










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgronomía Mesoamericana
versión On-line ISSN 2215-3608versión impresa ISSN 1659-1321
Agron. Mesoam vol.24 no.1 San Pedro jun. 2013
Nematodos fitoparásitos asociados al tomate en la zona occidental de Nicaragua
Parasitic nematodes associated to tomato plants in western Nicaragua
Parasitic nematodes associated to tomato plants in western Nicaragua
*Dirección para correspondencia:
Resumen
El objetivo de este estudio fue identificar la diversidad y dinámica poblacional de los nematodos fitoparásitos asociados al tomate en los departamentos de León y Chinandega pertenecientes al occidente de Nicaragua. Durante el ciclo agrícola 2010-2011 se muestrearon cinco plantaciones en cada departamento. Se realizaron ocho muestreos en cada plantación para un total de 80 muestras en las diez plantaciones seleccionadas. Estas fueron tomadas a 10 m entre sí en forma de zig-zag. Se extrajeron los nematodos de suelo mediante el método de Baermann modificado y extracción de nematodos de raíces mediante el método de macerado y tamizado de raíces. Los datos obtenidos fueron correlacionados con factores que afectan las poblaciones de nematodos, tales como tipo de suelo, rotación de cultivos y precipitaciones. Igualmente se describió el crecimiento poblacional de los principales géneros mediante modelos matemáticos. Las densidades más altas fueron obtenidas de muestras de suelo, los más frecuentes en 100 g de suelo fueron especies no identificadas de Meloidogyne sp. con 739, Pratylenchus con 555, Tylenchorhynchus 386 y Helicotylenchus 252 individuos. Se evidenció también que el factor precipitaciones no tuvo un efecto significativo sobre las poblaciones de nematodos. Finalmente, Meloidogyne y Pratylenchus presentaron un crecimiento poblacional que se ajusta a un modelo matemático lineal (R2=0,98 para ambos géneros), donde existió una correlación positiva y lineal entre las etapas fenológicas del tomate y el incremento de nematodos en el suelo.
Palabras clave: dinámica poblacional, Meloidogyne, Pratylenchus, rotación de cultivos, tipos de suelos.
Abstract
The objective of this study was to identify parasitic nematodes associated with the tomato crop in Leon and Chinandega western region of Nicaragua. During the period of 2010-2011, five tomato fields were sampled in both Leon and Chinandega; these samples included soil and roots, the total sampled area was of 2.5 ha. Eight samples were collected from each of the ten tomato plantations studied, for a total of 80 samples. Soil and root samples were taken 10 m from each other in a zig-zag pattern. Soil nematode extraction was conducted using the Modified Baermann funnel technique and maceration and sieving method was used for root nematodes. The data obtained was correlated with predisposing factors that have been reported to influence nematodes such as soil type, crop rotation and precipitation. Similarly, the population growth of the main nematode genera was described through mathematical models. The greatest densities of nematodes were isolated from soil samples; the most common nematodes in 100 grams of soil were Meloidogyne with 739, Pratylenchus with 555, Tylenchorhynchus with 386 and Helicotylenchus with 252. It was proven that precipitation had no significant effect on the parasitic nematode population. Finally, it was determined that the increasing number of Meloidogyne and Pratylenchus in the tomato crop fit a linear mathematical model (R2=0,98 for both nematodes), showing an existing positive and linear correlation between the phenological stages of the tomato and the increment of the nematodes in the soil.
Key words: population dynamics, Meloidogyne, Pratylenchus, crop rotations, soil types.
Introducción
La producción de tomate (Solanum lycopersicum L.) en Nicaragua es muy importante debido a su alto contenido nutricional y buena aceptación en el mercado nacional. Según la FAO, en el año 2010 Nicaragua produjo 5900 toneladas de tomate en 400 hectáreas para un promedio de 14,76 t/ha. Esto representa un aumento del 100% en el rendimiento en comparación con lo reportado en el año 2006, cuando se obtuvo promedios de 7,30 t/ha. Sin embargo, a pesar de este incremento en la producción, Nicaragua es en la actualidad el país con menores rendimientos en el área centroamericana (FAO 2010).
La zona occidental de Nicaragua, que comprende León y Chinandega, no se ha caracterizado por ser una zona productora de hortalizas, sin embargo en los últimos años la producción de tomate ha tomado auge como parte de un proceso de diversificación de las fincas de pequeños y medianos agricultores (Salazar et al. 2009), demostrando la importancia que este rubro representa para esta zona agrícola.
Uno de los principales factores limitantes en la producción de tomate lo constituyen los nematodos fitoparásitos. Estos organismos generan pérdidas que han sido estimadas en 20% (Sasser y Freckman 1987, Sasser 1989, Shurtleff y Averre III 2000). Los principales nematodos asociados al cultivo del tomate son los géneros Meloidogyne sp. (Tzortzakakis 2000, Karssen y Moens 2006), Pratylenchus sp. (Blancard 2011) y Tylenchorhynchus sp. y Helicotylenchus sp., (Overman 1999, Shurtleff y Averre III 2000).
