











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los manglares son ecosistemas que se ubican en las zonas intermareales de las costas tropicales y subtropicales. Poseen una vegetación que presenta adaptaciones morfológicas, fisiológicas y reproductivas que les permiten adaptarse y sobrevivir en condiciones de salinidad, inundación, fuertes vientos y altas temperaturas (1). Asimismo, son uno de los ecosistemas más productivos (biomasa) en el mundo y poseen un gran valor ambiental y económico. Algunos servicios que brindan son: ser barreras naturales contra tsunamis e inundaciones, evitan la erosión de la costa, sirven de refugio para la biodiversidad, entre otros (2). Además de esto, forman parte importante para los ingresos económicos de los pueblos cercanos a la costa, ya que proveen alimento como los peces y algunos invertebrados, materia prima como leña y madera, entre otros (3).
Uno de los servicios ambientales más importante es su gran potencial para secuestrar y almacenar carbono (4) y pueden almacenarlo con una de las densidades más altas, en comparación con otros bosques en el mundo (5), (6), por lo que cumplen un importante papel en la adaptación y mitigación contra el cambio climático. Sin embargo, gran cantidad de área de manglares en el mundo se ha ido perdiendo debido al desarrollo de la acuicultura, la deforestación, la desviación de los cauces de agua dulce, entre otros (3). Lo anterior ha ocasionado que la estabilidad de estos ecosistemas se encuentre amenazada por la actividad del ser humano y generalmente, no son tomados en cuenta a la hora de realizar esfuerzos de ordenamiento territorial o restauración de ambientes naturales (7), afectando comunidades que dependen del aprovechamiento de los servicios de estos ecosistemas (8).
En Costa Rica, los manglares del Golfo de Nicoya representan el 36 % del área de manglares del país (9). El Golfo de Nicoya es un sitio importante para el cultivo de camarón al punto que, es considerado uno de los más eficientes para la producción de esta especie (10). Además, la producción de sal que se desarrolla en la zona también ha modificado y alterado este ecosistema a lo largo de los años (11). El Área de Conservación Arenal Tempisque otorgó permisos de uso para la producción camaronera y salinera. No obstante, algunos ya vencieron o incurrieron en actos que ocasionaron el cierre de estos, por lo cual, el Sistema Nacional de Áreas de Conservación (SINAC) no amplió su vigencia (12). Esto ha ocasionado que los sitios hayan sido abandonados después de realizadas las actividades de aprovechamiento y se han ido recuperando o regenerando de forma distinta de acuerdo con las condiciones antropogénicas en las que quedaron.
De acuerdo con lo anterior, se esperaría que conforme aumente la edad de abandono de los sitios, se propicie un desarrollo mayor de la vegetación, lo cual va a repercutir en una mayor composición florística, diversidad y almacenamiento de carbono. Por esta razón, se propone el presente estudio, que tiene como objetivo evaluar en una cronosecuencia la diversidad, estructura, composición florística y contenido de carbono de los manglares, en los Humedales Níspero y San Buenaventura-Colorado, ubicados en el Golfo de Nicoya, Costa Rica, con el fin de buscar y proponer acciones que ayuden a la regeneración de estos sitios y a la recuperación de los servicios ecosistémicos y beneficios que brindan.
Materiales y métodos
Área de estudio: El presente estudio se llevó a cabo en los manglares que se encuentran en los Humedales Níspero y San Buenaventura-Colorado, localizados en la costa noreste de la parte interna del Golfo de Nicoya, Costa Rica, entre la desembocadura del Río Tempisque y el Río Abangares (Figura 1). Las comunidades de Colorado y San Buenaventura del cantón de Abangares y la comunidad de Níspero del cantón de Cañas de la provincia de Guanacaste, son las principales dependientes de los recursos que brindan estos ecosistemas (12). El SINAC, mediante las oficinas administrativas del Refugio Nacional de Vida Silvestre Cipancí (RNVSC) del Área de Conservación Arenal Tempisque (ACAT), administra el manejo y conservación de estos humedales (12), (13).
