











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Durante muchas décadas las plantas han sido parte importante en el desarrollo de la humanidad, dentro de ellas encontramos la familia Orchidaceae con unas 28 000 especies que la convierte en la familia de plantas con más especies en el mundo; de ellas unas 7 000 se encuentran en el neotrópico (1), (2); en su mayoría son especies que se encuentran restringidas a las regiones subtropicales y tropicales que forman parte de los ecosistemas terrestres más complejos (3), para Costa Rica se reportan alrededor de 1 400 especies de las cuales un 20% son endémicas (2), (4) y abarcan desde los climas húmedos hasta los secos.
Las plantas epífitas conforman más del 30% de las plantas vasculares que contribuyen a la diversidad de los bosques húmedos tropicales (5) y debido a su complejidad biológica, son excelentes indicadores de la diversidad forestal general en un área, abarcando gran proporción de la diversidad de plantas (6), (7), ya que se adaptan a diferentes condiciones y rangos de dispersión, desde partes cercanas al suelo como en la parte del dosel del bosque donde el flujo de aire es más constate, permitiendo una mejor tolerancia de la radiación directa, disminuye la depredación, mejora su dispersión a través del viento y atrae polinizadores (8). No obstante, las especies epífitas que crecen en hábitats más secos tienen un gran desafío, el obtener humedad y nutrientes adecuados, ya que, para establecerse en un entorno irregular y un poco estresante, requiere varios factores bióticos y abióticos (9), (10).
El establecimiento exitoso de las epífitas puede estar relacionado con las características del forófito (árbol sobre el cual se encuentra la orquídea), según (11), (12), dicha especificidad puede influir en la distribución y abundancia de las especies. Se conoce que la corteza es un factor importante que influye en esta preferencia, sin embargo, las propiedades químicas y físicas de la corteza que permiten crear un especio propicio para su crecimiento son poco conocidos (13). Algunos estudios han demostrado que las epífitas son más comunes en árboles de madera densa con corteza rugosa (14), entre otras características del forófito (planta hospedera) como capacidad de retención de agua de la corteza, edad, crecimiento y pH de la corteza, que influyen en la distribución y diversidad de epífitas (15), (16).
Por su parte, la presencia de estas plantas se ve afectada por ciertos factores como lo es la escasez de precipitaciones (17), lo que hace que las áreas boscosas con precipitación reducida solo se puedan establecer familias más especializadas como Orchidaceae y Bromeliaceae (18). Particularmente, los bosques tropicales caducifolios con estaciones secas definidas pueden ser considerados ambientes marginales para las orquídeas, mostrando una escasez y distribución irregular de las mismas en estos hábitats (19).
Por esta razón, el objetivo de este estudio es determinar la presencia y asociatividad de las orquídeas con especies arbóreas en el bosque seco tropical del Parque Nacional Guanacaste.
Materiales y métodos
Área de estudio: El monitoreo de orquídeas se realizó en áreas de bosque del Parque Nacional Guanacaste, sector Pocosol (figura 1). El sitio registra una temperatura promedio de 25°C y una precipitación media anual de 1500 mm, no obstante, puede variar entre 900 - 2600mm, presenta una estación seca bien marcada de entre 5 y 6 meses del año, generalmente de diciembre a mayo (20), (21).
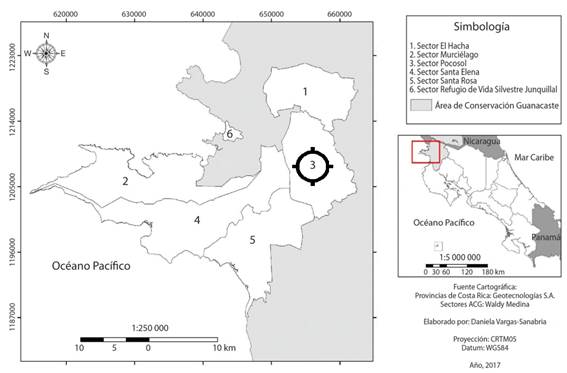
Fuente: (21).
