











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Las enfermedades cardiovasculares continúan siendo la causa principal de morbilidad y mortalidad en los países desarrollados y su incidencia es aún mayor en aquellos en vías de desarrollo (Nichols, 2012). El evento común que desencadena las dos manifestaciones clínicas más frecuentes y dañinas de las dolencias del sistema circulatorio -la isquemia cardíaca (o infarto) y la enfermedad cerebrovascular (o ictus)- es la formación de coágulos sanguíneos (trombos) que interrumpen el suministro de oxígeno y nutrientes al corazón y al cerebro, respectivamente. A pesar de la importancia médica de estas patologías, las bases estructurales y funcionales de estos procesos no se conocen por completo, por lo que una comprensión profunda de los mecanismos que conducen a la formación del coágulo de fibrina es fundamental para obtener una imagen detallada de los eventos patofisiológicos asociados con la generación de los trombos. Esta información es también fundamental para interpretar las consecuencias de las mutaciones identificadas en pacientes con hemofilia o con trombosis, y para avanzar hacia el tratamiento personalizado de las enfermedades cardiovasculares, redundando así, a mediano o largo plazo, en importantes beneficios sanitarios y económicos.
El proceso de coagulación de la sangre ocurre como resultado de una compleja red de proteínas activadas de forma concertada, progresiva y estrictamente regulada, que incluyen varios lazos de retroalimentación positiva y negativa. Estos lazos son esenciales para una rápida -pero controlada- formación del coágulo, desencadenada por estímulos fisiológicos en los vasos sanguíneos dañados (por ejemplo, al sufrir una herida en la piel). Entre las proteínas más importantes que participan en este proceso se encuentra la trombina, que pertenece a la familia de las proteasas serínicas similares a la tripsina. Su precursor, la protrombina, se sintetiza en el hígado y se secreta al torrente sanguíneo. Esta proteasa se considera uno de los factores de la coagulación más importantes porque juega un papel clave en la activación de varias moléculas esenciales para la formación del coágulo, principalmente el fibrinógeno, el factor XIII y los receptores activados por proteasas (PARs) en la superficie de las plaquetas. Asimismo, la trombina inicia uno de los más importantes lazos de retroalimentación positiva al modular su propia generación activando los cofactores procoagulantes V (FV) y VIII (FVIII) (Davie et al., 1991).
Los cofactores procoagulantes FV y FVIII son grandes glicoproteínas multidominio que comparten una organización de dominios A1-A2-B-A3-C1-C2. Polipéptidos relativamente largos y ricos en residuos ácidos llamados a1, a2 y a3 conectan los dominios del FVIII A1-A2, A2-B y B-A3, respectivamente, dando una organización de dominios que se puede representar como A1-(a1)-A2-(a2)-B-(a3)-A3-C1-C2. De forma similar, la estructura de dominios del FV se puede describir como A1-A2-(a2)-B-(a3)-A3-C1-C2. Ambos cofactores son activados por la trombina mediante proteólisis limitada de enlaces peptídicos específicos. En el caso del FVIII humano, la trombina hidroliza los enlaces Arg372-Ser373 entre las regiones a1 y A2, Arg740-Ser741 entre el conector a2 y el dominio B, y Arg1689-Ser1690 en la frontera a3-A3, generándose el heterotrímero A1/A2/A3-C1-C2 y liberando el altamente glicosilado dominio B (ver figura 1).
El reconocimiento y procesamiento de muchos sustratos fisiológicamente relevantes de la trombina requiere interacciones iniciales con regiones positivamente cargadas presentes en la superficie de esta proteasa, y distantes del sitio activo, conocidas como exositios I y II. El exositio I es esencial para el reconocimiento de sustratos (fibrinógeno, PARs, FV, FVIII), el receptor endotelial trombomodulina y diversos inhibidores como la hirudina (Corral-Rodríguez et al., 2011; Gandhi et al., 2010; Fuentes-Prior et al., 2000). Por otra parte, el exositio II es el sitio de unión del sulfato de heparán y la heparina, así como nuevamente de los cofactores V y VIII (Corral-Rodríguez et al., 2011, Segers et al., 2007; Myles et al., 2002).
