donde las comunidades de peces fueron más homogéneas. Destacó el pez león con 522 individuos en el PNG (8-31 ind/sitio) y sólo 114 individuos para el PNAX (0-8 ind/sitio).
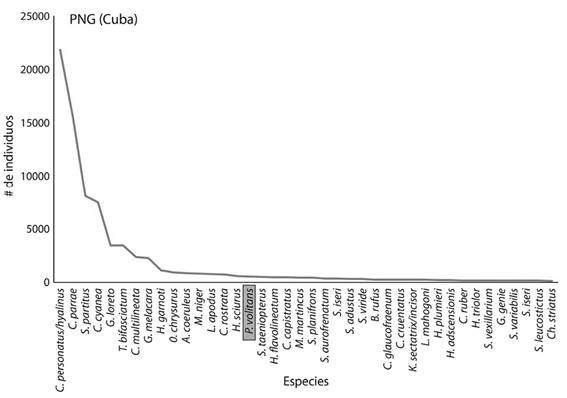
Figura 4 Número de individuos de las 40 especies más abundantes en el Parque Nacional Guanahacabibes (PNG).
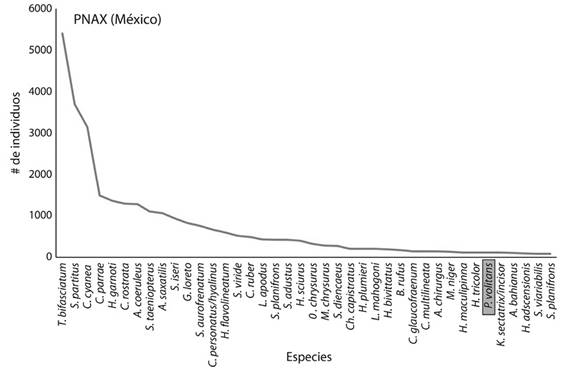
Figura 5 Número de individuos de las 40 especies más abundantes en el Parque Nacional Arrecifes de Xcalak (PNAX).
Antes-después del pez león: Los índices de diversidad y equidad en función de la ausencia (2003-2008) y presencia (2009-2015) del pez león en el PNG no mostraron cambios en la mayoría de los sitios. Yemayá fue el único sitio que mostró diferencias significativas en el índice de diversidad de Shannon en función de la ausencia (H’media: 2.7 ± 0.42) o presencia (H’media: 1.97 ± 0.38) del pez león. El resto de los sitios presentaron valores similares de diversidad y equidad (Cuadro 2). En el PNAX, no se observaron diferencias significativas en estos índices en ningún sitio a partir de los resultados de la prueba de t de Student (Cuadro 3). Al analizar la diversidad ponderada por curvas de rarefacción solo se encontraron diferencias significativas en Río Huach: la diversidad bajó de 3.11 ± 0.08 en 2006-07 a 2.57 ± 0.13 en 2014-15 (F= 27.2; p<0.05) y Xcalak disminuyó su diversidad de 3.3±0.08 en 2006-07 a 2.9±0.07 en 2014-15 (F=9.80; p<0.05).