Figure 1. Location of the study area.
Levantamiento de información de campo: En dos parcelas permanentes de monitoreo de una hectárea, cada una de ellas fue subdividida en 25 subparcelas de 20 x 20 m; para el estudio se escogieron las cuatro de las esquinas y una del centro, en un esfuerzo total de muestreo de 0,2 ha por sitio. De manera complementaria se inventarió las orquídeas en dos transectos lineales de 10 x 200 m (uno en cada sitio de estudio), en rumbo norte franco. En el centro del transecto se tendió una cuerda, como referencia para realizar el recorrido de este, con una cobertura visual de cinco metros a ambos lados. En el respectivo formulario se registró la unidad de muestreo (parcela y subparcela o transecto), fecha, ubicación geográfica y coordenadas. Además, se registraron los datos del forófito (árbol sobre el cual se encuentra la orquídea), indicando número de árbol, género, epíteto, diámetro a la altura del pecho (DAP) y los siguientes datos, de acuerdo con el protocolo de mediciones forestales utiliza el Programa de Gestión de Bosques del Instituto de Investigación y Servicios Forestales (PGB-INISEFOR):
Condición del árbol, según figura de códigos.
ADAP: altura a la que se midió el DAP, cuando es diferente a 1,3 m.
Exposición de copa (EXPCOPA) con cinco categorías: 1: Árbol en claro o emergente; 2: Luz plena vertical; 3: Alguna luz vertical; 4: Luz lateral; 5: Sin luz lateral.
DOSEL en el que se encuentra el árbol, con cinco categorías: 1: Emergente; 2: Dosel superior; 3: Dosel intermedio; 4: Dosel inferior y 5: Sotobosque.
Forma de copa (FORMCOPA) con cinco categorías: 1: Copa perfecta, círculo completo; 2: Copa buena, círculo irregular; 3: Copa tolerable, media copa; 4: Copa pobre, menos de media copa y 5: Copa muy pobre, una o pocas ramas.
Tipo de corteza (TIPCORT) con seis categorías: 1: Lisa; 2: Corrugada; 3: Con fisuras transversales; 4: Con fisuras longitudinales; 5: Con lenticelas y 6: Exfoliante.
Espesor de corteza (ESPCORT) con tres categorías: 1: Delgada; 2: Media y 3: Gruesa.
ALTOT: Altura total del árbol en metros.
ALTFUSTE: Altura del fuste en metros.
A cada individuo de orquídea inventariado se le asignó un código único, compuesto por la letra inicial del género y del epíteto; iniciales del sitio; número subparcela o número de transecto y número de consecutivo para el individuo. Además, se indicó la ubicación de la orquídea en el forófito según la clasificación de Johansson 1974 (22), pero realizando una adaptación, ya que se visualizaron líneas horizontales desde el piso del bosque, al considerar que al hacerlo de esa manera se definen zonas con similares condiciones de humedad, temperatura, exposición a la luz, tal como se muestra en la figura 2, zona 0: piso del bosque, zona 1: desde la base del tronco hasta 50 cm; zona 2: desde 50 cm hasta primera ramificación; zona 3: primer tercio ramificación; zona 4: segundo tercio ramificación y; zona 5: último tercio longitud total de ramas.
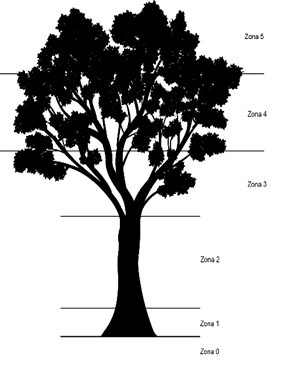
Figure 2. Zones in which phorophyte was divided.