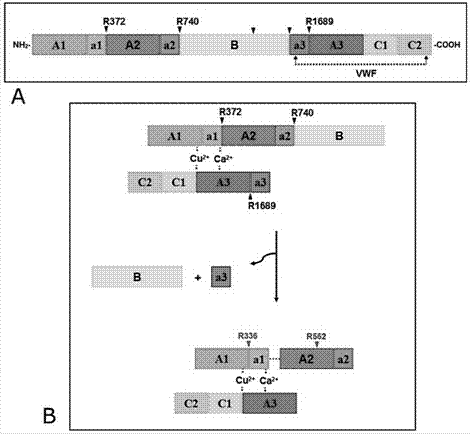
Figura 1: Representación esquemática del mecanismo de activación del factor VIII. Los factores V y VIII son activados por proteólisis limitada efectuada principalmente por la trombina, en presencia de fosfolípidos. A. Estructura del FVIII inactivo con los dominios globulares A1, A2, A3, B, C1 y C2, así como los conectores interdominios a1, a2 y a3. Estos conectores son regiones ricas en residuos ácidos que preceden a los sitios de corte y activación de la trombina. Dos flechas en el dominio B indican los sitios de corte reconocidos por las endoproteasas intracelulares que dan origen al heterodímero del FVIII. Se muestra también la zona de interacción con su proteína transportadora, el factor de von Willebrand (vWF). B. La trombina hidroliza los enlaces Arg372-Ser373, Arg740-Ser741 y Arg1689-Ser1690 en el heterodímero del FVIII (R372, R740 y R1689, respectivamente, indicados con flechas). Como consecuencia de estas reacciones se libera el dominio B y el conector a3 de la molécula precursora, dando origen al heterotrímero activo, el FVIIIa, compuesto ahora por los dominios A1, A2, y una cadena formada por los módulos A3, C1 y C2. Este heterotrímero se asocia por interacciones electroestáticas (interlineado) y es estabilizado por iones Cu+/2+ y Ca2+. Las flechas rojas indican los sitios de corte de la proteína C activada (APC), que conducen a la disociación del dominio A2 y con ello a la pérdida de la función activadora del cofactor.
Estudios bioquímicos previos sugieren que la activación de los cofactores V y VIII por la trombina requiere de interacciones entre los exositios I y II y los conectores ácidos (a1, a2, a3) que preceden a los residuos de corte (figura 1A), ya que estos contactos permiten orientar los enlaces peptídicos a hidrolizar dentro del sitio activo de la trombina, en una conformación óptima para la catálisis (Newell & Fay 2008; Segers et al., 2007; Myles et al., 2002). Sin embargo, el conocimiento acerca de las bases atómico-moleculares de estas interacciones es todavía muy limitado y permanece casi sin explorar. Esta información estructural y funcional es esencial para obtener una comprensión profunda de los procesos que conducen a la formación de trombos.
Por esta razón, para estudiar las interacciones moleculares entre el FVIII humano y la trombina, se sobreexpresaron de forma heteróloga en Escherichia coli los conectores interdominios ácidos del factor VIII (FVIIIa1, FVIIIa2 y FVIIIa3). Esto permitió obtener los fragmentos proteicos recombinantes suficientes para evaluar su calidad y pureza, y realizar ensayos preliminares para la búsqueda de las condiciones de cristalización de complejos binarios específicos de FVIII con trombina.
Materiales y Métodos
Clonaje de los conectores interdominios del FVIII humano
Las secuencias de los conectores del FVIII, junto a algunas de sus características fisicoquímicas relevantes, se muestran en el cuadro 1. Los fragmentos FVIII (330-372), FVIII (712-740) y FVIII (1649-1689), en adelante llamados FVIIIa1, FVIIIa2 y FVIIIa3, incluyen una cola de histidinas en el extremo N-terminal que facilita su purificación por cromatografía de afinidad. Se trabajó también con variantes del FVIII con mutaciones puntuales del conector, FVIIIa1 (R372Q, G373P) y FVIIIa3 (R1689Q).
Cuadro 1 Secuencias y características fisicoquímicas básicas de los fragmentos derivados del factor VIII humano. Se incluye el peso molecular (Pm) expresado en daltons (Da), el punto isoeléctrico teórico (pI) y la absorbancia teórica para una solución de 1 g/L de la proteína pura a 280 nm (Abs 0,1%). Los parámetros se calcularon utilizando las herramientas ProtParam del servidor de ExPASy (www.expasy.org/tools) (Wilkins et al., 1999).
