











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
Telipogon Kunth (Oncidiinae) comprises approximately 250 species ranging from Mexico to South America and the Antilles. This genus is notably diverse in the humid, montane forests of southern Central America and the Tropical Andes (Amezcua-Trigos et al. 2018, Pérez-Escobar et al. 2017a, POWO 2024, Pupulin & Bogarín 2023), where it likely originated and rapidly diversified (Pérez-Escobar et al. 2017b). Phylogenetically, Telipogon, Hofmeisterella Rchb.f., and Trichoceros Kunth form the Telipogon Alliance within the Oncidiinae (Williams et al. 2005). These genera are distinguished by their unique morphological features, including actinomorphic or zygomorphic flylike flowers, four pollinia, and lamellate viscidia in Hofmeisterella; uncinate viscidia in Telipogon; and cochleariform viscidia in Trichoceros (Martel et al. 2020b). Additionally, some species of Telipogon and Trichoceros employ mimicry of tachinid flies, using specialized pollination strategies like pseudocopulation, deceiving pollinators into mating with the flowers (Martel et al.2019, 2020b).
Dressler (1993) treated the members of the Telipogon Alliance, including Stellilabium Schltr., within the subtribe Telipogoninae. Later, Dressler (1999) classified Stellilabium into four sections: Stellilabium (including Cordanthera L.O.Williams, Darwiniera Braas & Lückel, and Sodiroella Schltr.), Dipterostele, Rhamphostele, and Taeniorhachis. Therefore, Stellilabium, as circumscribed by Dressler (1999), comprises smaller plants, typically with deciduous leaves during flowering, inflorescences with a flattened peduncle and rachis, and smaller, monosymmetrical flowers (less than 2 cm) with ciliate or denticulate petals. However, molecular studies showed Stellilabium nested within Telipogon in the Oncidiinae (Chase 2009, Neubig et al. 2012, Williams et al. 2005). Despite the ongoing debate regarding the broad circumscription of Telipogon (Kolanowska et al. 2017, Martel et al. 2020a), we follow the classification initially proposed by Williams et al. (2005) until further research involving more comprehensive clade sampling and multiple molecular markers, provides more precise insights into the relationships within the Telipogon alliance.
Telipogon, in a broad sense, is recognized by the epiphytic or terrestrial (in humus-rich soil) growth habit, absent or reduced pseudobulbs, conduplicate, persistent or deciduous, distichous leaves, and inflorescences featuring either a cylindrical, flattened or triquetrous peduncle and rachis and the uncinated viscidia. Telipogon, in the strict sense, is known for its showy, large-flowered plants, which are either monosymmetric or semi-actinomorphic, displaying subequal, inconspicuous triangular sepals and often larger, more visually striking petals and lips adorned with reddish or purple stripes or reticulations (Martel et al. 2019, Pabón-Mora & González 2008) including Hofmeisterella and Stellilabium.
While the taxonomy of the large-flowered Telipogon species in Costa Rica has been revised (Dodson & Escobar 1987a, 1987b, Pupulin & Bogarín 2023), its closely related group (Williams et al. 2005), the Central American clade of small-flowered Telipogon (formerly Stellilabium) lacks a comprehensive taxonomic treatment (Dressler 2001, Pupulin 2003). This group, comprising 16 of the 55 species of Telipogon in the country, is predominantly epiphytic and found at elevations between 500 and 2500 meters (Bogarín 2012, Pupulin & Bogarín 2023, Pupulin et al. 2023).
Given the need for further taxonomic revision, this paper focuses on the small-flowered Telipogon group in Costa Rica. Our study involves a morphological comparison of two newly identified species with the known species from the Neotropics. This comparative study has resulted in the identification and description of two novel small-flowered Telipogon species.
Materials and methods
We collected plants in the wild in September 2002, November 2022, and February 2023 from the Orosi and Pejibaye regions adjacent to Tapantí National Park, Cartago, Costa Rica. The living specimens were photographed using advanced macro photography equipment, including a Nikon D850 camera paired with a Nikon AF-S DX NIKKOR 18-105mm f/3.5-5.6 G ED macro lens. We utilized Nikon PB6 bellows and a 40 mm Zeiss Luminar lens for enhanced detail. Photo stacking, which improves the depth of field in the images, was performed using ZereneStacker® software. Detailed botanical drawings were produced using a range of Leica® stereomicroscopes (MZ7.5, MZ9.5, and M80) equipped with drawing tubes to complement the photographic documentation. These hand-drawn sketches were digitized and compiled into comprehensive composite plates using Adobe Photoshop 2024®.