Estudios recientes indican que la capacidad de causar daño de estos nematodos se ve favorecida por factores edafoclimáticos y agronómicos como, monocultivo (Chen y Tsay 2006), altas precipitaciones (Masse et al. 2004) y suelos arenosos (Starr et al. 1993). A pesar de la importancia de estos factores, los estudios nematológicos en Nicaragua son muy pocos y los que se han realizado, han estado enfocados principalmente a nematodos fitoparásitos asociados al café (Herrera y Marbán-Mendoza 1999, Herrera et al. 2011). Mientras que los estudios dirigidos a cultivos hortícolas son muy escasos (Martínez y Salazar 2007, Salazar 2009), y no están disponibles para la mayoría de agricultores y profesionales agrícolas.
El presente estudio tuvo como objetivo identificar la diversidad y dinámica poblacional de los nematodos fitoparásitos asociados a tomate y los factores agroclimáticos que influyen en sus poblaciones en León y Chinandega, Nicaragua.
Materiales y Métodos
Ubicación del estudio
El estudio se realizó durante el periodo 2010-2011 en la región occidental de Nicaragua, departamentos de León y Chinandega. El clima de la zona es tropical, con temperaturas que varían entre 26,1 y 36,3oC, precipitación de 1388 a 1702 mm anuales y una altura entre 130-200 msnm (MAGFOR 2000).
Muestreo de nematodos fitoparásitos
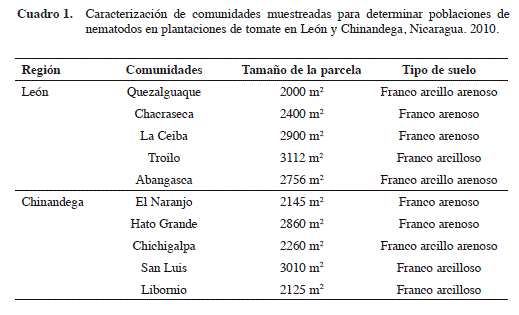
El muestreo se realizó en diez plantaciones de tomate, cinco ubicadas en el departamento de León y cinco en el departamento de Chinandega (Cuadro 1). Las plantaciones seleccionadas tenían sembrado tomate variedad Shanty, una de las más utilizadas por los productores en el occidente de Nicaragua.
En total se muestrearon 2,5 ha, con plantaciones de tomate que variaron entre 2000 y 3112 m2. Se realizaron cuatro muestreos en cada plantación, el primero a los 27 días después de la germinación (DDG) (al trasplante); el segundo a los 50 DDG (floración); el tercero a los 85 DDG (inicio de la fructificación) y el cuarto a los 120 DDG (fructificación plena). En cada una de las 10 plantaciones estudiadas se realizaron cuatro muestreos de suelo y cuatro de raíces para un total de 80 muestras evaluadas.
Cada muestra estaba constituida por 10 submuestras según recomendación de Shurtleff y Averre III (2000), las cuales fueron tomadas a una distancia de 10 m entre sí en forma de zig-zag. En cada punto de muestreo se extrajeron raíces y suelo de la rizosfera de la planta a una profundidad de 30 cm con la ayuda de un barreno metálico. Las submuestras fueron mezcladas homogéneamente y de ellas se tomó una muestra de 454 g de suelo y 25 g de raíces representativo de la plantación. Las muestras de suelo y de raíz fueron empacadas en bolsas plásticas, rotuladas y enviadas al Laboratorio de Fitopatología de la UNAN-León donde se realizaron los análisis.
Análisis de suelo, nematodos y datos pluviométricos
Análisis de suelo: antes de establecer el cultivo, se tomaron muestras de suelo, determinándose la textura y porcentajes de arcilla, limo y arena mediante el método de Bouyoucos (Bouyoucos 1936).
Extracción de nematodos de suelo y raíces: la extracción de nematodos de suelo se realizó mediante el método de Baermann modificado (Hooper et al. 2005). Se pesaron 100 g de suelo y se colocaron en un filtro de papel toalla que estaba extendido sobre un tamiz dentro de un recipiente. Luego se agregó agua hasta alcanzar humedecer el nivel superior del suelo sin saturarlo. Se incubó por 48 horas y luego se filtró la solución dos veces por un tamiz de 400 mesh.
La extracción de nematodos de raíces se realizó mediante el método de macerado y filtrado (Hooper et al. 2005). Se pesaron 25 g de raíces y se colocaron en una licuadora con 200 ml de agua, seguidamente se licuó por 30 segundos. La solución se filtró a través de tamices de 200 y 400 mesh, luego con la ayuda de una piceta los nematodos retenidos fueron transferidos a un beaker hasta completar 20 ml. La identificación de nematodos se realizó por parámetros morfométricos y la clave de identificación de nematodos diseñada por Mai y Mullin (1996).
Para la cuantificación de nematodos se tomó una alícuota de 1 ml de la solución con nematodos; este procedimiento se repitió cinco veces para obtener el valor promedio.
Análisis pluviométricos: para determinar la influencia de la precipitación sobre las poblaciones de nematodos, se colocó pluviómetros en las zonas bajo estudio y se calculó el índice de precipitación media para los meses septiembre-noviembre.