Cuadro 3: Variación del índice de diversidad de Shannon (H’n, en nats por individuo), índice de equidad de Pielou (J’n) y abundancia (media del número de individuos total de pez león) en tres sitios del Parque Nacional Arrecifes de Xcalak (XA: Xahuayxol, RH: Río Huach, XC: Xcalak) antes y después de la presencia de pez león TABLE 3: Variation of Shannon diversity index (H’n, in nats per individual), Pielou evenness index (J’n) and abundance (average number of lionfish) at three sites in Xcalak Reef National Park (XA: Xahuayxol, RH: Río Huach, XC: Xcalak) before and after the presence of lionfish
| Año | Épocas | Sitios | Ausencia-presencia de pez león | Abundancia (Media del número de individuos total de pez león) | H’n | T-student | J’n | T-student |
|---|---|---|---|---|---|---|---|---|
| 1997-1998 | Seca | XA | Ausencia | 0.00 | 2.80 | NS | 0.70 | NS |
| 1997-1998 | Lluvia | XA | Ausencia | 0.00 | 2.50 | 0.60 | ||
| 2010 | Seca | XA | Ausencia | 0.00 | 2.70 | 0.70 | ||
| 2010 | Lluvia | XA | Presencia | 0.04± | 2.90 | 0.70 | ||
| 2014 | Seca | XA | Presencia | 0.20±0.41 | 2.64 | 0.72 | ||
| 2014 | Lluvia | XA | Presencia | 0.25±0.55 | 2.70 | 0.70 | ||
| 2015 | Seca | XA | Presencia | 0.40±0.50 | 2.63 | 0.71 | ||
| 2015 | Lluvia | XA | Presencia | 0.35±0.80 | 2.74 | 0.74 | ||
| 1997 | Seca | RH | Ausencia | 0.00 | 2.87 | NS | 0.76 | NS |
| 1998 | Lluvia | RH | Ausencia | 0.00 | 1.90 | 0.50 | ||
| 2006 | Lluvia | RH | Ausencia | 0.00 | 3.20 | 0.86 | ||
| 2014 | Seca | RH | Presencia | 0.10±0.30 | 2.58 | 0.74 | ||
| 2014 | Lluvia | RH | Presencia | 0.15±0.36 | 2.46 | 0.75 | ||
| 2015 | Seca | RH | Presencia | 0.35±0.48 | 2.66 | 0.76 | ||
| 2015 | Lluvia | RH | Presencia | 0.25±0.55 | 2.79 | 0.77 | ||
| 2006 | Lluvia | XC | Ausencia | 0.00 | 2.89 | NS | 0.80 | NS |
| 2007 | Seca | XC | Ausencia | 0.00 | 2.57 | 0.70 | ||
| 2010 | Seca | XC | Ausencia | 0.00 | 2.30 | 0.60 | ||
| 2010 | Lluvia | XC | Ausencia | 0.00 | 2.80 | 0.71 | ||
| 2014 | Seca | XC | Presencia | 0.30±0.47 | 2.11 | 0.70 | ||
| 2014 | Lluvia | XC | Presencia | 0.00 | 2.39 | 0.68 | ||
| 2015 | Seca | XC | Presencia | 0.00 | 2.51 | 0.74 | ||
| 2015 | Lluvia | XC | Presencia | 0.25±0.71 | 2.10 | 0.67 |
Discusión
En el PNG se registró menor número de especies de peces que los inventariados en otros estudios (Cobián et al., 2011), lo cual pudiera estar relacionado con el tipo de metodología utilizada. En esta área tradicionalmente se han realizado transectos lineales, los cuales se reconocen como muy efectivos para evaluar la diversidad, al cubrir un área relativamente grande y considerarse más útil para los organismos móviles y con distribución parchada (Claro & García-Arteaga, 1994). El método estacionario es muy eficiente para determinar la frecuencia y abundancia relativa de los peces de manera rápida y con un mayor número de réplicas que le confieren alta confiabilidad estadística (Bohnsack & Bannerot, 1986). En el caso del PNAX se observaron 15 especies que no habían sido observadas en estudios anteriores con la misma metodología (Schmitter-Soto, Cruz-Martínez, Herrera, & Hernández, 2007; Morales-Aranda et al., 2012). El biotopo de camellones y canales de la pendiente arrecifal del PNG presentó mayor riqueza debido a la elevada complejidad del relieve, que proporciona abundantes refugios y alimento para los peces; estos resultados coinciden con los encontrados por Cobián et al. (2011) en Cuba y con lo encontrado por otros autores en otras regiones (Alcolado, 1984; Núñez-Lara & Arias-González, 1998).
Los sitios de mayor riqueza en el PNAX se ubicaron en el frente arrecifal, el hábitat de mayor complejidad. La Poza es considerada como un sitio especial debido a su estructura única (un cañón submarino) en el Caribe mexicano, por lo que el plan de manejo del área lo ubica dentro de sus principales objetos de conservación (CONANP, 2004). Por otra parte, La Chimenea y Bacalar Chico se ubican en la zona de protección “Bacalar Chico” según la zonificación funcional del PNAX. En estos sitios está prohibida la pesca, solo se permite buceo contemplativo, investigación científica, monitoreo, restauración ecológica, protección y conservación ecológica. Esta zona limita con Belice, cuya zona está declarada como Parque Nacional y Reserva Marina Bacalar Chico (Grimshaw & Paz, 2004), por lo que la zona sur del PNAX recibe los beneficios de ambas AMP, lo cual podría explicar la riqueza alta de especies. Estos sitios presentaron bajos valores de diversidad y equidad, debido a la dominancia de unas pocas especies.