Further, detailed botanical illustrations in black and white were created using an Apple Pencil® on an iPad Pro® tablet, adhering to specific settings (8.5 × 11 inches, 800 dpi resolution). The digitized sketches were further printed and refined with stippling and shading using a 0.1 mm Rotring rapidograph® to enhance three-dimensional details. Our species descriptions and comparative analysis were based on wild-collected specimens. We referred exclusively to the protologues or taxonomic studies cited in the literature section for species comparisons and plant descriptions. Voucher specimens were deposited in the JBL and USJ herbaria.
Taxonomic treatment
Telipogon lateritius Pupulin, sp. nov.
TYPE: Cartago: Orosi, Purisil, road to Monte Sky, 1550-1600 m, premontane wet forest, epiphytic on trees in thick layers of debris along the roadside, 29 September 2002, F. Pupulin 4186, R.L. Dressler, K. Dressler, H. Léon-Páez, A.C. Rodríguez & E. Salas (holotype, USJ). Fig. 1, 3A.
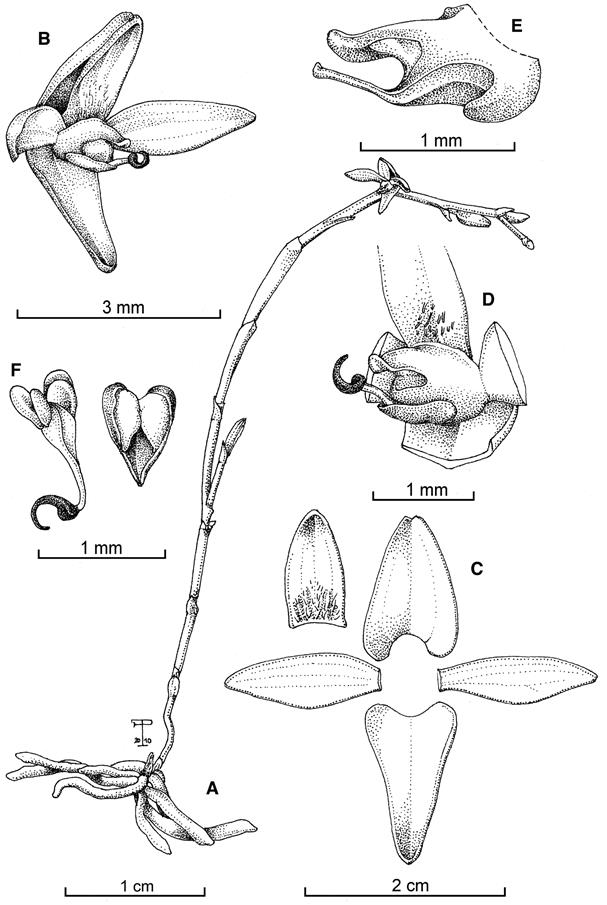
Drawn by F. Pupulin from the holotype.
A. Habit. B. Flower. C. Dissected perianth. D. Column with pollinarium and anther cap, part of the lip and petals, – view. E. Column, emasculate, lateral view. F. Pollinarium and anther cap.
Figure 1 Telipogon lateritium Pupulin.
Diagnosis: Species quae ad Stellilabium globi pertinet, ab omnibus aliis generis sui speciebus floribus glabris, lateritiis brunneo striatis, sepalis lateralibus in triangulari-ovato synsepalo connatis, disco labelli paucis trichomatibus sparsis recedit.