Análisis estadístico
Se evaluaron cuatros factores y su incidencia sobre poblaciones de nematodos. Los factores fueron, cuatro etapas fenológicas (trasplante, floración, inicio de la fructificación y fructificación plena), tres tipos de textura de suelo (franco arcilloso, franco arcillo arenoso y franco arenoso), tres tipos de rotación (sin rotación, de ciclo de por medio y por más de dos ciclos) y precipitaciones anuales (≤700 mm, 700-1000 mm y ≥1000 mm). Los datos fueron procesados mediante un análisis de varianza (ANDEVA) y un test de separación de medias según Duncan al 0,05% de confianza. Se realizaron regresiones lineales simples para determinar la correlación entre la población de nematodos y las etapas fenológicas del tomate, con el propósito de establecer un modelo matemático. Adicionalmente se realizó correlaciones de Pearson con el fin de medir el grado de covarianza entre la población de nematodos y las etapas fenológicas.
Resultados y Discusión
Nematodos fitoparásitos identificados en la zona occidental de Nicaragua
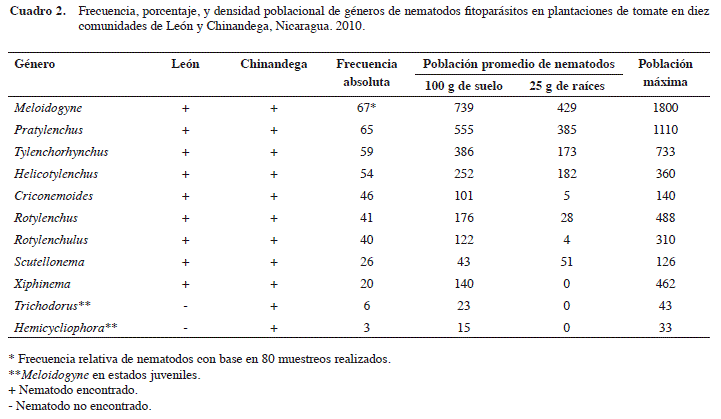
Se encontraron en total once géneros de nematodos fitoparásitos asociados al tomate. Los géneros Trichodorus y Hemicycliophora solamente fueron encontrados en comunidades del Departamento de Chinandega (Cuadro 2).
Los géneros nematodos más frecuentes fueron Meloidogyne y Pratylenchus, en 67 y 65 muestras (Cuadro 2). Estos dos géneros representan el 31% de los nematodos fitoparásitos encontrados. Los géneros Tylenchorhynchus y Helicotylenchus fueron aislados de 59 y 54 muestras respectivamente y representan el 14% y 3% del total de nematodos fitoparásitos encontrados.
Meloidogyne presentó las poblacionales más altas en el estudio con 739 juveniles de segundo estadio en 100 g de suelo y la densidad poblacional máxima con 1800 individuos (Cuadro 2). Este género es muy importante (Agrios 1997) ya que causa en tomate hasta un 90% de los daños por nematodos. Similarmente se ha reportado que es el principal nematodo fitoparásito asociado al tomate durante sus diferentes etapas fenológicas, su importancia se basa en su alta capacidad para dañar las raíces secundarias de las plántulas de tomate reduciendo su capacidad de alimentación y desarrollo (Tzortzakakis 2000, Karssen y Moens 2006). Pratylenchus fue el segundo género más abundante con un promedio de 555 individuos en 100 g de suelo y una densidad poblacional máxima de 1110 (Cuadro 2). Este nematodo tiene una amplia distribución geográfica y causa daños de importancia económica en el cultivo del tomate (Blancard 2011, Perdomo 1990). Este nematodo es de mucha importancia ya que causa serias pérdidas al tomate; sin embargo, este problema es poco conocido lo que hace que usualmente sean subestimados a la hora de establecer medidas de control (Blancard 2011).
Las poblaciones más altas de Tylenchorhynchus y Helicotylenchus fueron encontradas en muestras de suelo de 100 g cada una (Cuadro 2), el primero presentó 386 individuos y el segundo 265 individuos. Estos dos géneros han sido reportados previamente en el tomate (Overman 1999, Shurtleff y Averre III 2000), lo que concuerda con los datos aquí presentados, que indican una amplia distribución de estos dos géneros en la zona occidental de Nicaragua. Se consideran de poca importancia debido a que no han sido asociados a pérdidas económicas considerables en el cultivo de tomate (Perdomo 1990), no obstante, su potencialidad como nematodos de importancia económica es latente, puesto que están distribuidos en todas las zonas bajo estudio. Fernández-Solano y Quesada-Solís (2009) coinciden con lo indicado en el presente estudio para Costa Rica, sobre la presencia de Helicotylenchus asociado al cultivo del tomate, no así para el caso de Tylenchorhynchus, el cual para el año 2009 no estaba reportado en dicho cultivo.
