










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgronomía Mesoamericana
versión On-line ISSN 2215-3608versión impresa ISSN 1659-1321
Agron. Mesoam vol.24 no.1 San Pedro jun. 2013
Recuperación del 15n en la planta de banano y en el suelo de áreas con origen sedimentario
Recovery of 15n in the banana plant within sediment origin soil
Recovery of 15n in the banana plant within sediment origin soil
*Dirección para correspondencia:
Resumen
Entre octubre, 2003 a febrero, 2004 en la Finca Venecia, situada en Matina de Limón, se determinó la movilidad del nitrógeno en el suelo y la recuperación en la planta de banano. Se empleó el procedimiento isotópico con 15N, mediante método directo. Se evaluaron tres fuentes enriquecidas con 15N: úrea (U), nitrato de amonio (NA) y sulfato de amonio (SA), con una dosificación de 424 kg de N por hectárea por año; estas se “monitorearon” a través del perfil a cuatro profundidades: 10, 30, 60 y 90 cm del suelo y el cultivo del banano en cuatro estados fenológicos (F10, Fm, floración y cosecha). Se determinaron formas de N disponibles provenientes del SA, en mayor cantidad en el suelo y en menor cantidad en los tejidos de la planta, lo que indica una menor tendencia a perderse o absorberse del sistema que las otras fuentes. La urea es la fuente nitrogenada que produjo la mayor acumulación de N en la fruta y afectó de forma positiva el peso seco de la planta, en comparación con las otras fuentes nitrogenadas evaluadas (NA y SA). El uso de SA parece tener un efecto negativo en el crecimiento del rizoma de la madre y del hijo de sucesión, por lo cual su uso como única fuente de fertilización nitrogenada no es recomendable. Tal comportamiento podría indicar la necesidad de emplear una fertilización donde la concentración total de N, provenga de una mezcla de varias fuentes.
Palabras clave: estados fenológicos, absorción, concentración de nitrógeno, fertilizante sintético, acumulación de nitrógeno.
Abstract
Between October, 2003 to February, 2004 at the Finca Venecia, located in Matina in Limón, mobility in soil and recovery of banana plant nitrogen was determined, using the direct method with 15N isotopic method. Three sources enriched with 15N: urea (U), ammonium nitrate (NA) and ammonium sulfate (SA) were evaluated, at a dosage of 424 kg N per hectare per year and “monitored” through profile at four depths: 10, 30, 60 and 90 cm of soil and four growth stages (start, F10, Fm, flowering and harvest) of banana cultivation. Forms of N available from the SA was determined in greater quantity in the soil and in lesser amounts in tissues of the plant, indicating a lower tendency to be lost or absorbed from the system than the other sources. Urea is the nitrogen source that produces the greatest accumulation of N in the fruit and positively affects the plant dry weight, compared with other nitrogen sources tested (NA and SA). The use of SA appears to have a negative effect on the growth of the mother and offspring rhizome so its use as the sole source of N fertilization is not recommended. Such situation may indicate the need to use a fertilizer where the total concentration of N, comes from a mixture of several sources.
Key words: phenological stages, absorption, concentration of nitrogen, synthetic fertilizer, nitrogen accumulation.
Introducción
El nitrógeno (N) es considerado uno de los nutrimentos de mayor importancia en el manejo de la fertilización del cultivo del banano, debido a que las necesidades de este nutrimento en la planta de banano son altas, ya que participa en la fotosíntesis, en la respiración y muchos otros procesos metabólicos y fisiológicos (Ortiz et al. 1999). Además, es un componente importante de la estructura de proteínas, ácidos nucleicos, vitaminas, reguladores de crecimiento y muchos otros componentes (López y Espinosa 1995).
Los suelos tropicales suelen ser más deficientes en N que en cualquier otro elemento; las plantas absorben el N principalmente en las formas de amonio (NH4+) y en nitrato (NO3-) (Bertsch 1998). El NO3-, generalmente se encuentra en mayores concentraciones y se puede movilizar hacia las raíces principalmente por flujo de masas (Ramírez 2001, Elizondo 2006). El NH4+ puede adherirse con facilidad a los sitios de intercambio catiónico en los coloides inorgánicos y orgánicos del suelo y puede ser interceptados por las raíces, pues es poco móvil (Ramírez 2001), a pesar de que el transporte hacia la raíz ocurre por flujo de masas a partir de la solución de suelo. Pero debido a un sinnúmero de factores, es posible afirmar que gran parte de los fertilizantes nitrogenados aplicados no lo aprovecha la planta de banano. La volatilización, la lixiviación y la escorrentía explican en parte dicho fenómeno (Soto 1995).
El problema ambiental más importante relativo al ciclo del N, son las pérdidas del nitrógeno del suelo, ya sea en forma de gases, por los procesos de denitrificación y volatilización de óxidos de nitrógeno (N2O), lo que contribuye al rompimiento de la capa de ozono (asociado al calentamiento global). Asimismo, la acumulación de NO3en el subsuelo, por lixiviación, puede incorporarse a las aguas subterráneas o bien ser arrastrados por escorrentía hacia los cauces y reservorios superficiales (Weisenburger 1991).