Xcalak se ubica en la zona de aprovechamiento sustentable del PNAX, donde se permite buceo y pesca, por lo que es evidente la presión de pesca a las que están sometidas las comunidades de peces. Este arrecife manifiesta una degradación debido a la elevada presión de pesca ribereña para consumo y al comercio local, debido al incremento del turismo en la zona a pesar de ser un AMP (Garza-Pérez, 1999; Ruiz-Zárate, Hernández-Landa, González-Salas, Núñez-Lara, & Arias-González, 2003). Otro sitio con similar presión de pesca es Siete Cocos Dentro, el cual presentó baja riqueza de especies, además de ubicarse en la laguna arrecifal, hábitat menos complejo; esto coincidió con lo encontrado en la mayoría de los estudios en esta área, mayor riqueza en el frente arrecifal que en la laguna arrecifal, la cual cuenta con grandes extensiones de pastos marinos y arena (Carranza-Sánchez, Molina-lslas, Bezaury-Creel, López-Santos, & McCann, 1994; Morales-Aranda et al., 2012). Esta tendencia ha sido encontrada en arrecifes con igual perfil en la región del Caribe mexicano (Núñez-Lara et al., 2003).
La mayoría de los sitios en PNG se ubican en la pendiente arrecifal (borde de pared). Este hábitat favorece una alta dominancia de unas pocas especies (Coryphopterus personatus/hyalinus, Gramma loreto, Chromis cyanea, C. multilineata, Stegastes partitus, Clepticus parrae), lo que hace que la equidad sea menor y, por lo tanto, el índice de diversidad también. Estas especies conformaron el 73% de todos los individuos en un estudio similar por Cobián y Chevalier Monteagudo (2009) en la misma área. La misma tendencia fue observada en la pendiente arrecifal de Bahía de Cochinos (Cuba) por Chevalier y Cárdenas (2005).
Yemayá y Uvero Quemado fueron los sitios menos diversos en el PNG, lo que pudiera estar relacionado con pesca ilegal en esta zona (aunque no intensa), que se encuentra más desprotegida y accesible a los pescadores. La baja equidad se debe a la dominancia de pocas especies, fundamentalmente C. cyanea. C. multilineata y C. parrae que habitan en la columna de agua, lo que coincide con estudios anteriores (Cobián et al., 2011).
La época climática no tuvo efectos sobre los índices de diversidad. Estos resultados coinciden con los encontrados por Cobián et al. (2011) en los mismos sitios. En el PNAX, tampoco se observaron diferencias en cuanto a las épocas en este estudio. Sin embargo, Schmitter-Soto et al. (2007) encontraron diferencias significativas para el índice de riqueza, específicamente en el sitio Xahuayxol. Con respecto al índice de equidad según Schmitter-Soto et al. (2007), las especies muestreadas en las distintas épocas climáticas para 2010 tuvieron mayor homogeneidad en distribución que en 2000 (Morales-Aranda et al., 2012). Estos autores mencionan que las diferencias estacionales podrían estar relacionadas con la pesca histórica en el área.