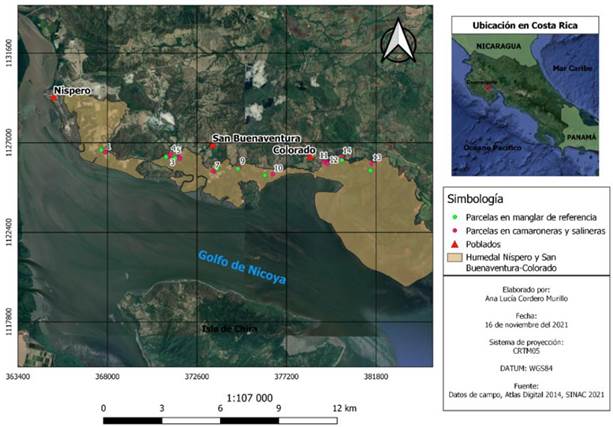
Figura 1 Ubicación de los Humedales Níspero y San Buenaventura- Colorado y de las parcelas de muestreo utilizadas en este estudio, Golfo de Nicoya, Costa Rica, 2021.
Figure 1. Location of Níspero and San Buenaventura-Colorado wetlands and sampling plots used in this study, Golfo de Nicoya, Costa Rica, 2021
En la región predominan las zonas de vida del Bosque Seco Tropical (bs-T) y Bosque Húmedo Premontano Bajo (bh-PB) (13). La temperatura media anual oscila entre los 26 y 28 °C (14) y se encuentra entre los 0 y 300 msnm (15). El lugar se caracteriza por presentar una estación seca con precipitaciones anuales entre 1400 y 2000 mm (11). Las especies de mangle reportadas para el sitio son Rhizophora mangle L. (1753), Rhizophora racemosa G. Mey., Laguncularia racemosa (L.) C.F.Gaertn., Conocarpus erectus L., Avicennia bicolor Standl. y Avicennia germinans (L.) L. (14), (16).
Diseño de muestreo: Se utilizó la metodología descrita en Kauffman et al. (17) y el Manual centroamericano para inventarios de Carbono Azul de Cifuentes-Jara et al. (18). Además, se utilizó datos ya existentes del trabajo realizado por Pineda (19).
Se realizaron muestreos en 11 sitios que anteriormente eran utilizados para la producción de camarón y sal, que actualmente se encuentran cerrados. Además, para conocer el estado del manglar antes de las perturbaciones, se muestreó el manglar intacto o de referencia en ocho sitios. En los sitios se estableció parcelas de muestreo circulares anidadas de 7 m de radio, de forma aleatoria. Para su análisis en una cronosecuencia, las parcelas establecidas en estos sitios se agruparon por rangos de edad (según el año en el que se cerró el aprovechamiento en cada uno, desde el año 2004 hasta el año 2021) de la siguiente manera: de 1 a 10 años: 8 parcelas, de 11 a 20 años: 9 parcelas y en el manglar de referencia: 8 parcelas.
Estructura del manglar, diversidad y composición florística: En cada parcela circular anidada, se midió los árboles con un diámetro mayor o igual a 5 cm. El diámetro se midió a 1,30 m de altura sobre el suelo y en caso de que existiera presencia de raíz fulcrea o alguna irregularidad en el fuste, la medición se realizó a 30 cm por encima de estas. Se tomó datos de altura total y se hizo la identificación taxonómica de cada individuo para los cálculos de estructura y composición florística.
Para la estructura horizontal, se realizó el cálculo del área basal (G) y número de individuos (N) por hectárea, la distribución diamétrica por hectárea del número total de individuos y el Índice de Valor de Importancia (IVI) de las especies presentes en cada estrato establecido por rango de edad. Para la estructura vertical, se realizó la distribución de clases de altura definidas de acuerdo con las categorías de la Unión Internacional de Organizaciones Forestales (IUFRO), la cual define tres estratos de altura a partir de la altura superior del dosel de cada sitio, siendo el piso superior: altura ≥ 2/3 de la altura superior; piso medio: 2/3 > altura ≥ 1/3 de la altura superior; piso inferior: altura < 1/3 de la altura superior del vuelo.