Plant epiphytic, small, monopodial herb to 6 cm long, including the inflorescence. Roots flexuous, numerous, short, to 2 mm in diameter, with green apices. Stem abbreviated, completely concealed by the base of the amplectant leaf-sheaths. Leaves 2, elliptic, acute, gently ribbed on the underside, 16-22 × 4-6 mm, often deciduous at flowering. Inflorescence lateral, from the base of the stem, a successively few-flowered (to 4-5) raceme to 6 cm long, sometimes with a short branch about 2 cm long in the lower half; the peduncle born terete and becoming dorsiventrally flattened-ancipitous toward the apex, the flattened internodes with a faint longitudinal keel, provided with a triangular, obtuse, weakly keeled, pale green, glumaceous bract at the apex, decurrent on the peduncle, becoming dry-papyraceous with age; the rachis subtrigonous, straight to lightly sigmoid. Floral bract triangular, acute, strongly conduplicate-keeled, 1.5 × 1.0 mm. Pedicellate ovary terete-subclavate, rounded in section, ca. 2 mm long. Flowers non-resupinate, the sepals not completely spreading, brick-colored, the sepals, petals, and lip longitudinally striped with chestnut brown, the anther cap brownish red. Dorsal sepal triangular, acute, the margins shortly inflexed, 1.6 × 0.9 mm, 3-veined. Lateral sepals connate into a triangular-ovate synsepal, acute, minutely emarginate, 1.5 × 1.0 mm, 5-veined, the margins lightly involute to become adpressed to the apex of the lip. Petals asymmetrically lanceolate, acute, 1.7 × 0.5 mm, 3-veined. Lip narrowly ovate from a broad base, acute, conduplicate-involute at apex, 1.2 × 0.6 mm, the basal third provided with short, stiff, scattered hairs. Column semiterete, 1 mm long, the stigma recessed under a long stigmatic lobe, the anther dorsal (facing down in natural position), the rostellum long-attenuated, reaching the apex of the stigmatic lobe. Anther cap cucullate, cordate, acute, with two large flaps protecting the pollinia. Pollinia 4, discoid-complanate, in two pairs of different size, on a lanceolate, conduplicate, hyaline stipe and a hook-shaped, brown viscidium.
Distribution: endemic to Costa Rica.
Etymology: from the Latin lateritius, “made of brick”, in reference to the brick-colored flowers.
Habitat and Ecology: The species has only been observed in the type collection from the premontane wet forest of the Caribbean watershed of the Cordillera de Talamanca, near the Monte Sky area, close to Tapantí National Park in Costa Rica. This area is known as one of the country’s rainiest regions (Boza 1986).
Phenology: Flowering of T. lateritius was documented in September.
Notes: Telipogon lateritius is part of a group of species characterized by their small size, often deciduous at flowering, with dorsiventrally complanate and ancipitous inflorescences and small, glabrous flowers. Dressler (1999) previously assigned these species to the Stellilabium Section Rhamphostele. This group predominantly inhabits the southern region of the Mesoamerican isthmus, including species such as T. barbozae (J.T.Atwood & Dressler) N.H.Williams & Dressler, T. campbelliorum (J.T.Atwood) N.H.Williams & Dressler, T. fortunae (Dressler) N.H.Williams & Dressler, along with other yet-to-be-described species from Costa Rica and Panama. Telipogon lateritius is distinguished within this group by its brick-colored flowers, sepals, petals, and lip striped with chestnut brown; the lateral sepals connate into a triangular-ovate synsepal and the sparse, stiff pubescence at the base of the lip.
Telipogon muntzii Bogarín, O.Pérez & Pupulin, sp. nov.
TYPE: Costa Rica. Cartago: Jiménez, Pejibaye, Reserva Biológica Guaitil, ca. 6 km al norteste de El Humo, entre Ríos Humo y Río Taus, 9°47’44.48”N 83°45’07.82”W, 1152 m, bosque pluvial premontano, epífita en bosque secundario a orillas del sendero, 25 noviembre 2022, D. Bogarín 14185, R. Gómez, G. Villalobos & R. Müntz (holotype, JBL; isotype, JBL). Fig. 2, 3B, 4.