La precisión en la dosificación y en el manejo de las diferentes fuentes nitrogenadas (urea, nitrato de amonio y sulfato de amonio) de los fertilizantes; asimismo en la distribución, el manejo en cuanto a la aplicación según la época y al estado fenológico de la plantación, son aspectos que deben ser abordados a fin de minimizar los procesos de lixiviación del N hacia las reservas de agua. Por tal motivo, el objetivo de este trabajo fue determinar con 15N la movilidad del nitrógeno a través del perfil del suelo y recuperación en la planta de banano.
Materiales y Métodos
Ubicación del estudio. Para realizar el experimento se seleccionaron lotes representativos de suelos utilizados frecuentemente en la explotación bananera al este del río Reventazón, para ello se escogió la Finca Venecia, que se encuentra ubicada cerca del poblado Venecia en Matina, de Limón. Geográficamente, la finca Venecia se localiza entre las coordenadas Lambert Norte 223.000-227.000 y 618.000-621.000 y el sitio experimental en las coordenadas 618.755 y 224.995 de las hojas cartográficas Matina 3546 III y Moín 3546 II, a una altitud que varía de 10 a 14 msnm.
Área experimental. Se localizó en la tapa derecha del cable 40 entre las torres 8 y 17 de la Finca Venecia, la cual consta de tres parcelas (boquetes) cuyas medidas son de 27,7 m de ancho por 45 m de largo. En donde, cada parcela o boquete se subdividió en tres bloques de 19,5 m de ancho (segregando 4,1 m de los bordes de los canales terciarios) por 12 m de largo (segregando 9 m de borde del canal secundario: dumpling) para un total de nueve parcelas, en donde se distribuyeron los tres tratamientos y las tres repeticiones del diseño experimental. El suelo se clasificó taxonómicamente según el Soil Taxonomy (USDA 1998) como Fluvaquentic Eutrudepts (Acón 2002), de origen aluvial calcáreo y se localiza en relieve plano, con pendientes entre 0 a 3%.
Tratamientos y diseño experimental. En este experimento se evaluaron tres fuentes nitrogenadas: úrea (U), nitrato de amonio (NA) y sulfato de amonio (SA), con una dosificación de 424 kg de N por hectárea por año. Para determinar la movilidad y recuperación del nitrógeno, se emplearon fuentes enriquecidas con 15N y se “monitorearon” a través del perfil del suelo y el cultivo del banano. Para este fin, se muestreó el suelo a cuatro profundidades, 0-10, 10-30, 30-60 y 60-90 cm y las plantas de banano en cuatro estados fenológicos, F10 (primera hoja con ancho de 10 cm), Fm (emisión de la primera hoja verdadera), floración (emisión de la chira) y cosecha (Tavares y Falquez 1997).
Para evaluar la recuperación en el banano y la movilidad en el suelo del N, se utilizó el procedimiento isotópico con 15N mediante método directo. En este proceso se aplica el N enriquecido con 15N y se evalúa la recuperación, movilidad y acumulación mediante la determinación de la relación isotópica 14N/15N en los tejidos de la planta y en el suelo.
Para el tratamiento con urea (U) se aplicaron 38,8 g diluidos, desglosado en 34,5 g de U simple y 4,33 g enriquecida al 10,0% 15N átomos en exceso en la primera fertilización y al 3,0% 15N átomos en exceso en la segunda, tercera, cuarta y quinta fertilización por planta, para el tratamiento de nitrato de amonio (NA) se aplicó 53,3 g, desglosado en 41,73 g de NA simple y 11,61 g, enriquecido al 2,6% 15N en átomos en exceso y para el tratamiento de sulfato de amonio (SA) se utilizó 87,2 g, desglosado en 78,24 g simple y 8,94 g enriquecido al 10,6% 15N en átomos en exceso. Además, cada planta recibió por mes una dosis de 5,0, 31,5, 2,5 y 3,0 g de P2O5, K2O, MgO y S, respectivamente, lo cual fue equivalente a 423,9, 108, 608,4, 54 y 64,8 kg de N, P2O5, K2O, MgO y S, respectivamente por hectárea por año, con una densidad promedio de 1800 plantas por hectárea.
El diseño experimental fue de bloques completos al azar con tres tratamientos (urea, nitrato de amonio y sulfato de amonio) y tres repeticiones. En cada unidad experimental o parcela se escogieron ocho plantas de banano con los estados fenológicos que se evaluaron.
Las cinco aplicaciones de los tratamientos de las tres fuentes nitrogenadas se realizaron los días: 4/10/2003, 4/11/2003, 1/12/2003, 30/12/2003 y 28/01/2004 y después de cada aplicación de los tratamientos se muestrearon los suelos cada 15 y 30 días, 17/10/2003, 31/10/2003, 14/11/2003, 28/11/2003, 15/12/2003, 29/12/2003, 13/1/2004, 28/1/2004, 11/02/2004 y 26/02/2004.
Los muestreos de suelos se realizaron con barreno en la banda de abonamiento a cuatro profundidades (0-10, 10-30, 30-60 y 60-90 cm), de cada profundidad se tomaron tres submuestras las cuales se mezclaron y se conformó una muestra. Estas, una vez secadas en el laboratorio de suelos, se molieron con un mortero y se extrajo aproximadamente 5 g para ser enviado a la OIEA (Organización Internacional de Energía Atómica) para el respectivo análisis del porcentaje de nitrógeno total (%N-Total) y el porcentaje de nitrógeno derivado del fertilizante (%Nddf).