Estimación de contenido de carbono almacenado: Los datos de campo se recolectaron de la siguiente manera: Árboles: en cada parcela circular anidada (18), se midió los árboles con un diámetro mayor o igual a 5 cm. Se determinó si el árbol estaba vivo o muerto y se clasificó en tres categorías, según el estado de descomposición de cada árbol (18).
Regeneración: se utilizó subparcelas de 2 m de radio. Se midió e identificó todos los individuos con una altura mayor a 1,3 m y con diámetro menor a 5 cm.
Madera caída: se utilizó la metodología de plano de intersección (18). A lo largo de cuatro transectos de medición de 12 m de longitud y hasta 2 m de altura, se caracterizó la madera en dos categorías de diámetro: desde los 2 hasta los 5 m del transecto se midieron las piezas con diámetro ≥ 2,5 cm a diámetro < 7,5 cm (piezas medianas) y a lo largo de todo el transecto, se midieron piezas con diámetro ≥ 7,5 cm (piezas grandes). Los planos de intersección estuvieron orientados hacia los puntos NE, NO, SE y SO e iniciaron en el centro de cada parcela circular. A cada pieza de madera con diámetro ≥ 2,5 cm se le asignó una categoría de descomposición.
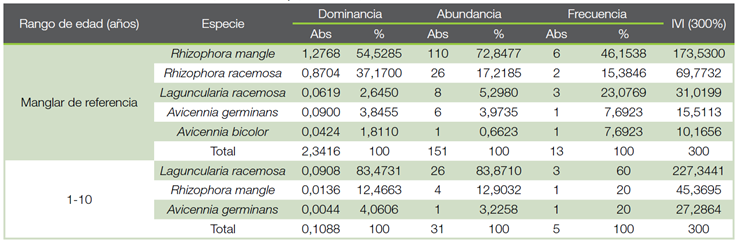
Cuadro 1 Índice de Valor de Importancia (IVI) para las especies presentes por rango de edad en una cronosecuencia, de los árboles con diámetro mayor a 5 cm, en los Humedales Níspero y San Buenaventura-Colorado, Golfo de Nicoya, Costa Rica, 2021.
Sedimento/suelo: para realizar los análisis de carbono correspondientes, se tomó muestras de suelo utilizando un barreno para suelo no consolidado, en un punto de muestreo aleatorio dentro de cada parcela circular. Estas muestras se extrajeron en intervalos de 0-15, 15- 30, 30-50, 50-100 y 100-200 cm de profundidad. Para determinar la densidad aparente del suelo, se tomó una muestra del centro de cada intervalo de profundidad. Luego, el material restante se homogenizó y se tomó una submuestra entre 100 y 200 g, que se llevó al laboratorio y fue secada en horno durante 24 horas a 105 °C, para determinar el carbono por medio el método de combustión seca total.
Para calcular la biomasa de los árboles, la regeneración y las raíces, se utilizó ecuaciones alométricas por especie (18). Para calcular la biomasa de árboles muertos en pie se utilizó un factor de corrección, según su estado de descomposición. La biomasa de la madera caída se calculó con las fórmulas consideradas por Cifuentes- Jara et al. (18). Para el cálculo del carbono en el suelo, se utilizó los datos químicos obtenidos a partir de los análisis realizados en el laboratorio, multiplicando el porcentaje de carbono por el intervalo de profundidad (cm) y por la densidad aparente (gcm3). Para obtener la existencia total de carbono en el suelo hasta la profundidad muestreada, se realizó la sumatoria de los contenidos de carbono por intervalo de profundidad para cada punto de muestreo.
Para calcular el carbono total del ecosistema de manglar, primero se convirtió los datos de biomasa a carbono de cada uno de los componentes (que fueron convertidos a Mg/ha) y luego, se multiplicó este dato de biomasa por la concentración de carbono de cada componente del ecosistema. La información más reciente sugiere que el contenido de carbono en la biomasa ronda alrededor de 0,47 y varía por especie y componente del ecosistema (18). Para la especie R. mangle se utilizó un factor de conversión de 47,52 %; para Pelliciera rhizophorae de 46,67 %; para otras especies de 47 % y para las raíces un factor de 39 %. Luego de hacer esta conversión, se hizo la sumatoria del carbono almacenado en cada componente.