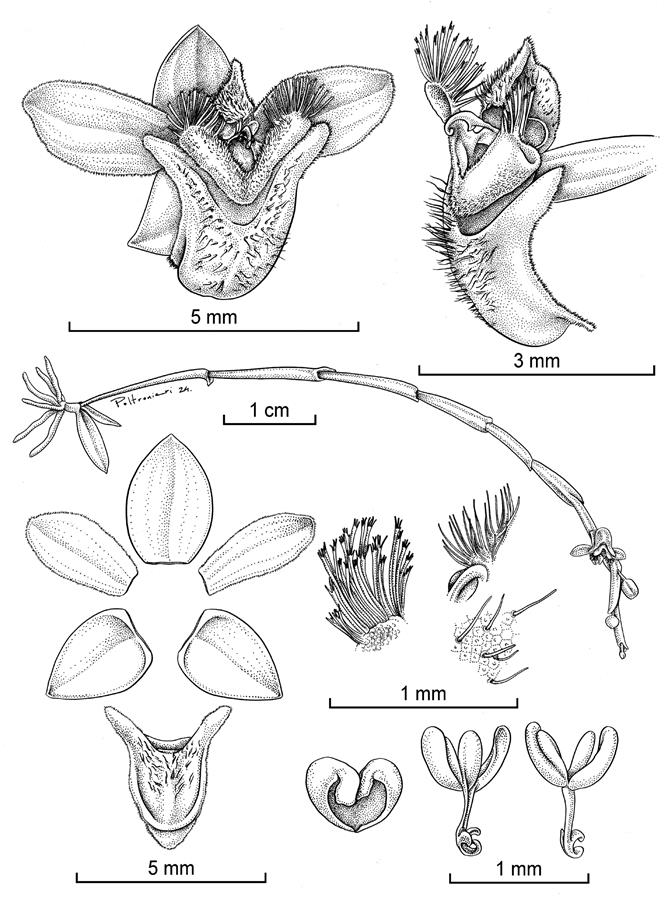
Drawn by D. Bogarín and S. Poltronieri from the holotype.
A. Habit. B. Flower. C. Ovary, column, and lip, lateral view. D. Dissected perianth. E. Setae of the lateral, median lobe of the column and the lip, from left to right. F. Pollinarium and anther cap.
Figure 2 Telipogon muntzii Bogarín, O.Pérez & Pupulin.

Photos by F. Pupulin (A, C) and D. Bogarín (B).
B.Telipogon muntzii (D. Bogarín 14185, JBL). C.Telipogon anacristinae (D. Bogarín 14185, JBL).
Figure 3 Flower morphology of A . Telipogon lateritium (F. Pupulin 4186, USJ).

Photo by D. Bogarín from the paratype.
Figure 4 Macrophotography of Telipogon muntzii (D. Bogarín 14494, JBL).
Diagnosis: Species praecipue Telipogoni anacristinae (Pupulin) Dressler & N.H.Williams similis, sed floribus viridi-olivaceis, labello sparsim trichomatoso, lobulis lateralis columnae anguste oblongis trichomatibus dendromorphibus solum apice munitis, galea lobuli medii apicaliter decrescenti solum trichomatibus simplici fornita recedit.
Plant epiphytic, small, nearly acaulescent. Roots thick, flexuous, grayish with green tips, rounded in section, 2 mm in diameter. Stem subterete, abbreviated about 3 mm long, completely concealed by the base of the amplectant leaf sheaths. Leaves 2 (-3), green, distichous, conduplicate, narrowly elliptic to lanceolate, acute, 1.0 × 0.3 cm, the base enclosing the stem, frequently leafless at flowering. Inflorescence a simple, successively flowered (to 6) raceme to 8 cm long; peduncle terete, to 5 cm long, with 2-6 infundibuliform, triangular, acute bracts about 1.5 mm long; rachis flattened, 2 mm wide; floral bracts triangular, obtuse, slightly carinate along the middle, the margins decurrent along the edges of the rhachis, 1 × 2 mm. Ovary linear-subclavate, ca. 3 mm long, including the pedicel. Flowers non-resupinate, small, with greenish-yellowish sepals, olive green petals, and lip, the lip with purple bases of the bristles, the column greenish-purplish with purple dendroid bristles, and purple anther cap, to 0.7 mm in diameter. Dorsal sepal elliptic, acute, 3.4-3.5 × 2.2-2.3 mm. Lateral sepals ovate to lanceolate-elliptic, acute, concave, strongly deflexed in natural position, 2.9-3.0 × 2.0-2.2 mm wide. Petals elliptic to obovate, subobtuse to acute, ciliate-denticulate, 3.2-3.3 × 1.5-1.7 mm, 3-veined. Lip 3-lobed, hastate, 3.7-3.9 × 3.7-3.9 mm wide across the lateral lobes, the basal lobules narrowly linear, acute, the margins finely ciliate, the median lobe ovate-elliptic, subacute, the lamina convex, rounded up to near the acute apex, sparsely covered with stiff hairs, the margins minutely ciliate-denticulate. Column 3-lobed, the lateral lobes narrowly oblong, flattened, V-shaped, pubescent, 2.4-2.5 × 0.9-1.0 mm, the apices ca. 2.0-2.2 × 2.3-2.4 mm, covered with short dendroid-stellate bristles, the median lobe cucullate, narrowing towards the apex, 1.3 × 0.9 mm, covered with short, simple hairs; stigma rounded, shiny, surrounded by the lobes of the column. Anther cap cucullate, cordate, 1-celled, 8 × 5 mm. Pollinia 4 in two pairs of different sizes, oblong, laterally complanate-concave, on an oblong, basally attenuate stipe; viscidium uncinate.