Otros géneros que se encontraron en el estudio en 100 g de suelo fueron Rotylenchus (176), Rotylenchulus (141), Xiphinema (140), y Criconemoides (116). Rotylenchus y Xiphinema, aunque han sido reportados como nematodos fitoparásitos asociados al cultivo del tomate (Overman 1999, Bitterlin y Gonsalves 1987), no se encontraron en poblaciones que representen un riesgo para el cultivo. En particular, el género Xiphinema ha sido reportado como transmisor del virus de la mancha anular del tomate (Overman 1999), lo que lo convierte en un nematodo potencialmente importante en este cultivo.
Rotylenchulus ha sido encontrado asociado al cultivo del tomate (Khan et al. 1985), sin embargo sus densidades generalmente son bajas y con poca incidencia en el tomate (Shurtleff y Averre III 2000). En el caso de Criconemoides fue reportado en tomate en grandes poblaciones y con una alta frecuencia en España (Bello y Lara 1986). Sin embargo, en el presente estudio la frecuencia de aislamiento de este género en la zona occidental de Nicaragua fue baja en comparación con los resultados antes descritos.
Los géneros Scutellonema, Trichodorus y Hemicycliophora fueron encontrados en bajas poblaciones en todas las zonas evaluadas. Scutellonema se encontró asociado a tomate, con una frecuencia de 26 y densidad máxima de 126 individuos. Su promedio más alto fue de 51 individuos encontrados en 25 g de raíces de tomate.
Los nematodos Trichodorus y Hemicycliophora fueron identificados como los géneros menos frecuentes, ya que se encontraron solamente seis y tres veces en las ochenta muestras, respectivamente. El promedio más alto para el género Trichodorus fue de veintitrés individuos, en 100 g de suelo, y la densidad máxima fue de 43 nematodos. En el caso de Hemicycliophora, el promedio más alto fue de quince nematodos en 100 g de suelo y su densidad máxima 33.
Influencia de factores edafoclimáticos y agronómicos sobre poblaciones de nematodos fitoparásitos asociados a tomate en la zona occidental de Nicaragua
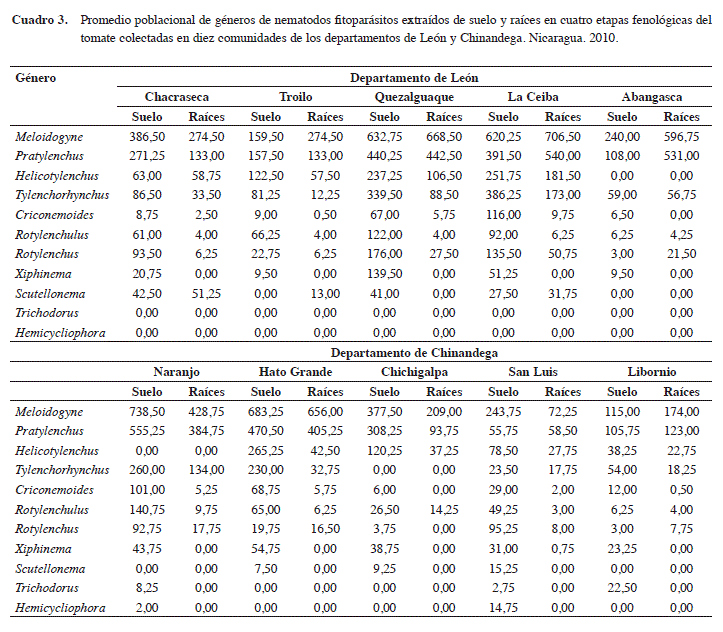
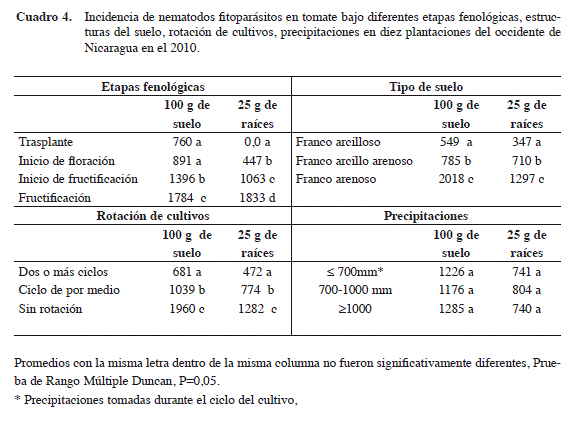
Tipo de suelo y su efecto sobre poblaciones de nematodos: los niveles poblacionales más altos de nematodos fitoparásitos fueron observadas en las comunidades La Ceiba, Hato Grande y El Naranjo (Cuadro 3). Estas comunidades tienen en común que poseen suelos de tipo franco-arenoso, el cual favorece el desarrollo de los nematodos. El promedio de estos en suelos franco arenosos (2018 en suelo y 1297 en raíces) fue significativamente más alto (P≤0,05) que el encontrado en suelos franco arcillosos (549 en suelo y 347 en raíces) (Cuadro 4).