Análisis de datos: Para analizar la estructura del manglar y su composición florística, además del contenido carbono almacenado, se aplicó estadística descriptiva y se generaron gráficos para determinar el comportamiento de las variables evaluadas. La diversidad se analizó por medio del índice de diversidad de Shannon-Wiener para diversidad alfa, utilizando el software Past 4.03.
Resultados y Discusión
Estructura del manglar, diversidad y composición florística: En los Humedales Níspero y San Buenaventura- Colorado, se encontró cinco de las siete especies de mangle presentes en los manglares de Costa Rica (20): R. mangle, R. racemosa (Rhizophoraceae), L. racemosa (Combretaceae), A. germinans y A. bicolor (Acanthaceae). La especie R. racemosa solo está presente en el manglar de referencia, al igual que A. bicolor, que solamente presentó un individuo en este estrato. Por su parte, A. germinans se encuentra presente en todos los rangos de edad, mientras que L. racemosa y R. mangle están presentes en los rangos de edad de manglar de referencia y de 1 a 10 años.
Cuadro 2 Índice de Valor de Importancia (IVI) para las especies presentes por rango de edad en una cronosecuencia, para la regeneración (d<5 cm), en los Humedales Níspero y San Buenaventura-Colorado, Golfo de Nicoya, Costa Rica, 2021.
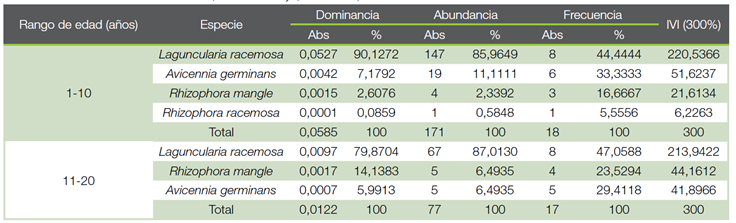
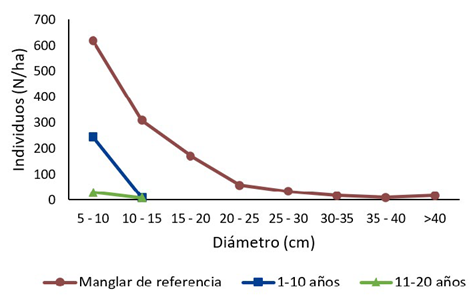
Figura 2 Número de individuos promedio (N/ha) según categoría diamétrica, por rango edad en una cronosecuencia, para los árboles con diámetro mayor a 5 cm, en los Humedales Níspero y San Buenaventura-Colorado, Golfo de Nicoya, Costa Rica, 2021.
Figure 2. Number of individuals per hectare (N/ha) by diameter class distribution in a chronosequence of mangroves, for trees with a diameter above 5 cm, in Níspero and San Buenaventura-Colorado wetlands, Golfo de Nicoya, Costa Rica, 2021.
El Cuadro 1 muestra el Índice de Valor de Importancia (IVI) para cada especie en cada uno de los rangos de edad, para los árboles mayores a 5 cm de diámetro. En el manglar de referencia, la especie con mayor peso ecológico fue R. mangle con un IVI de 173,53 %, seguida por R. racemosa con 69,77 %. En el rango de edad de 1 a 10 años, la especie con mayor IVI corresponde a L. racemosa con un 227,44 % y luego de esta se encuentran R. mangle y A. germinans con 45,37 % y 27,29 %, respectivamente. En el rango de edad de 11 a 20 años, solo se encontró presente la especie A. germinans.
En un estudio realizado por Alfaro-Sibaja et al. (21) en el manglar Palo Seco, Quepos, Costa Rica, determinó que la abundancia de R. mangle se atribuye a que es una especie que se adapta a climas tanto lluviosos como secos y que se caracteriza por estar en ambientes expuestos a fuerte oleaje. Además, en estos ecosistemas existe un gradiente ambiental que hace que ocurra un cambio en la estructura y composición de la vegetación a medida que se aleja de la costa (22).