La toma de las muestras foliares de los diferentes tejidos para los diferentes estados fenológicos, se realizó de acuerdo al método del cuarteo (Segura 2003)5 cuando el estado fenológico de inicio (y) se desarrolló a F10, cuando el F10 pasó a Fm, cuando el Fm llegó a floración y cuando el estado floración pasó a cosecha. Se midió la altura de la planta madre, de la superficie del suelo hasta donde emerge la fruta y se cosechó la misma sin cortar el tallo y pesó el racimo de banano en el campo, luego se realizó un hueco de 1 m2 al frente del hijo de producción con el fin de muestrear las raíces de la madre, hijo de producción y del suelo. Se extrajo el hijo de producción entero, seccionando y muestreando el cormo y en suelo se seccionó el tallo y las hojas en cuatro partes. El tallo (vástago) de la madre se seccionó en tres partes y se pesó cada una, luego se muestreó una rodaja (“galleta”) de una pulgada de grosor de cada sección y se pesó en campo y en laboratorio. La fruta se pesó en su totalidad y se seccionaron todas las manos de banano y de cada una se tomó dos dedos con corona. Del pinzote se muestrearon tres secciones en medio y en ambos extremos. El cormo se seccionó en cuatro partes y de una de ellas se tomó una muestra para su evaluación. Por último, las hojas se cortaron, se pesaron y se estibaron y al azar se escogieron cuatro hojas, que se muestrearon en la vaina de abajo, en medio (vaina y lámina) y en la punta.
Para los estudios de seguimientos de la dinámica y recuperación del N, se analizaron el porcentaje y gramos del nitrógeno total (N-total) y del nitrógeno derivado del fertilizante (Nddf) enriquecido con 15N en el suelo y en la planta, en tejidos procedentes de las plantas madre e hijo. El enriquecimiento isotópico se determinó por espectrometría de masa en los laboratorios del Organismo Internacional de Energía Atómica (OIEA) en Seibersdorf, Austria (Zapata 1990, IAEA 2001).
La composición isotópica de nitrógeno, o sea, la relación 15N/N de cualquier material puede expresarse en átomos de porcentaje de abundancia de 15N, que se mide directamente por una determinación única mediante emisión óptica o espectrometría de masas. Dado que la composición isotópica de 15N en la atmósfera es aproximadamente 0,366% del N total, hay que restar esta cifra del porcentaje de átomos de abundancia de 15N de cualquier material enriquecido para obtener el porcentaje de átomos de 15N en exceso (% 15N a.e.). Entonces el nitrógeno derivado del fertilizante (Nddf) es la fracción de N en la planta o en el suelo que se deriva del fertilizante marcado con 15N, que se determina de la siguiente manera:

O como porcentaje

Por lo tanto, para calcular el porcentaje de nitrógeno derivado del fertilizante (% Nddf) es necesario determinar el % de átomos en exceso de 15N en las muestras vegetales o del suelo y en el fertilizante utilizado en el experimento.
Análisis de la información. Para el análisis de la información se utilizó el programa Anawin®6 el cual compara el efecto de las variables dependientes sobre las independientes y, a la vez compara y separa las medias con una probabilidad menor a p<0,05.
Resultados y Discusión
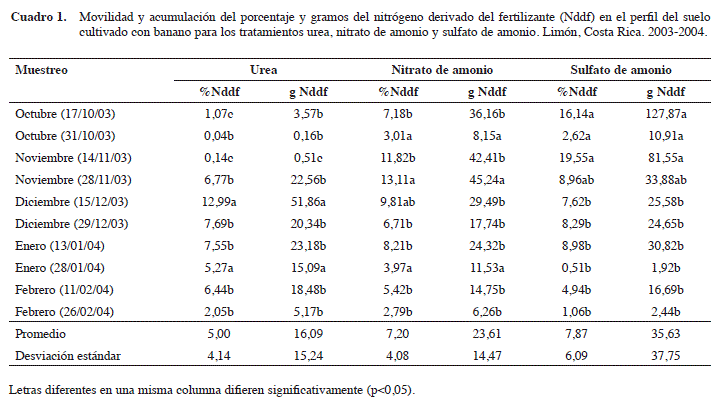
Movilidad y acumulación del nitrógeno derivado del fertilizante (Nddf) en el perfil del suelo procedente de los tratamientos de urea (U), nitrato de amonio (NA) y sulfato de amonio (SA). Hubo diferencias (p<0,05) en el porcentaje y gramos de Nddf producto de las fuentes nitrogenadas evaluadas y de las épocas de muestreo (Cuadro 1). Estas diferencias podrían estar influenciadas por las condiciones ambientales, como la temperatura y la precipitación diaria y acumulada durante la aplicación del fertilizante, así como por la humedad del suelo entre la aplicación y el muestreo. Ello debido a que las formas disponibles de N una vez en el suelo son susceptibles a la gasificación (denitrificación biológica y no biológica) y a lixiviación (nitrificación) (Prasertsak et al. 2001, Ramírez 2001, Bertsch 1998, Fassbender y Bornemisza 1994).
La variabilidad obtenida entre muestreos es alta y sin una tendencia establecida según sus valores promedios, probablemente por las variaciones climáticas (temperatura y lluvias), debidas a las condiciones de humedad del suelo durante el periodo de la investigación en el área estudiada. No obstante, se observó que en los meses de octubre y noviembre existe una menor acumulación de las formas disponibles de N procedentes de la urea, debido a una mayor tendencia de las pérdidas por los procesos de volatilización (NH3), fomentados por presentar estos suelos altos contenidos de humedad (de capacidad de campo a ligeramente saturados), pH pocos ácidos a neutros (6,5 a 6,8) y contenidos altos a medios (4,3 a 3,8%) de materia orgánica (MO) favorables a los procesos de pérdidas (Videla 1994, Bertsch 1998).