La textura es un factor que influye significativamente en la diversidad y niveles poblacionales de microorganismos en el suelo (Buyer et al. 2002, Cavigelli et al. 2005). El tipo de suelo afecta la distribución de los nematodos, debido a que características como la textura, son muy estables y afectan la tasa reproductiva y los niveles poblacionales de los nematodos en la rizósfera (Windham y Barker 1986). Los suelos con alto contenido de arena constituyen un ambiente ideal para que los nematodos fitoparásitos puedan moverse y conseguir su alimento, lo que favorece su tasa reproductiva y su desarrollo (Windham y Barker 1986, Robinson et al. 1987). Igualmente, existe una asociación positiva entre altas densidades de nematodos y suelos con un alto contenido de arena (Starr et al. 1993).
Los promedios poblacionales más bajos de nematodos (P≤ 0,05) se encontraron en San Luis y Libornio, ubicadas en el departamento de Chinandega (Cuadro 3). Estas localidades se caracterizan por poseer suelos con altos contenidos de arcilla (franco-arcilloso) y poca porosidad en su estructura, lo que afecta el incremento de las poblaciones de nematodos, debido a que dificultan su acceso a fuentes de alimentación y su reproducción (Griffiths y Caul 1993).
Tipo de rotación y su efecto sobre poblaciones de nematodos: en relación con los tipos de rotación, las mayores poblaciones de nematodos (P≤0,05) fueron encontradas en plantaciones utilizadas continuamente para la producción de tomate. En estas, se obtuvieron promedios poblacionales de 1960 y 1282 individuos en suelo y raíces (Cuadro 4). La ausencia de rotación de cultivos favoreció el incremento del número de nematodos en el suelo al mantenerse el mismo hospedante por largos periodos de tiempo (Chen y Tsay 2006).
Las densidades de nematodos en plantaciones con rotación de cultivos de dos o más ciclos utilizando gramíneas fueron más bajas (P≤0,05) que las encontradas en comunidades sin rotación. Los promedios en plantaciones con rotación de cultivos fueron de 681 y 472 individuos en suelos y raíces, en comparación con 1960 y 1282 en suelo y raíces que se obtuvo en suelos sin rotación. Johnson (1985) afirma que la rotación de cultivos reduce el daño causado por nematodos a los cultivos subsiguientes al modificar los nichos ecológicos en los que estos organismos se desarrollan y reproducen, generando condiciones de suelo que reducen su capacidad de penetrar e infectar las plantas. Meloidogyne y Pratylenchus han sido reportados como susceptibles a las rotaciones de cultivos, por lo que sufren reducciones significativas en sus poblaciones (Chen y Tsay 2006). Esto se ha logrado, sin incrementar la población de otras especies de nematodos parasíticos para el cultivo subsiguiente a la rotación.
Etapas fenológicas y su efecto sobre poblaciones de nematodos
El desarrollo de las plantas de tomate y su paso por sus diferentes etapas fenológicas influyó en el incremento poblacional de nematodos fitoparásitos. Se evidenció un incremento significativo (P≤ 0,05) del número de nematodos al comparar las poblaciones al momento del trasplante (760 nematodos) y al momento de la fructificación (1784) (Cuadro 4).
Este comportamiento se debe a que los nematodos fitoparásitos necesitan, para completar su ciclo de vida, una relación estrecha con su hospedero el cual le provee alimento y condiciones favorables para su desarrollo (Cadet et al. 2005). Las comunidades de nematodos evolucionan paralelamente a los cambios que sufren las comunidades de plantas a las que están asociados (Cadet et al. 2005). Esta dependencia que muestran los nematodos a los cambios en sus hospederos ha sido estudiada previamente. Masse et al. (2004) indican que esta relación de dependencia entre planta y patógeno es predecible puesto que la presión de cambio en la vegetación hospedera afecta a los organismos vivos que dependen de ella.
Precipitaciones y su efecto sobre poblaciones de nematodos: al comparar las poblaciones de nematodos asociados a tomate con precipitaciones de menos de 700 mm, entre 700-1000 mm y más de 1000 mm, se observó, que los promedios poblacionales no fueron significativamente diferentes (P≥0,05).
A pesar de que las precipitaciones han sido consideradas como un factor que favorece el incremento poblacional de los nematodos en el suelo (Haque y Mukhopadhyaya 1982, Mase et al. 2005), en el presente estudio no influyeron de manera significativa sobre los cambios poblacionales de los nematodos. La tasa poblacional de estos no presentó diferencias significativas en los tres niveles de precipitaciones analizadas (Cuadro 4). Esto podría estar relacionado al hecho que las plantaciones de tomate poseían sistemas de riego para complementar la humedad generada por las lluvias, lo que implica que las condiciones de humedad fueron constantes todo el tiempo.