El Índice de Valor de Importancia (IVI) para las especies presentes en la regeneración, muestra que la especie L. racemosa es la de mayor peso ecológico en ambos estratos, con un 220,54 % en el rango de 1 a 10 años y un 213,94 % en el rango de 11 a 20 años (Cuadro 2). En el estrato del manglar de referencia, solamente se encontró la especie R. mangle. Estos resultados se pueden explicar debido a que L. racemosa corresponde a una especie indicadora de un dosel más abierto, originado por altera-ciones (23). La dominancia de L. racemosa en ambos rangos de edad, seguida por A. germinans en el rango de 1 a 10 años, puede explicarse por la gran producción de propágulos de estas especies (24), además de ser especies pioneras y colonizadoras en áreas donde se han presentado disturbios (25).
El índice de Shannon-Wiener para el manglar de referencia fue de 0,85. Manrow (26) reportó valores de 0,33 y 0,6 para los manglares de Gandoca y Moín, respectivamente. Valores como el obtenido en el presente estudio son de esperarse, ya que en general la diversidad de los ecosistemas de manglar es baja y dominada por algunas especies que son las que presentan las adaptaciones morfológicas y fisiológicas necesarias para crecer en zonas inundadas y tolerar la salinidad (22).
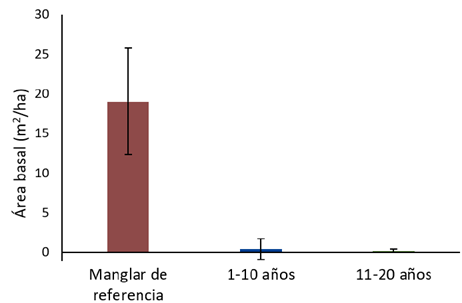
Figura 3 Área basal (m2/ha) de los árboles mayores a 5 cm de diámetro, según los diferentes rangos de edad en una cronosecuencia, en los Humedales Níspero y San Buenaventura-Colorado, Golfo de Nicoya, Costa Rica, 2021.
Figure 3. Mean basal área (m2/ha) of trees with a diameter above 5 cm, in a chronosequence of mangroves, in Níspero and San Buenaventura- Colorado wetlands, Golfo de Nicoya, Costa Rica, 2021.
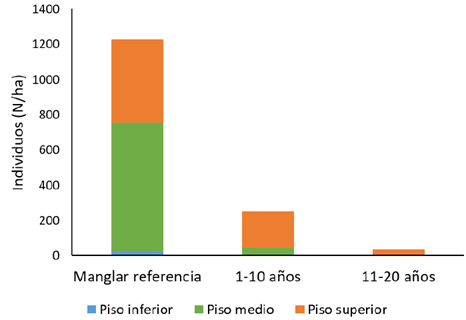
Figura 4 Número de individuos (N/ha) por pisos de altura y según rango de edad en una cronosecuencia, en los Humedales Níspero y San Buenaventura-Colorado, Golfo de Nicoya, Costa Rica, 2021.
Figure 4. Number of individuals (N/ha) by height class in a chronosequence of mangroves, in Níspero and San Buenaventura- Colorado wetlands, Golfo de Nicoya, Costa Rica, 2021.
El rango de edad del manglar de referencia presenta un comportamiento de J invertida, donde el 50,33 % de los árboles se encuentra en la categoría de 5-10 cm, mientras que el 16,24 % se ubica en la última categoría (d
> 40 cm), la cual presentó un pequeño incremento de los individuos con respecto al comportamiento decreciente que se da en todas las categorías anteriores (Figura 2). El estrato de edad de 1-10 años solo presenta árboles en las dos primeras categorías y en la primera de ellas (5-10 cm) se concentra el 96,77 % de los individuos. En el estrato de 11-20 años, ocurre de igual forma que con el estrato anterior, sin embargo, la cantidad de individuos por hectárea es mucho menor.
Esta distribución obtenida de J invertida es típica de los bosques secundarios y bosques maduros saludables (27). Merecí (28) también observó un patrón de J invertida, obteniendo la mayor presencia árboles entre 5 y 20 cm y una baja presencia de individuos de 36-50 cm de diámetro, similar a lo obtenido en este estudio. Lo anterior sugiere que la regeneración de estos manglares podría no estarse viendo tan afectada (7). El proceso de regeneración natural en los manglares se puede ver afectado por la salinidad, la precipitación, la sedimentación, la frecuencia, el nivel y la duración de las inundaciones, entre otros (24), (29) rangos de edad de 1-10 años y de 11-20 años, donde se observa un patrón muy distinto al manglar de referencia, ya que en estos estratos no hay una distribución de J invertida y solo presenta individuos en las dos primeras clases diamétricas.