Los tratamientos presentaron una acumulación promedio en el perfil del suelo en orden descendente en gramos de N derivado del fertilizante (Nddf) de 35,63±37,7 de SA; 23,61±14,47 de NA y de 16,09±15,24 de urea. Esto permite concluir que su uso como fertilizante, es la forma de N más susceptible a perderse como gas del sistema suelo-planta, y se manifiesta como un menor aprovechamiento por parte de la planta. Las diferencias que se obtuvieron de la acumulación en el porcentaje de Nddf fluctuaron entre 0,67 y 2,87%, en este sentido, en orden descendente se registró en promedio al final del muestreo, sin considerar las fechas de muestreos mensuales, de 7,87±6,09% para el SA, de 7,20±4,08% para el NA y por último, el tratamiento con aplicación de U con 5,00±4,14%, lo que indica, que de forma similar, el N disponible procedente de la U se acumuló menos en el suelo, durante los meses de octubre y noviembre, por su susceptibilidad a perderse, por la volatilización del NH3, por la fijación del NH4+ entre las láminas de los minerales arcillosos del tipo 2:1 (esmécticas) que son predominantes en estos suelos (Arias 2009), por la rápida hidrólisis de la U a NH3 + CO2 + H2O por las adecuadas condiciones de humedad (de capacidad de campo a ligeramente saturados durante la investigación), coadyuvado por el pH ligeramente ácido a neutro (6,5 a 6,8) (Tisldale y Nelson 1970) en los horizontes superficiales del suelo y a los cambios de periodos lluviosos y de sequía entre muestreos.
Hubo diferencias (p<0,05) en la cantidad de Nddf acumulados de acuerdo con la profundidad de muestreo del perfil del suelo (Cuadro 2). Se determinaron diferencias significativas con respecto a las profundidades y al momento del muestreo. Las formas de N disponible obtenidas en porcentaje de Nddf, con valores promedios a las profundidades evaluadas de 0-10 cm; 10-30 cm, 30-60 cm y de 60-90 cm fueron de 13,80±7,66%; 4,33±2,51%; 4,53±4,36% y 4,10±2,92%, respectivamente. En cambio, para los gramos recuperados de Nddf, los valores obtenidos fueron de 35,53±22,75 g; 15,91±9,20 g; 29,50±40,49 g y 19,50±18,08 g, respectivamente.
En ambos casos, se notan dos rangos de profundidad donde se obtuvo una acumulación de estas fuentes disponibles de N, los rangos entre 0 a 10 cm (superficial) y entre 30 a 60 cm (subsuelo). En el primer caso, se podría asumir un efecto residual en la banda de fertilización aplicada de las formas de N disponible, por las formas amoniacales y se acentúan en los períodos de condiciones de alta humedad en el suelo y saturación superficial (mala aireación) asociados a los meses de mayor precipitación y alta temperatura, períodos de menor susceptibilidad a pérdidas por lixiviación (Fassbender y Bornemisza 1994, Bertsch 1998, Tisdale y Nelson 1970). En el segundo rango se pudo deber a una reducción en el movimiento del agua a través del perfil, por la disminución de la conductividad hidráulica y un aumento en la densidad aparente a esas profundidades.
En general, se obtiene un porcentaje de recuperación de Nddf de 26,76% en un rango de profundidad de 0 a 90 cm, en donde se registra que el 73,24% de N, sufre procesos de volatilización, lixivación, fijación y absorción interlaminares de las arcillas, o interacción de ellos sin que se pueda definir en este trabajo, el porcentaje de afectación de cada uno de estos procesos sobre la concentración de N en el suelo. A medida de referencia, Salifu et al. (2009) informa de 43% de recuperación de N en el suelo, sembrado de café, fertilizado con sulfato de amonio enriquecido con 5% a.e de 15N. Al compararlo con los resultados obtenidos en esta investigación, se obtiene un valor de 1,6 veces mayor en la recuperación del N, situación debida a las condiciones específicas de cada trabajo y a la fisiología de la planta.
Con respecto a la movilidad y acumulación de las formas de N disponibles por fuente y a las cuatro profundidades, se evidencia un comportamiento similar sobre la presencia de dos rangos de recuperación y acumulo de formas de N disponible para las fuentes de NA y SA, caso contrario con el tratamiento con U donde se nota un decrecimiento paulatino con la profundidad (Cuadro 2). En todos los rangos de profundidad evaluados, el tratamiento de SA presentó una recuperación mayor de formas de N disponibles, evaluadas por medio del porcentaje y los gramos del Nddf con respecto a los tratamientos de U y NA. En el rango de 30 a 60 cm de profundidad evaluado, se da un aumento en la acumulación de formas de N disponibles, esta acumulación y movilidad mayor procedente de SA, podría asociarse al efecto del ión acompañante (SO4-2) con respecto a los tratamientos de U y NA y asociado a la precipitación acumulada registrada desde el momento de la fertilización a cada muestreo. Otro aspecto a considerar es la estacionalidad de las formas disponibles de N a esa profundidad, debido a la tendencia de aumentar la microporosidad (densidad aparente: 0,996 g/cm3) asociados al desarrollo estructural bajo y a la permeabilidad lenta (1,8 a 1,6 cm/hora) de este subhorizonte (Acón 2002).