Otro aspecto incluido en el formulario fue si la planta de orquídea se encontraba creciendo en el suelo, dado el hábito terrestre de varias de ellas o si era una epífita que se halló caída en el piso del bosque. Se registró además la fenología de la planta, con los siguientes estados: follaje; cantidad de: flores en botón, flores abiertas, cápsulas inmaduras y cápsulas maduras, así como cualquier otra observación de utilidad. Como ayuda para la ubicación e identificación de orquídeas en campo se utilizó binoculares, pues el bosque presentó condiciones favorables para visualizar todas las ramas.
Análisis de información
La información recolectada permitió analizar la densidad de orquídeas por hectárea y estimar el índice de valor de importancia (IVI), con base en la frecuencia y abundancia relativas. También se calculó la abundancia específica según las distintas variables definidas (zona en el forófito, tipo de corteza y espesor de corteza); a estos datos se les aplicó la prueba de normalidad de Shapiro. Se realizó una prueba de Chi-cuadrado para analizar el grado de asociatividad de las especies por zonas del forófito específicas; consecutivamente se hizo un análisis de correspondencia (Detrended Corrrespondence Analysis) para visualizar esta relación. Por otro lado, se realizó un análisis de varianza utilizando la prueba de Kruskal-Wallis para conocer el nivel de asociación que existe entre las variables zona vertical, tipo de corteza y espesor de corteza con la abundancia de individuos de las especies de orquídeas por forófito; también se estimó la correlación estadística entre el DAP del forófito y la abundancia por medio del método de Spearman. Todos los cálculos estadísticos se realizaron con el software RStudio versión 4.1.1, a un nivel de significancia del 0,05.
Resultados y discusión
En el área muestreada se logró ubicar un total de cuatro especies de árboles fungiendo como forófitos: Swietenia macrophylla (caoba), familia Meliaceae; Rehedera trinervis (melón, yayo), familia Verbenaceae; Quercus oleoides (encino), familia Fagaceae y; Semialarium mexicanum (guácharo), familia Salicaceae. Asimismo, un total de seis géneros y cinco especies de orquídeas fueron inventariadas sobre dichos forófitos: Brassavola nodosa (huele de noche); Encyclia cordigera (Semana Santa); Laelia rubescens, Oncidium spp. Prosthechea fragrans y Stilifolium ascendens. La especie Sacoila lanceolata es una especie terrestre y fue hallada creciendo en el piso del bosque, en estado fenológico de floración.
De acuerdo (23), en el bosque seco del Parque Nacional Barra Honda, es común encontrar las orquídeas en los árboles de cenízaro (Samanea saman), espavel (Anacardium occidentale), cedro (Cedrela odorata) y pochote (Pachira quinata), propios de las áreas bajas. En las partes altas del parque, el principal hospedero es un árbol conocido como flor blanca (Plumeria rubra). Ninguna de esas especies ha sido ubicada dentro de las PPM de Pocosol, pero es importante anotar que la madera de las cuatro primeras es de alto valor comercial, lo cual evidentemente influye en la corta de esas especies y en la consecuente pérdida de hábitat para las orquídeas en las áreas no protegidas.
Se encontró un total de cinco árboles de Q. oleoides que cumplían la función de forófitos, hospedando la mayor cantidad de plantas de orquídea, un total de 16 plantas de tres especies diferentes. Su corteza fisurada, no exfoliante, favorece el establecimiento y permanencia de las orquídeas; esto añade un factor más a la importancia ecológica del encino, pues en estudios anteriores se determinó que es la principal especie arbórea utilizada por las abejas sin aguijón para establecer sus nidos (24). Adicionalmente se enciende la alarma debido a que el encino es una especie sensible a las variaciones climáticas producto de las sequías (25), (26), lo que puede ocasionar su desaparición a futuro y por ende su función de forófito afectando este tipo de epífitas que se establecen en ellos y otras posibles.
Por su parte R. trinervis presentó seis forófitos en el área de estudio, sobre los cuales crecía un total de ocho plantas de orquídea de tres especies, como se muestra en la figura 3. Se destaca la forma del tronco que presenta esta especie, pues su irregularidad o acanaladuras hace que se formen intersticios que favorecen el establecimiento de epífitas y al igual que su tipo de corteza con fisuras longitudinales, que brindan un espacio vital con condiciones óptimas que les permiten establecerse y cumplir su ciclo de vida (13).