El área basal promedio de los árboles con diámetro mayor a 5 cm en el estrato del manglar de referencia fue de 19,01 ± 6,75 (n=8) m2/ha, mientras que en el rango de edad de 1-10 años el área basal promedio fue de 0,35 ± 1,30 (n=9) m2/ha y en el de 11-20 años se obtuvo un valor de 0,16 ± 0,21 (n=8) m2/ha (Figura 3). No hubo diferencias significativas entre las medias de los rangos de edad de 1-10 años y de 11-20 años, pero sí las hubo entre estos y el manglar de referencia.
Flores (30) reportó un área basal de 44,42 m2/ha en un manglar en Honduras, mientras que Manrow (26) reportó áreas basales de 26,35 m2/ha en Gandoca y 26,37 m2/ha en Moín, y Cifuentes-Jara et al. (7) reportó un promedio de 12,61 m2/ha para el Golfo de Nicoya. De estos valores, el obtenido para el Golfo de Nicoya es el que más se acerca a lo obtenido en este estudio para el manglar de referencia. Lo anterior podría estar ocurriendo debido a que en el Pacífico Norte de Costa Rica las lluvias son menores y estacionales, lo que ocasiona que los manglares presenten desarrollos estructurales reducidos (31), además de que los manglares del Caribe se distinguen por ser de mayor porte (7).
En los rangos de 1-10 y de 11-20 años el área basal obtenida representó 1,84 % y 0,84 % respectivamente (Figura 3), del valor obtenido para el manglar de referencia, debido a la poca presencia de árboles e incluso a la no presencia de estos en algunos de los sitios. Además, los diámetros de los árboles en estos sitios son mucho menores a los de los individuos presentes en el manglar de referencia. Cifuentes-Jara et al. (7), encontraron
que los manglares que presentaron una estructura con mayor porte fueron los que se encontraban expuestos a aguas menos saladas. Además, Jiménez y Soto (31), encontraron que las mayores alturas y áreas basales coinciden con sitios que presentan un suministro abundante de agua dulce, lo cual no está ocurriendo en ciertos sitios presentes en estos rangos de edad, debido a la existencia de muros o barreras que impiden que ocurra este suministro de agua.
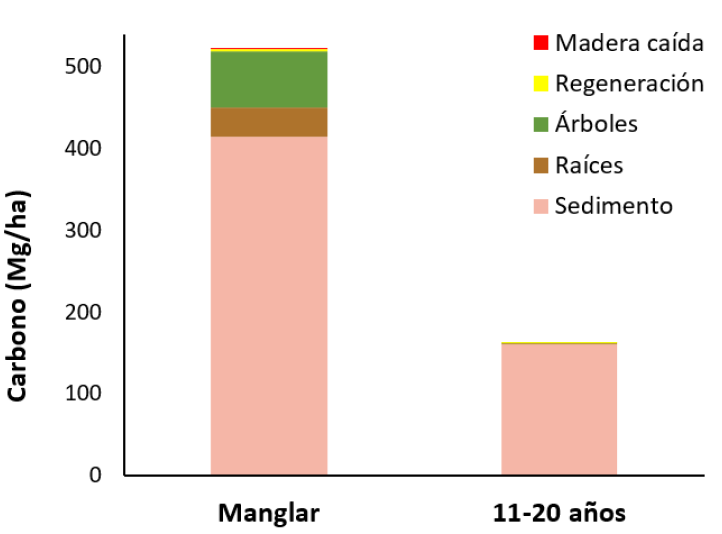
Figura 5 Existencias de carbono (Mg/ha) en el manglar de referencia y rangos de edad para una cronosecuencia, en los Humedales Níspero y San Buenaventura-Colorado, Golfo de Nicoya, Costa Rica, 2021.