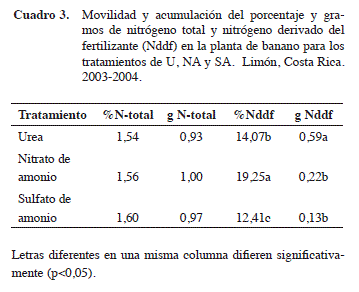
Movilidad y acumulación del nitrógeno total (N-total) y nitrógeno derivado del fertilizante (Nddf) en los diferentes tejidos del banano. Con respecto a la movilidad y acumulación del porcentaje y gramos de N-total y del Nddf a nivel de la planta de banano, no existieron diferencias con respecto al porcentaje y gramos de N-total a nivel de planta de banano entre los diferentes tratamientos evaluados en este trabajo (Cuadro 3). Las plantas de banano absorbieron y acumularon en cantidades similares formas de N disponibles, independiente de las fuentes empleadas.
No obstante, en cuanto al porcentaje de Nddf, a nivel de planta de banano se encontraron diferencias en la movilidad y acumulación, donde se favorece la acumulación del nitrógeno derivado del NA, debido a que se recuperó 1,37 y 1,55 veces más N que en los tratamientos fertilizados con U y SA, respectivamente. Al comparar la recuperación entre la U y el SA, este último es superado 1,13 veces por la U. Aunque en esta investigación no se valoró la concentración de formas de nitrógeno disponible, la diferencia obtenida en el porcentaje de recuperación podría deberse a la proporción entre el contenido de amonio (NH4+) y el nitrato (NO3-), debido a un sinergismo en el momento de la absorción del N por la planta (Ramírez 2001), en donde el NA presenta las dos formas de nitrógeno de forma más accesible, que las otras fuentes. Los datos obtenidos son similares a los informados por Prasertsak et al. (2001) para el porcentaje de recuperación de nitrógeno, donde reportan un valor de recuperación del 15% en relación con el nitrógeno aplicado.
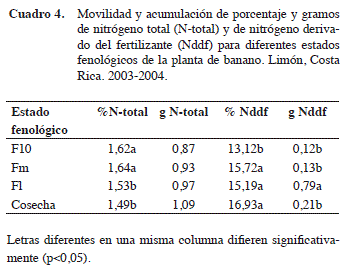
De acuerdo con la movilidad y acumulación del porcentaje y los gramos de N-total y Nddf para los diferentes estados fenológicos de la planta de banano (Y>F10; F10>Fm; Fm>Fl y Fl>cosecha), en el Cuadro 4 se observa que para el porcentaje de N-total, se obtuvo diferencias significativas en los estados fenológicos F10 (1,62%) y Fm (1,59%) con respecto a Fl (1,53%) y cosecha (1,49%); no obstante, en cuanto a los gramos de N-total entre los diferentes estados fenológicos no existieron diferencias significativas entre ellos.
Al comparar la información obtenida en esta investigación, con lo obtenido por Tavarez y Falquez (1997) en condiciones tropicales para el contenido de nitrógeno en los diferentes estados fenológicos: F10, FM, Fl y a cosecha, con valores de 1,68, 1,52, 1,53 y 1,34 % de N-total, respectivamente, se obtuvo un rango similar para la concentración de N. Cabe destacar que la reducción en el porcentaje de N-total y aumento de los gramos de N en la planta, podría deberse a un efecto de dilución del contenido de N con respecto a la biomasa total, debido a que durante las etapas de crecimiento de la planta del banano se puede pasar de un peso en base seca de 1 kg hasta 13 kg al culminar la cosecha.
También, se movilizó y acumuló más nitrógeno procedente del Nddf en los estados Fm (15,72%); Fl (15,19%) y cosecha (16,93%) (p≤0,05) con respecto al F10 (13,12%) (Cuadro 4). Los datos obtenidos son congruentes a lo informado por Prasertsak et al. (2001), para el porcentaje de recuperación.
Con respecto a la movilidad y acumulación del porcentaje y gramos de N-total y Nddf en los diferentes tejidos de la planta de banano conformada por la madre e hijo de producción (i), en el Cuadro 5, se observa que en el hijo de producción (i), el porcentaje de N-total es mayor con respecto a la planta madre, a saber, en los tejidos de hoja (i) (2,26%) y tallo (i) (2,10%), seguido de las hojas de la madre (1,94%). En cuanto al contenido en gramos, se observó que las hojas (1,56 g) y el rizoma (1,35 g) de la madre y el del hijo (1,27 g) se acumuló más con respecto a los otros tejidos, producto de la relación nitrógeno y peso seco de la planta(p≤0,05). Al analizar la unidad de producción como un todo (madre e hijo), se nota que del contenido total de nitrógeno de la planta, el 58,64% se acumula en la madre y el restante 41,36% en el hijo, lo que indica la importancia de cuidar ambas unidades para obtener una producción óptima en un corto y largo plazo.
En cuanto al porcentaje de Nddf, este se acumuló en mayor cantidad en las hojas (i) y tallo (i) del hijo de producción en 21,19% y 23,7%, respectivamente, (p<0,05), seguido del rizoma (i) del hijo (16,69%) y las raíces (16,46%) y pinzote (15,34%) de la madre. En cambio, no se obtuvieron diferencias significativas en el valor de gramos de N en ninguno de los tejidos evaluados.