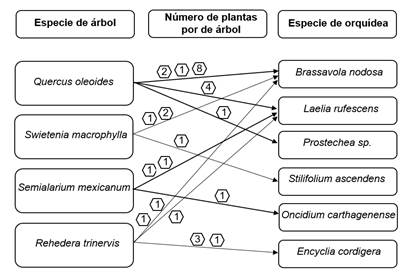
Figure 3. Phorophytes, species and number of orchid plants present.
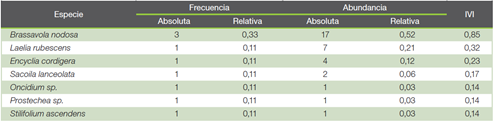
Índice de valor de importancia ecológica de las orquídeas
La especie Brassavola nodosa fue la que presentó mayor importancia ecológica (Cuadro 1) seguido por Laelia rubescens y Encyclia cordigera. Dicho resultado está relacionado con factores ambientales y ecosistémicos que permiten la permanencia de dichas especies en el entorno. Según (27), Brassavola nodosa es una especie que a menudo crece en simpatía con Laelia rubescens, coincidiendo con lo reportado en este estudio y que a su vez fueron las dos especies con mayor IVI reportado. El éxito ecológico de estas especies se asocia con la adaptación ante la variabilidad climática, polinizadores, dispersión, disponibilidad de forófitos para establecerse, entre otros. Por otra parte (27), (28), mencionan que otro factor importante a considerar es la distancia de dispersión del polen, ya que está influenciada por el rango de alimentación del polinizador y/o límites territoriales. Además, influye el potencial que tengan las especies para producir gran cantidad de semillas (29) y una alta facilidad de dispersión en largas distancias (30). En este caso, al ser orquídeas arbóreas, tienen la ventaja de liberar sus semillas más arriba en columnas de aire, donde pueden ser recogidas por las corrientes de viento, lo que aumenta la probabilidad desplazamiento hacia otros sitios (27), esto puede favorecer el establecimiento de estas en el área permitiendo ubicarse entre los valores más altos.
Distribución vertical de las orquídeas
Según la ubicación de las orquídeas en las distintas zonas del forófito, la mayor cantidad de orquídeas se presentó en la zona 5, o sea en la parte superior de la copa; seguido de las zonas 4 y 3 (figura 4), coincidiendo en parte con lo reportado por (7), que obtuvo en un estudio similar la mayor presencia de orquídeas en las zonas 3 y 4. Además, se lograron encontrar dos individuos de Sacoila lanceolata, una especie terrestre (zona 0). Los datos indican una diferencia poco significativa entre zonas (x2=3,376, p-valor=0,193).
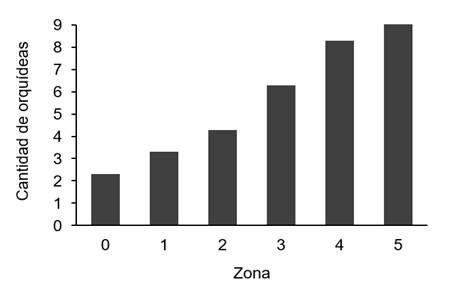
Figure 4. Abundance of orchids by vertical distribution zone in the phorophyte.
Sin embargo, la prueba de Chi-cuadrado mostró que existe una diferencia entre la frecuencia de especies y zonas en particular (x2= 85,416, p-valor<0,05); esto se visualiza en la figura 5, donde es evidente que Encyclia cordigera tuvo preferencia por la zona 2, mientras que Laelia rubescens se encontró presente con mayor frecuencia en la zona 1; así como la especie Sacoila lanceolata, de hábito terrestre.