El pez león se ubicó entre las especies más abundantes en los arrecifes coralinos del PNG, ocupando el lugar 16 en la lista a solo seis años de su primer registro. En el PNG no existe un programa de manejo dirigido para el pez león, solo se han realizado capturas con fines científicos (Cobián et al., 2016). En cambio, en el PNAX se ejecuta un programa eficiente de manejo desde el 2009 a través, de torneos de pesca y programas gubernamentales de empleo temporal (Sabido-Itzá, Medina, Navarrete, Gómez-Poot, & García-Rivas, 2016). En el PNAX el pez león ocupó el lugar 35 y fue menos abundante, lo que pudiera indicar el efecto positivo de los programas sistemáticos de remoción (Barbour, Allen, Frazer, & Sherman, 2011; De León et al., 2013). No obstante, en el PNG existen otros factores ambientales y físicos que favorecen la elevada abundancia de pez león, como una mayor complejidad estructural del arrecife (superior a la observada en el PNAX), que provee abundante refugio y alimento a esta especie (Cobián et al., 2016), también pudieran influir la geomorfología de la zona, la dinámica de las corrientes u otras variables abióticas como la temperatura, que sería interesante analizar en otros estudios.
La mayoría de los sitios en el PNG no mostraron cambios en los índices, al comparar datos antes y después de la invasión de pez león. Sin embargo, Yemayá mostró una disminución significativa en el índice de diversidad (bajos valores riqueza, diversidad y equidad), lo que se ha relacionado con la pesca furtiva del área. Yemayá presentó un incremento de pez león de manera significativa del 2009-2013, (Cobián et al., 2016) donde se mantienen las elevadas abundancias de pez león (0.086 ± 1.20). En este caso se aplicó la metodología de conteos estacionarios a una escala menor, mientras, que Cobián et al. (2016) aplicaron transectos lineales, abarcando una mayor área. También se consideraron abundantes los mesodepredadores nativos, Cephalopholis cruentata (0.37 ± 0.055), Lutjanus apodus (0.075 ± 3.21) y C. fulva (0.010 ± 0.30), los cuales no son objeto de pesca en la zona. Según evidencias de estudios anteriores, los efectos negativos en las comunidades de peces podrían ser mayores cuando estén presentes los depredadores nativos y el pez león (Albins, 2013), lo cual se uniría a los efectos de extracción existentes del pez león. Estos efectos no fueron detectables a seis años de la invasión en el PNG, por lo que se sugiere continuar con los monitoreos en esta área, con diseños que puedan detectar estos impactos a largo plazo, ya que el pez león pudiera desestabilizar la dinámica depredador-presas nativas, donde el invasor puede causar tres veces una mortalidad mayor que los mesodepredadores nativos (Ingeman & Webster, 2015).
Rio Huach y Xcalak reportaron bajas abundancias de pez león durante este estudio (Río Huach: 0.02±0.35; Xcalak: 0.013±0.44), lo cual pudiera deberse al programa de captura de pez león que organiza el PNAX desde 2009 a través de los programas de empleo temporal. Por lo anterior, asumimos que los cambios en la diversidad detectados se deben fundamentalmente a la pesca histórica del área (Schmitter-Soto et al., 2007; Morales-Aranda et al., 2012).
Las diferencias encontradas en cuanto a riqueza, diversidad y equidad en las comunidades de peces en ambas AMP no parecen estar asociadas a la abundancia del pez león y sí a la estructura particular de los arrecifes y a los efectos de la pesca. En arrecifes de Venezuela, Elise, Urbina-Barreto, Boadas-Gil, Galindo-Vivas y Kulbicki (2015) no detectaron cambios significativos en la estructura de las comunidades de peces nativos al analizar los efectos del pez león. Sin embargo, Green et al. (2012), Côté et al. (2013), Albins (2013), Albins, (2015) y Dahl, Patterson III y Snyder (2016) mencionan que el pez león ha causado disminuciones rápidas y significativas en la riqueza y diversidad de especies en los arrecifes. Sugerimos continuar los monitoreos en las AMP de Cuba y México, así como la implementación de experimentos que permitan revelar el efecto real del pez león sobre las especies presentes para detectar posibles cambios en la estructura de las comunidades de peces nativos. También, recomendamos trazar estrategias de control dirigidas al pez león: torneos de pesca, colocación de trampas y redes en zonas profundas, motivar su consumo o el uso en la artesanía. Algunas de estas acciones, como la promoción de consumo y torneos de pesca, han sido efectivas en otros sitios del Caribe (Morris & Akins, 2009). La implementación adecuada de estas estrategias puede contribuir a la conservación de las especies nativas en los arrecifes del PNG y PNAX.