Al analizar la interacción entre la movilidad y acumulación del porcentaje y gramos de N-total y Nddf en los tres tratamientos (U, NA y SA) con los cuatro estados fenológicos (F10, Fm, Fl y cosecha) (Cuadro 6), se observa que el porcentaje y los gramos de N-total no presentan diferencias significativas (p≥0,05) entre los diferentes estados fenológicos para cada tratamiento, ni tampoco para los diferentes estados fenológicos con igual tratamiento.
No obstante, para igual estado fenológico con diferentes tratamientos, se presentó diferencias significativas para el estado fenológico F10, donde la Urea (1,11 g) superó al NA y SA. Para el estado Fm, el tratamiento de SA (1,16 g) obtuvo un mayor valor con respecto a los demás tratamientos. En el caso del estado Fl, el tratamiento de SA (1,07 g) y al momento de la cosecha, con el tratamiento de NA (1,27 g) se encontraron diferencias significativas con respecto a los demás tratamientos en cada etapa fisiológica. Tal comportamiento, podría indicar la necesidad de implementar un programa de fertilización que intercale las diferentes fuentes empleadas en este trabajo, para obtener la concentración mayor de N en los tejidos de la planta, para optimizar la productividad de la plantación.
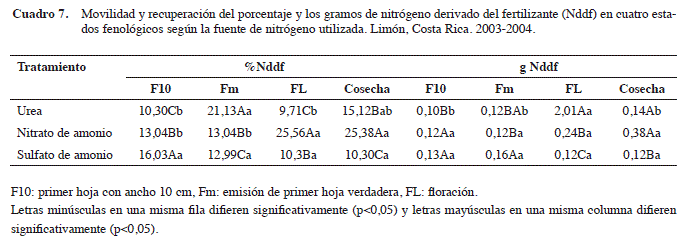
Al analizar el porcentanje y los gramos de Nddf (Cuadro 7), se observa que para los cuatro estados fenológicos, el tratamiento con U, presentó mayor concentración de N, donde el porcentaje de recuperación del Nddf en el estado Fm (21,13%) fue mayor con respecto a los demás estados fenológicos. No obstante, para el tratamiento con NA, se presentó una acumulación mayor (p<0,05) en los estados Fl (25,38%) y cosecha (25,38%) con respecto a los otros estados y con el tratamiento de SA, la fuente no afectó el porcentaje de recuperación del 15N.
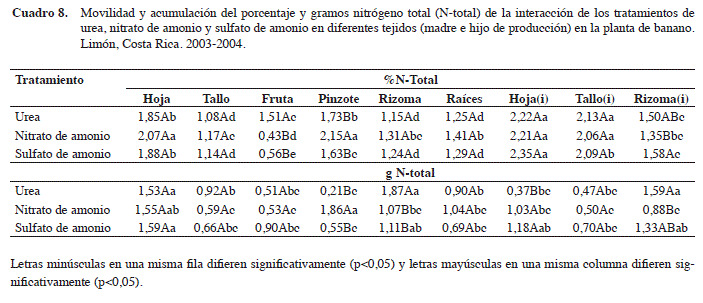
En el Cuadro 8 se observa que el porcentaje de N-total para los tratamientos de U, NA y SA presentó una acumulación mayor en los tejidos de las hojas(i) y tallo(i) del hijo de producción (p<0,05). En donde, se registró porcentajes de recuperación de 2,22% y 2,13% para el tratamiento con U, de 2,21% y 2,06% con el NA y de 2,35% y 2,09% para el tratamiento con SA, para los tejidos de la hoja y del tallo de la planta hijo, respectivamente. En la planta madre, el N se acumuló en las hojas y el tallo, a saber: para el tratamiento de U: 1,85 y 1,08%; de NA: 2,07 y 1,17% y en forma intermedia para el tratamiento de SA: 1,88 y 1,14 %, respectivamente.
En este sentido, los porcentajes promedios de recuperación del N-total en la biomasa total, según el tratamiento y la información obtenida, son de 1,60%, 1,57% y 1,53% para los tratamientos con U, NA y SA (Cuadro 8), lo que coincide con los valores determinados por tratamientos, según información generada anteriormente, en el Cuadro 5 para la planta total (U: 1,5%, NA: 1,56% y SA: 1,60%). En donde los valores totales recuperados de N-total en la biomasa total fueron de 14,42; 14,16 y 13,76 % para los tratamientos con U, NA y SA, respectivamente. Según la proporción, entre el N analizado en la planta madre como en la planta hijo, las relaciones determinadas en promedio fluctuaron entre 60:40 planta madre:planta hijo. Lo cual ratifica la importancia de ambas unidades para las producciones siguientes. En cuanto a los contenidos en gramos, para el tratamiento con U, el N se acumuló en forma significativa en los tejidos de la planta madre en las hojas (1,53 g), rizoma (1,87 g) y en el rizoma(i) del hijo de producción con 1,59 g.