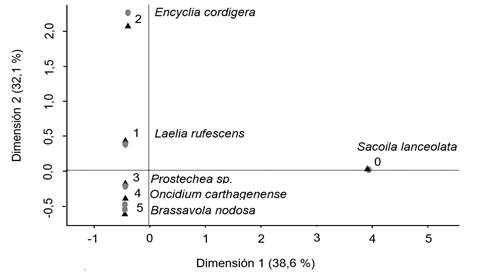
Figura 5 Representación del análisis de correspondencia de las especies de orquídeas por zona en el forófito.
Figure 5. Representation of the correspondence analysis of orchid species by zone in the phorophyte.
Abundancia de orquídeas por características del forófito: Los resultados mostraron una mayor abundancia de orquídeas en los forófitos cuya corteza presenta fisuras longitudinales (tipo 4) seguida de las de tipo corrugado (tipo 2); de forma opuesta, los árboles con cortezas lisas (tipo 1) fueron las de menor abundancia de orquídeas (figura 6); esta diferencia es poco significativa (x2=3,607, p-valor=0,307). De acuerdo con (31), la rugosidad de la corteza y el área de sustrato donde se establecen las orquídeas son características importantes del forófito que influyen sobre la abundancia de estas, coincidiendo con lo obtenido en este estudio.
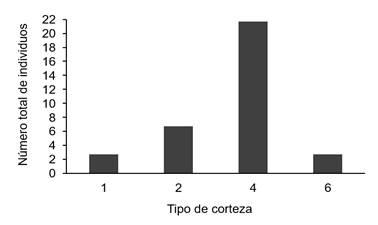
Figure 6. Number of orchids by bark type.
En cuanto al espesor por corteza, el tipo 3 (gruesa) fue el que presentó una mayor abundancia de orquídeas figura 7); mientras que los forófitos con cortezas delgadas (tipo 1) como el Rehedera trinervis presentaron una media de solamente una orquídea por forófito, a excepción de un valor atípico compuesto por varias cepas de Encyclia cordigera; estas diferencias fueron poco significativas (x2=3,290, p-valor=0,193).
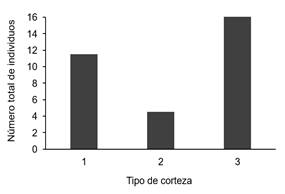
Figure 7. Orchids abundance by bark thickness
Por otra parte, se encontró una relación positiva (p-valor=0.444) entre el diámetro de los forófitos y la abundancia de orquídeas, tal y como se observa en la figura 8. De acuerdo con los resultados se puede observar la presencia de orquídeas en la mayoría de las clases diamétricas, lo que indica tener una ubicación espacial en el sitio siempre y cuanto cuente con forófitos donde puedan establecer. Según (10) esta especificidad de la relación orquídea-forófito puede estar directamente relacionado con su abundancia, lo que es de particular interés, debido a que las epífitas se encuentran confinadas en una distribución irregular y discontinua del sustrato; desempeñando un papel importante en el funcionamiento de los ecosistemas al proporcionar comida y refugio principalmente a insectos y microorganismos (32).
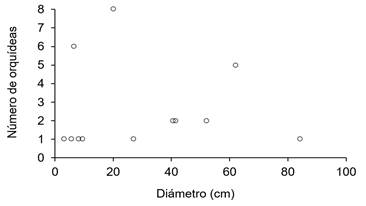
Figure 8. Relationship between orchid abundance and phorophyte diameter.
Conclusiones
La cantidad más considerable de especies de orquídeas se encontró en los forófitos que presentan una corteza fisurada longitudinalmente y no exfoliante, lo que les permite un mejor establecimiento y permanencia.
El Quercus oleoides funge como uno de los principales forófitos en el área de estudio, hospedando la mayor cantidad de plantas de orquídeas.
El mayor número de orquídeas se encontró en la parte superior de las copas y en especies con corteza gruesa, debido a que proporciona mejores condiciones para un desarrollo exitoso en cuanto características de disponibilidad de luz, retención de humedad, sustrato y calidad del microclima.