Figure 5. Carbon stocks (Mg/ha) in mangroves without impact and other age ranges in a chronosequence of mangroves, in Níspero and San Buenaventura-Colorado wetlands, Golfo de Nicoya, Costa Rica.
La altura total promedio para el manglar de referencia fue de 10 ± 1,73 (n=8) m. En los rangos de edad de 1-10 y de 11-20, la altura total promedio fue de 5,91 ± 2,20 (n=9) m y 5,88 ± 1,75 (n=8) m respectivamente. En el manglar de referencia los pisos de altura son de 5, 10 y 16 m para los pisos inferior, medio y superior, respectivamente. En este rango, aproximadamente el 59 % de los individuos se encuentra en el piso medio y el 38 % en el piso superior. En el rango de edad de 1 a 10 años, el piso inferior fue de 2 m, el medio de 4 m y el superior de 7 m. Aquí los individuos se comportan de forma distinta al rango anterior, ya que más del 80 % se encuentra en el piso superior y aproximadamente el 19 %, en el piso medio, sin presencia de individuos en el piso inferior. En el rango de 11 a 20 años los pisos de altura son 2, 4 y 6 m, para los pisos inferior, medio y superior, respectivamente. En este último rango de edad la totalidad de los individuos se concentra en el piso superior (Figura 4). La altura promedio de los árboles en los manglares rara vez sobrepasa los 20 m (21), (26)-(28), (30). Álvarez (27), reportó en su estudio que el 70 % de los individuos del manglar se encontraron en el piso medio, mientras que en este estudio ese porcentaje fue del 59 %. En los otros dos rangos de edad la altura promedio fue mucho menor en comparación con el manglar de referencia, debido a ser sitios en recuperación y con condiciones muy distintas a las de un manglar maduro.
Estimación de contenido de carbono almacenado: En este estudio, el promedio de las reservas de carbono de la biomasa total de los árboles (arriba del suelo) para el manglar de referencia es de 106, 60 ± 63,34 (n=8) Mg/ ha. Este valor fue menor a lo obtenido en otros estudios: Flores (30), reportó reservas de carbono de 145,84 ± 48,52 Mg/ha, mientras que Bhomia et al. (32), reportaron 147 Mg/ha, ambos para un manglar en Honduras. Por otro lado, Adame et al. (33), reportaron 215 Mg/ha en Laguna Nichupté en la Península de Yucatán, México. El valor promedio para el Golfo de Nicoya fue de 76,75 Mg/ha (7), lo cual corresponde a aproximadamente un 71 % del valor obtenido en este trabajo. Con respecto al rango de edad de 1-10 años, el valor obtenido fue de 13,25 ± 9,51 Mg/ha y en el de 11-20 años de 2,87 ± 3,47 Mg/ha. Esto significa que el manglar de referencia tiene 87,57 % más de carbono almacenado que el rango de 1 a 10 años y 97,31 % más que el rango de 11-20 año, debido al tipo de manejo que recibieron estos sitios en el pasado cuando fueron camaroneras y salineras.
El aporte de la madera caída representa 0,071 % del total de las existencias de carbono del manglar de referencia y es inexistente en los sitios evaluados (Figura 5). Esto se explica debido a que los almacenes de carbono en estos ecosistemas se encuentran principalmente en el componente aéreo (biomasa de árboles vivos y muertos) y subterráneo (sedimento y biomasa de raíces) (17). El componente que más cantidad de carbono representa en ambos rangos de edad, es el del suelo. Las existencias promedio de carbono bajo el suelo a 2 m de profundidad para el manglar de referencia son de 414, 95 ± 47, 84 Mg/ha y para el rango de edad de 11-20 años son de 159, 89 ± 131, 71 Mg/ha. Este valor en el manglar de referencia es casi tres veces mayor a lo obtenido en el rango de edad de 11-20 años (Figura 5). En el manglar de referencia este valor corresponde a aproximadamente el 80 % de las existencias totales. Lo anterior evidencia que un gran porcentaje de este carbono almacenado en el suelo se pierde como resultado de la conversión del manglar a camaroneras y salineras (34), condición que fue confirmada en este estudio.