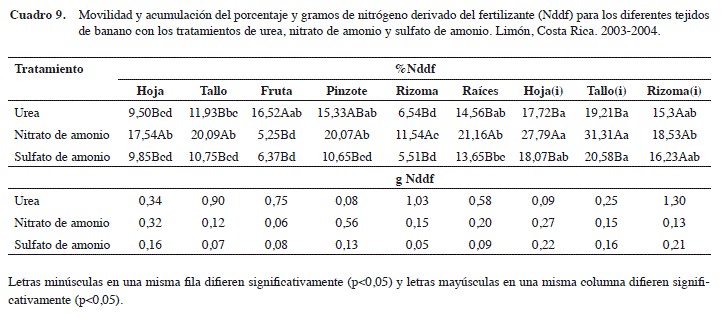
En el Cuadro 9, se observa que se presentó una acumulación mayor en todos los tratamiento con U, NA y SA en los tejidos de las hojas y tallo del hijo de sucesión, en donde, los tratamientos de U y SA presentaron para las hojas(i) y tallo(i) 17,72%; 19,21% y 18,07 y 20,58%, respectivamente, siendo el tratamiento de NA, en el cual se acumuló una mayor cantidad de N con: 27,79% y 31,31%, respectivamente; en cuanto a los gramos de Nddf, no evidenció ninguna diferencia significativa, por tratamiento ni por tejido. Con respecto a la acumulación del porcentaje y gramos de Nddf de igual tejido con diferentes tratamientos, el de NA respecto a los de U y SA presentó diferencias significativas en: hojas (17,54%); tallo (20,09%); pinzote (20,07%); rizoma (11,54%) de la planta madre y en la hoja(i) (27,79%); tallo(i) (31,31%) y rizoma(i) (18,53%) del hijo de sucesión. El tratamiento de U presentó una mayor acumulación en el porcentaje de Nddf en la fruta (16,52%) con respecto a los tratamientos de NA (5,25%) y SA (6,37%).
En los contenidos (gramos) de Nddf en la interacción entre los tratamientos U, NA y SA con los diferentes tejidos, no se presentaron diferencias significativas. Se observa que el tratamiento de U, en la fruta obtuvo 0,75 g en comparación con los de NA y SA, con 0,06 y 0,08 g, y los superó en 12,5 y 9,4 veces más de Nddf, respectivamente. En los tejidos de rizoma y raíces del tratamiento con U, se obtuvo una cantidad de Nddf de 1,03 y 0,58 g, respectivamente, con respecto a 0,15 y 0,05 g para el NA y de 0,20 y 0,09 g para el de SA, en ambos tejidos, respectivamente, lo que describe, una mayor acumulación de N en el tratamiento de U en el rizoma y raíces, de 6,9 a 5,2 y 11,6 a 6,4 veces más de Nddf que las otras dos fuentes. Tal comportamiento podría tener una explicación en la nutrición de planta, debido a que la formas de N disponibles: el nitrato (NO3-) y amonio (NH4+) deberían ser entendidos como dos nutrientes diferentes debido a su comportamiento dentro de la planta, donde se relaciona el efecto del NH4+ con lo verdoso de esta, mientras el NO3- produce crecimiento o vigor de las plantas (Mengel y Kirbby 1982, Mills y Jones 1996, Ramírez 2001).
Sin embargo, dentro de la planta, el NH4+ es tóxico y debe combinarse con carbono para formar compuestos orgánicos nitrogenados; esta utilización del carbono retarda el crecimiento y por ende, el peso seco de las plantas. El primer efecto del amonio es la reducción del crecimiento de raíces por el metabolismo del carbono, por consiguiente, menor biomasa al combinarse con el carbono en las raíces y ante la falta de carbono, el amonio se va acumulando (Mengel y Kirbby 1982, Mills y Jones 1996 y Ramírez 2001). Este efecto parece evidenciarse al analizar el porcentaje y gramos de Nddf (Cuadro 9) en el tejido del rizoma, donde los contenidos del porcentaje de Nddf entre la U (6,54%) y SA (5,51%) son similares; sin embargo, los gramos de Nddf son bajos para el SA (0,05 g), lo que influyó en el poco crecimiento y el bajo peso seco del rizoma. Al comparar el porcentaje de Nddf en el rizoma entre el tratamiento de U (6,54%) y el SA (5,51%), el primero lo supera en 1,18 veces, mientras que los gramos de Nddf es veinte veces mayor en la U (1,03 g) que el SA (0,05 g); esto denota la diferencia en el peso del rizoma que se obtuvo en esta investigación entre la U (1,03 g) y el NA (0,15 g), lo que evidencia el efecto negativo del aumento del amonio sobre el rizoma y en las raíces.
El mismo efecto se evidenció en el rizoma del hijo, donde a pesar de que el porcentaje de Nddf del SA es mayor que la U, los gramos de Nddf del SA es seis veces menor. Este efecto es evidente también en los tallos de la madre y el hijo de sucesión y en la hoja de la planta madre.