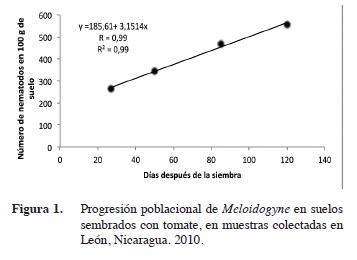
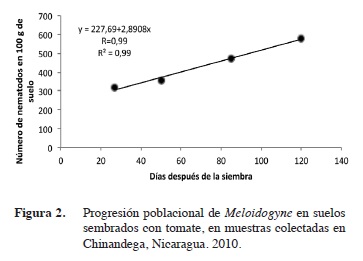
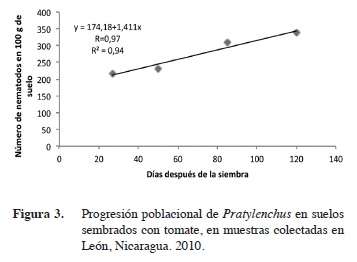
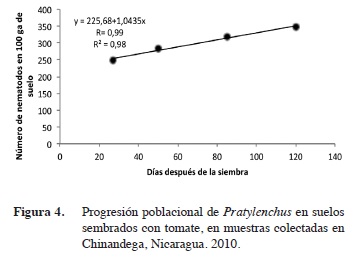
Dinámica poblacional de Pratylenchus y Meloidogyne
La regresión lineal simple realizada a Meloidogyne y Pratylenchus, versus las etapas fenológicas del cultivo, indica que ambos nematodos presentaron una dinámica poblacional similar.
Las poblaciones de Meloidogyne, al correlacionarse con las etapas fenológicas presentaron un alto coeficiente de correlación de R= 0,99 en León y Chinandega (Figuras 1 y 2); esto demuestra una relación de dependencia entre ambas variables. El coeficiente de determinación fue igualmente alto con un R2= 0,99. Las poblaciones de Pratylenchus, presentaron similarmente una alto coeficiente de correlación (R= 0,97 y 0,99) en León y Chinandega (Figuras 3 y 4). Estos datos prueban que existe una fuerte relación entre el número de nematodos y las etapas de crecimiento del tomate. Los coeficientes de determinación de Pratylenchus sp. fueron altos con una R2= 0,95 y R2=0,98 (Figuras 3 y 4) en León y Chinandega. El análisis de correlación de Pearson realizado a las poblaciones de Meloidogyne y Pratylenchus en León y Chinandega presentan una correlación igual a 0,99 excepto en el caso de Pratylenchus en León, donde se obtuvo un coeficiente de correlación de Pearson de 0,97, con una significancia estadística que varió entre 0,02-0,23 menos que 0,05 por lo que se puede afirmar que las poblaciones de nematodos están correlacionadas linealmente con las etapas fenológicas del cultivo del tomate.
Ambos géneros de nematodos presentaron coeficientes de determinación superiores a 0,90 lo que significa que el 90% de la variación en sus poblaciones está explicada por las diferentes etapas de desarrollo del tomate.
Este tipo de crecimiento poblacional puede explicarse con una función de regresión lineal, por lo que el modelo matemático que mejor se ajusta al crecimiento poblacional de estos nematodos fue el lineal. Este modelo de crecimiento ya ha sido observado para poblaciones de nematodos como Pratylenchus en el cultivo del arroz en Costa Rica (Guzmán-Hernández et al. 2011). De igual forma, este tipo de crecimiento poblacional ha sido reportado por González et al. (2007), los que encontraron que poblaciones del género Radopholus asociadas al cultivo del plátano se incrementaron consistentemente desde el inicio hasta la senescencia de la planta, llegando a obtener poblaciones de 40 000 individuos lo que se considera extremadamente alto. Este comportamiento también coincide con lo encontrado en el presente estudio y se debe a la alta dependencia que los nematodos endoparásitos muestran en relación a sus hospederos (Cadet et al. 2005), lo que hace que incrementen sus poblaciones proporcionalmente a la disponibilidad de alimentos que sus hospederos ofrecen. Este crecimiento lineal se ve influenciado también por el hecho de que plantas con sistemas radiculares extensos, como es propio de plantas adultas, proveen más alimentos y refugio a los nematodos que plantas jóvenes.
Otro factor a considerar es que, en el caso del cultivo del plátano, raíces de plantas jóvenes son menos atacadas por nematodos debido a su estado de desarrollo y función en términos de absorción de nutrimentos para la planta (Castrillón y Castrillón 2000). En cambio raíces de plantas adultas o en época de cosecha pierden esta funcionalidad, se debilitan y se hacen más susceptibles al ataque de nematodos (Gómez 1983, Castrillón y Castrillón 2000). Esto podría explicar el crecimiento lineal que estos nematodos presentaron en el cultivo del tomate, donde las últimas etapas fenológicas coincidieron con las más altas poblaciones. El estudio reporta que los nematodos más frecuentes en el tomate en el occidente de Nicaragua son Meloidogyne, Pratylenchus, Tylenchorhynchus sp. y Helicotylenchus. Igualmente, confirma que suelos franco-arenosos y ausencia de rotación de cultivos promueven su incremento poblacional de nematodos. Además, se determinó que el crecimiento poblacional de Meloidogyne y Pratylenchus en el tomate se ajusta a un modelo matemático lineal, con una correlación positiva y lineal entre las etapas fenológicas y el incremento de nematodos.