Para el rango de edad de 1-10 años, debido a inconvenientes de financiamiento, no fue posible obtener datos de carbono en el suelo; sin embargo, se pueden encontrar valores similares o menores al rango de 10-20 años. Lo anterior, ya que las reservas de carbono del suelo dependen de muchos factores (35), es decir, el manglar maduro es un ecosistema donde se lleva a cabo un intenso procesamiento de materia orgánica, debido a que presentan flujos de caída y descomposición de hojarasca, exportación e importación de carbono orgánico particulado y disuelto por efecto del hidroperiodo (36). La principal consecuencia de la deforestación en manglares es la liberación de gases de efecto invernadero como dióxido de carbono (CO2) y metano (CH4) (37). Los resultados obtenidos por Merecí (28), para el manglar maduro, triplican lo obtenido en los suelos bajo camaroneras, por lo que se evidencia el potencial y la importancia de los suelos de manglar, en comparación con los de uso acuícola, como sumideros de carbono.
Acciones para la restauración del manglar de Humedales Níspero y San Buenaventura- Colorado, Golfo de Nicoya: Según los resultados obtenidos y lo observado a lo largo del presente estudio, la regeneración de los sitios se está dando, sin embargo, no al mismo tiempo ni de la misma forma en cada uno. Es necesario llevar a cabo acciones que asistan la recuperación de estos sitios que han sido degradados de distintas formas, teniendo en cuenta que debe existir coordinación con el SINAC y además se deben buscar las respectivas fuentes de financiamiento.
Una acción de restauración que se puede implementar en varios de los sitios estudiados es la restauración/ rehabilitación hidrológica de estos, derribando muros o barreras existentes producto de las antiguas actividades de aprovechamiento, para lograr recuperar el flujo de agua en algunos de los sitios en los que esto no está ocurriendo. Si luego de realizar monitoreos, se han identificado los canales de marea naturales de los sitios, se puede proceder al desazolve de estos. Si se detecta que debido a la perturbación del sitio y al cambio en su topografía es necesaria la habilitación de canales nuevos, se puede realizar esta acción. Para acciones de restauración, es importante considerar hacer uso de la siembra de propágulos de plántulas colectadas o cultivadas, después de determinar que el reclutamiento natural no garantizará la cantidad de plántulas exitosamente establecidas.
Conclusiones
La colonización y desarrollo de los manglares que han sido perturbados puede ser lenta e impredecible, lo cual se evidencia en los sitios en los que ha transcurrido más tiempo desde que se dejó el aprovechamiento, que deberían estarse acercando a un estado similar al manglar de referencia, pero algunos no lo están haciendo e incluso ciertos sitios más jóvenes están teniendo mayor cantidad de regeneración.
La edad de abandono de los sitios no determina la composición de estos, si no las condiciones en las que quedan estos sitios después de que se deja de realizar la actividad de aprovechamiento. Se esperaría que un sitio con mayor cantidad de años en recuperación tenga condiciones, estructura y composición similares a las de un manglar maduro, sin embargo, esto no está ocurriendo ya que, según los resultados obtenidos y las observaciones en campo, existen barreras físicas que impiden que la hidrología de algunos de estos sitios sea la apta.
Un gran porcentaje del carbono almacenado por los manglares en todos sus componentes, pero especialmente en el suelo, se pierde como resultado de la conversión del manglar a camaroneras y salineras; por lo que se evidencia el potencial y la importancia de los suelos de manglar, en comparación con los de uso acuícola, como sumideros de carbono.
Recomendaciones
Para futuros estudios, se sugiere aumentar el número de parcelas por sitio para tener una mejor representación de las condiciones en la que estos se encuentran con respecto a su recuperación. Evaluar los efectos de variables ambientales que no se tomaron en cuenta en este estudio, como la densidad aparente del suelo o la disponibilidad de nutrientes, para lograr comprender de una mejor manera la recuperación que se está dando en estos sitios
Para futuros proyectos o acciones de restauración, tomar en cuenta el involucramiento de las comunidades locales en estos procesos, ya que estas personas son las que se benefician de forma directa de estos ecosistemas. Además, de esta forma se puede sensibilizar a cerca de la importancia que representan los ecosistemas de manglar.