Literatura citada
Acón, J. 2002. Estudio detallado de suelos y clasificación de las tierras según su aptitud para el cultivo de banano de la finca perteneciente a la compañía Hacienda Río Palacio S.A., situada en Venecia y Saborío, distrito Carrandí, Cantón Matina, Provincia de Limón. Instituto de Desarrollo Agrario (IDA). Costa Rica. 84 p. [ Links ]
Arias, F. 2009 Caracterización química y mineralógica de suelos cultivos en banano (Musa AAA) en la zona Caribe de Costa Rica. Tesis Mag. Sc., Sistema de Estudios de Posgrado, Universidad de Costa Rica. San José, Costa Rica 135 p. [ Links ]
Bertsch, F. 1998. La fertilidad de los suelos y su manejo. Asociación Costarricense de la Ciencia del Suelo (ACCS). San José, Costa Rica. 57 p. [ Links ]
Elizondo, J. 2006. El nitrógeno en los sistemas ganaderos de leche. Agronomía Mesoamericana 17(1):69-77. [ Links ]
Fassbender, H; Bornemisza, E. 1994. Química de suelos con énfasis en suelos de América Latina. IICA. San José, Costa Rica. 420 p. [ Links ]
IAEA (International Atomic Energy Agency). 2001. Manual use of isotope and radiation methods in soil and water management and crop nutrition. Training course series 14. Vienna, Austria. 247 p. [ Links ]
López, A; Espinosa J. 1995. Manual de nutrición y fertilización del banano. Instituto de la Potasa y el Fósforo. Quito, Ecuador. 82 p. [ Links ]
Mengel, K; Kirbby, E. 1982. Principles of plant nutrition. International Potash Institut. Warbleufen-Bern. Suitzenland. 687 p. [ Links ]
Mills, HA; Jones, JB. 1996. Plant analysis handbook II. A practical sampling, preparation, analysis, and interpretation guide. Micromacro Publishing. Athens, Greece. 422 p. [ Links ]
Ortiz, R; López, A; Ponchner, S; Segura, A. 1999. El cultivo del banano. EUNED. San José, Costa Rica. 186 p. [ Links ]
Prasertsak, P; Freney, JR; Saffigna, PG; Denmead, OT; Prove, BG. 2001. Fate of urea nitrogen applied to a banana crop in the wet tropics of Queensland. Nutrient Cycling in Agroecosystems 59:65-73. [ Links ]
Ramírez, C. 2001. Nutrición nitrogenada de las plantas. In Villalobos, E. ed. Fisiología de la producción de los cultivos tropicales. Editorial de la Universidad de Costa Rica. San José, Costa Rica. p. 198-226. [ Links ]
Salifu, K; Islam, M; Jacobs, D. 2009. Retranslocation, plant and soil recovery of nitrogen-15 applied to bareroot black walnut seedlings. Communications in soil science and plant analysis 40:1408-1417. [ Links ]
Soto, M. 1995. Bananos: cultivo y comercialización. 2 ed. Litografía e Imprenta Lil. San José, Costa Rica. 649 p. [ Links ]
Tavares, G; Falquez, C. 1997. Contenido nutricional y absorción de nutrientes en plantas de banano (Musa AAA), subgrupo Cavendish, clon “Gran Enano”, en diferentes etapas fenológicas de desarrollo. Trabajo de Graduación. EARTH. Guácimo, Limón, Costa Rica. 77 p. [ Links ]
Tisdale, SL; Nelson, WL. 1970. Fertilidad de los suelos y fertilizantes. Ed. Montaner y Simonn S.A. Barcelona, España. 760 p. [ Links ]
USDA (United States Department of Agriculture). 1998. Keys to soil taxonomy. Agriculture Handbook No 436. Natural Resources Conservation Service. Soil Survey Staff. USA. 326 p. [ Links ]
Videla, C. 1994. La volatilización de amoníaco: una vía de pérdida de nitrógeno en sistemas agropecuarios. Boletín Técnico 131. EEA INTA Balcarce. Buenos Aires, Argentina. 16 p. [ Links ]
Weisenburger, D. 1991. Potential health consequences of ground-water contamination by nitrates in Nebraska. In Bogardi, I; Kuzelka, RD. eds. Nitrate contamination. Springer-Verlag. Berlin, Germany. p. 309-315. [ Links ]
Zapata, 1990. Técnicas isotópicas en estudios sobre la fertilidad del suelo y la nutrición de las plantas. In Hardason, G. ed. Empleo de técnicas nucleares en los estudios de la relación suelo-planta. Colección de Cursos de Capacitación Nº 2. Organismo Internacional de Energía Atómica. Viena, Austria. p. 79-171. [ Links ]
*Correspondencia a:
Jorge Acón-Ho. Programa de Estudios de Posgrado en Ciencias Agrícolas y Recursos Naturales. Doctorado en Sistema de Producción Agrícola Tropical Sostenible. Universidad de Costa Rica. San José, Costa Rica. jorge.acon@ice.co.cr
Carlos Cervantes-Umaña. Consultor de Suelos. Heredia, Costa Rica. ccervant.53t@hotmail.com
Rodolfo WingChing-Jones. Escuela de Zootecnia, Centro Investigación en Nutrición Animal. Universidad de Costa Rica. San José, Costa Rica. rodolfo.wingching@ucr.ac.cr
1. Trabajo de graduación del primer autor. Sistema de Estudios de Posgrado de la Universidad de Costa Rica, Costa Rica.
2. Programa de Estudios de Posgrado en Ciencias Agrícolas y Recursos Naturales. Doctorado en Sistema de Producción Agrícola Tropical Sostenible. Universidad de Costa Rica. San José, Costa Rica. jorge.acon@ice.co.cr
3. Consultor de Suelos. Heredia, Costa Rica. ccervant.53t@hotmail.com
4. Escuela de Zootecnia, Centro Investigación en Nutrición Animal. Universidad de Costa Rica. San José, Costa Rica. rodolfo.wingching@ucr.ac.cr
Recibido: 18 de julio, 2012. Aceptado: 1 de abril, 2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}