Literatura citada
Agrios, GN. 1997. Plant pathology. Academic Press. New York, USA. 4 ed. p. 565-598. [ Links ]
Bello, A; Lara, MP. 1986. Nematodos ectoparásitos de la superfamilia Criconematoidea, Taylor, 1936 (Geraert, 1966) encontrados en España continental. Bol. San. Veg. Plagas 12:51-93. [ Links ]
Bitterlin, MW; Gonsalves, D. 1987. Spatial distribution of Xiphinema rivesi and persistence of tomato ringspot virus and its vector in soil. Plant Disease 71:408-411. [ Links ]
Blancard, D. 2011: Enfermedades del tomate. Identificar, conocer, controlar. Ed. Mundi-Prensa. Madrid, España. 212 p. [ Links ]
Bouyoucos, GJ. 1936. Directions for making mechanical analysis of soils by the hydrometer method. Soil Sci. 42(3):225-229. [ Links ]
Buyer, JS; Roberts, DP; Russek-Cohen, E. 2002. Soil and plant effects on microbial community structure. Canadian Journal of Microbiology 48:955-964. [ Links ]
Cadet, P; Masse, D; Thioulouse, J. 2005. Relationships between plant-parasitic nematode community, fallow duration and soil factors in the Sudano-Sahelian area of Senegal. Agriculture, Ecosystems & Environment 108:302-317. [ Links ]
Castrillón, A; Castrillón, M. 2000. Reconocimiento de nematodos parásitos del cultivo del plátano (Musa AAB Simmonds) clon Dominico Hartón, en la granja LUKER (Palestina). Estudio de caso. Universidad de Caldas, Manizales, Colombia. p. 25. [ Links ]
Cavigelli, MA; Lengnick, LL; Buyer, JS; Fravel, D; Handoo, Z; McCarty, G; Millner, P; Sikora, L; Wright, S; Vinyard, B; Rabenhorts, M. 2005. Landscape level variation in soil resources and microbial properties in a no-till corn field. Applied oil Ecology 29:99-123. [ Links ]
Chen, P; Tsay, TT. 2006. Effect of crop rotation on Meloidogyne spp. and Pratylenchus spp. populations in strawberry fields in Taiwan. Journal of Nematology 38(3):339-344. [ Links ]
FAO. 2010. FAOSTAT-Agriculture (en línea). Consultado 14 julio 2011. Disponible en http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor. [ Links ]
Fernández-Solano, OM; Quesada-Solís, AS. 2009. Nemátodos asociados a los cultivos de Costa Rica. MAG. San José, Costa Rica. 49 p. [ Links ]
Gómez, TJ. 1983. Nematodos fitoparásitos y su importancia económica en plantas de plátano en Colombia. Memorias Primer Seminario Internacional sobre Plátano. Manizales. p. 80-96. [ Links ]
González, CM; Aristizábal, J; Aristizábal, J. 2007. Dinámica poblacional de nematodos fitopatógenos en plátano (Musa AAB) Dominico Hartón. Agronomía 15(2):25-31. [ Links ]
Griffiths, BS; Caul, S. 1993. Migration of bacterial-feeding nematodes, but not protozoa to decomposing grass residues. Biology and Fertility of Soils 15:201-207. [ Links ]
Guzmán-Hernández, TJ; Hernández-Villalobos, S; Varela-Benavides, I; Durán-Mora, J; Montero-Carmona, W. 2011. Nematodos fitoparásitos asociados al arroz en las regiones Huetar Norte y Huetar Atlántica de Costa Rica. Agronomía Mesoamericana 22(1):21-28. [ Links ]
Haque, MS; Mukhopadhyaya, MC. 1982. Infectivity of Rotylenchulus reniforrais on castor under moisture stress and salinity conditions of soil. Indian Phytopathology 35:518-520. [ Links ]
Herrera, SIC; Marbán-Mendoza N. 1999. Efecto de coberturas vivas de leguminosas en el control de algunos fitonematodos del café en Nicaragua. Nematrópica 29:223-232. [ Links ]
Herrera, I; Bryngelsson, T; Monzón, A. 2011. Occurrence of Meloidogyne sp. and Pratylenchus sp. in conventional and organic coffee systems in Nicaragua. Nematropica 41:82-90. [ Links ]
Hooper, DJ; Hallmann, J; Subbotin, SA. 2005. Methods for extraction, processing and detection of plant and soil nematodes. In Luc, M; Sikora, RA; Bridge, J. eds. Plant Parasitic Nematodes in Subtropical and Tropical Agriculture. 2 ed. CABI, UK. p. 53-86. [ Links ]
Johnson, AW. 1985. Specific crop rotation effects combined with cultural practices and nematodes. In Sasser, JN; Carter, CC. eds. An advanced treatise on Meloidogyne. Volume I. Raleigh, NC: North Carolina State University Graphics. USA. p. 283-301. [ Links ]
Karssen, G; Moens, M. 2006. Root-knot nematodes. In Perry, RN; Moens, M. eds. Plant nematology. CAB International, Wallingford, UK. p. 59-90. [ Links ]
Khan, RM; Khan, AM; Khan, MW. 1985. Cellular changes in tomato root following infection of reniform nematode. Pakistan Journal of Nematology 3:73-82. [ Links ]