Additional material examined: Cartago: Jiménez, Pejibaye, Reserva Biológica Guaitil, ca. 6 km al norteste de El Humo, entre Ríos Humo y Río Taus, 9°47’35.615”N 83°45’14.05444”W, 1186 m, bosque pluvial premontano, epífita en bosque secundario a orillas del sendero, 3 febrero 2023, D. Bogarín 14494, J. Fernández, R. Müntz & F. Pupulin (JBL) (Fig. 4).
Distribution: endemic to Costa Rica.
Eponymy: Dedicated to Dr. Robert Müntz, founder of Remedia Homeopathy GmbH and a visionary in the field of homeopathic pharmacy. Beyond his professional interests, Robert is an avid jungle explorer and nature conservationist. His active involvement in preserving Costa Rica’s tropical forests through the Guaitil Biological Reserve in Pejibaye de Jiménez, Cartago, has been inspiring. We are pleased to dedicate this species to Robert for his unwavering support for orchidological research and his passionate commitment to conserving their fragile ecosystems.
Habitat and ecology: The species has only been observed in the type collection from the premontane wet forest of the Caribbean watershed of the Cordillera de Talamanca, near El Humo de Pejibaye and Taus area, close to the northern boundaries of Tapantí National Park in Costa Rica. Plants grow on branches of young trees in secondary forest.
Phenology: Plants flower from November to February.
Notes: Telipogon muntzii exhibits morphological similarities to a group of species characterized by their distinctly 3-lobed lips and columns, with dendroid bristles on the lateral lobes and either simple or dendroid bristles on the median lobe. Dressler (1999) previously assigned these species to what was then known as the Stellilabium Section Taeniorhachis. Among these species, T. muntzii is morphologically similar to T. anacristinae (Pupulin) Dressler & N.H. Williams (Fig. 3C). However, it can be distinguished by several key features: T. muntzii has olive green flowers, compared to the pale rose to greenish-rose of T. anacristinae. Additionally, the lip of T. muntzii is sparsely covered with trichomes that develop from black glands, unlike the densely stiff-trichomed lip of T. anacristinae. The lateral lobes of the column in T. muntzii are narrowly oblong, bearing dendroid trichomes solely at the apex, a contrast to the elliptic, rounded lobes with dendroid trichomes along the lobes seen in T. anacristinae. Lastly, the helmet-shaped median lobe of the column in T. muntzii, which narrows towards the apex and is covered exclusively by simple bristles, differs from the rounded median lobe of T. anacristinae, which is covered by both simple and dendroid trichomes.
Other morphologically similar species to T. muntzii are T. distantiflorus (Ames & C.Schweinf.) N.H.Williams & Dressler and T. smaragdinus (Pupulin & M.A.Blanco) N.H.Williams & Dressler, especially in the green emerald color of flowers; however, these species have longer inflorescences, more than 10 cm in length, and an abruptly acute median lobe of the lip with an acute apicule (as opposed to less than 5 cm in T. muntzii and with an acute median lobe of the lip). Additionally, both species occur at higher elevations in oak montane forests (between 1500-2400 m) along the Cordillera de Talamanca, whereas T. muntzii occurs in the premontane wet forest of the Caribbean watershed of Cordillera de Talamanca.
Also, T. erratus (Dressler) N.H.Williams & Dressler, a common species around the Tapantí area, differs from T. muntzii in the inflorescences, longer than 16 cm in length, and the dark wine-purple flowers (as opposed to the greenish-yellowish sepals and olive-green petals of T. muntzii).