MAGFOR (Ministerio Agropecuario y Forestal). 2000. Regionalización biofísica para el desarrollo agropecuario de los Departamentos de León y Chinandega. Managua. Nicaragua. p. 179. [ Links ]
Mai, WF; Mulling, PG. 1996. Plant-parasitic nematodes: a pictorial key to genera. Cornell University Press. Ithaca, NY, USA. 277 p. [ Links ]
Masse, D; Manlay, RJ; Diatta, M; Pontanier, R; Chotte, JL. 2004. Soil organic matter dynamic and nutrients balance in a short-term fallows with different types of vegetation experiments in Senegal. Soil Use Manage 20:92-95. [ Links ]
Martínez, B; Salazar, W. 2007. Mapeo de nematodos fitopatógenos en cinco comunidades ubicadas en la cuenca baja del Molino Sur, Municipio de Sébaco, durante el ciclo agrícola 2006. Tesis. Ingeniero en Agroecología Tropical. Universidad Nacional Autónoma de Nicaragua-León. León, Nicaragua. 69 p. [ Links ]
Overman, AJ. 1999. Diseases caused by nematodos. In Jones, JB; Jones, JP. Stall, RE; Zitter, TA. eds. Compendium of tomato diseases. 2 ed. The American Phytopathological Society Press. St. Paul, MN. p. 49-52. [ Links ]
Perdomo, JR. 1990. Evaluación de alternativas de control de M. incognita en tomate y distribución y frecuencia de nematodos asociados a los principales cultivos hortícolas en el Valle de Comayagua, Honduras. Tesis de Magister Science. CATIE. Costa Rica. p. 65. [ Links ]
Robinson, AF; Heald, CM; Flanagan, SL; Thames, WH; Amador, J. 1987. Geographical distributions of Rotylenchulus reniformis, Meloidogyne incognita, and Tylenchulus semipenetrans in the Lower Rio Grande Valley as Related to Soil Texture and Land Use. Annals of Applied Nematology 1:20-25. [ Links ]
Salazar, W. 2009. Manejo agroecológico de nematodos fitopatógenos. Cuenta Reto del Milenio. UNAN-León. León, Nicaragua. p. 67. [ Links ]
Salazar, W; Berrios, V; Estrada, D; Caballero, A. 2009. Enfermedades de hortalizas: una guía para su identificación y manejo en campo. Cuenta Reto del Milenio. UNAN-León. León, Nicaragua. 103. P. [ Links ]
Sasser, JN; Freckman, DW. 1987. A world perspective on nematology. In Dickson, W; Veech, J. eds. Vistas on nematology. Hyattsville, MD, USA. p. 7-14. [ Links ]
Sasser, JN. 1989. Plant-parasitic nematodes: the farmer´s hidden enemy. Department of Plant Pathology and Consortium of International Crop Protection, North Carolina State University. Raleigh. USA. p. 115. [ Links ]
Shurtleff, MC; Averre III, CW. 2000. Diagnosing plant diseases caused by nematodes. The American Phytophatological Society. St. Paul, Minnesota, USA. p. 187. [ Links ]
Starr, JL; Robinson, AF; Smith, RG; Krausz, JP. 1993. Meloidogyne incognita and Rotylenchulus reniformis associated with soil textures from some cotton production areas of Texas. Journal of Nematology (Supplement) 25:895-899. [ Links ]
Tzortzakakis, EA. 2000. The effect of Verticillium chlamydosporium and oxamyl on the control of Meloidogyne javanica on tomatoes grown in a plastic house in Crete, Greece. Nematología Mediterranea 28:249-54. [ Links ]
Windham, GL; Barker, KR. 1986. Effects of soil type on the damage potential of Meloidogyne incognita on soybean. Journal of Nematology 18:331-338. [ Links ]
*Correspondencia a:
Wilber Salazar-Antón. Laboratorio de Fitopatología. Departamento de Agroecología. Universidad Nacional Autónoma de Nicaragua-León. Nicaragua. Apartado postal N.° 64. Teléfono:(505) 311-1779 / Fax: (505) 311-1780. wilber.salazar@ct.unanleon.edu.ni
Tomás de Jesús Guzmán-Hernández. Instituto Tecnológico de Costa Rica, Sede San Carlos. Apartado 159-7050, Florencia, Alajuela. Tel: (506) 2475-5310. tjguzman@itcr.ac.cr
1. Este trabajo forma parte de la tesis de doctorado en Ciencias Naturales para el Desarrollo (DOCINADE) del primer autor.
2. Laboratorio de Fitopatología. Departamento de Agroecología. Universidad Nacional Autónoma de Nicaragua-León. Nicaragua. Apartado postal N.° 64. Teléfono:(505) 311-1779 / Fax: (505) 311-1780. wilber.salazar@ct.unanleon.edu.ni
3. Instituto Tecnológico de Costa Rica, Sede San Carlos. Apartado 159-7050, Florencia, Alajuela. Tel: (506) 2475-5310. tjguzman@itcr.ac.cr
Recibido: 7 de junio, 2012. Aceptado: 1 de abril, 2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}