| Fragmento | Secuencia de aminoácidos | Pm (Da) | pI | Abs280 0,1% |
|---|---|---|---|---|
| FVIIIa1 (330-372) | ASHHHHHHPEEPQLRMKNNEEAEDYDDDLTDSEMDVVRFDDDNSPSFIQIRGSGC | 6404.7 | 4.51 | 0.233 |
| FVIIIa2 (712-740) | ASHHHHHHGDKNTGDYYEDSYEDISAYLLSKNNAIEPRGSGC | 4726.9 | 5.75 | 1.261 |
| FVIIIa3 (1649-1689) | ASHHHHHHGEITRTTLQSDQEEIDYDDTISVEMKKEDFDIYDEDENQSPRGSGC | 6284.5 | 4.54 | 0.474 |
Los conectores FVIIIa1, FVIIIa2 y FVIIIa3 fueron clonados en el vector pET-3a, respectivamente. Con cada plásmido se transformaron, mediante choque térmico, diferentes cepas competentes de E. coli [BL21(DE3), BL21(DE3) pLysS y BL21(DE3) pLysE] por triplicado. Las cepas transformadas se incubaron en placas Petri con agar LB, ampicilina (100 ug/mL) y/o cloranfenicol (34 ug/mL) y se incubaron durante toda la noche a 37 °C.
Para verificar la transformación se realizaron extracciones de ADN de plásmido a partir de precultivos de al menos tres colonias transformadas, empleando el kit comercial GeneJet Plasmid Miniprep Kit (Fermentas #K0503). Las muestras se cuantificaron en un espectrofotómetro (NanoDropTM Lite) y se visualizaron en un gel de agarosa al 1%. Las muestras por triplicado se enviaron a secuenciar a Macrogen (Maryland, Estados Unidos).
Sobreexpresión y purificación de los fragmentos recombinantes
A partir de las muestras confirmadas por secuenciación de ADN, se seleccionó una única colonia y se inoculó en 50 mL de medio LB con los antibióticos correspondientes. El cultivo se incubó durante la noche a 37 °C con agitación constante a 225 rpm. Al día siguiente, los 50 mL del precultivo se inocularon en 0.5 litros de medio LB con antibióticos, en un matraz de 2 litros, a 37 °C con agitación constante a 225 rpm. Una vez alcanzado el valor OD entre 0.4-0.6 para la absorbancia a 600 nm, se realizó la inducción adicionando IPTG a una concentración final de 0.55 mM. Se incubó a 37 °C durante seis horas con agitación constante a 225 rpm, se centrifugaron las células cultivadas a 4500 g durante 20 minutos y el sedimento obtenido se congeló a -20 °C.
La extracción de las proteínas sobreexpresadas en E. coli se realizó mediante lisis celular por congelación/descongelación, agregando tampón de lisis (40 mM Tris pH 8.0, 100 mM NaCl, 20 mM MgCl2, 1 mM PMSF, 1mM beta-mercaptoetanol, 0.5 ug/mL ADNasa). Se incubó durante 30 minutos en rotor y luego se realizaron dos ciclos de congelación/descongelación. Seguidamente, se centrifugó a 12000 g durante 30 minutos a 4 °C para separar la fracción soluble de la insoluble. A partir de la fracción soluble se efectuó la purificación de la proteína de interés mediante incubación con la resina de afinidad Ni-NTA acoplada a agarosa (Qiagen). La suspensión con resina se trasvasó a una columna, se recuperó la fracción no unida a la resina, se realizaron dos lavados con 30 mM imidazol, 50 mM NaH2PO4, 300 mM NaCl, pH 8.0. Finalmente, se realizó la elución del material unido de forma específica a la resina empleando 500 mM imidazol, 50 mM NaH2PO4, 300 mM NaCl, pH 8.0. Las muestras purificadas se evaluaron mediante SDS-PAGE al 12%.
La autenticidad y pureza de los fragmentos recombinantes purificados se comprobó mediante espectrometría de masas (MALDI TOF/TOF). Se evaluó la masa de cada uno de los fragmentos recombinantes sin digerir y digeridos con las endoproteinasas Asp-N o tripsina (RocheDiagnostics), según correspondía. Los resultados de espectrometría de masas se analizaron con el programa ProteinPilot.
Cristalización de los fragmentos recombinantes purificados con trombina humana
Para la búsqueda de las condiciones de cristalización, los fragmentos del FVIII purificados se incubaron durante 30 minutos a temperatura ambiente con trombina-α humana (Haematologic Technologies, Inc.), previamente dializada contra el tampón HEPES/NaCl pH 6.0. Los complejos respectivos se prepararon con un exceso molar del fragmento recombinante del sustrato (2:1) en el mismo tampón y se concentraron a 2.5 mg/mL utilizando dispositivos Amicon Ultra-15 con poro nominal de 3kDa (Millipore). Se empleó la técnica de difusión de vapor utilizando la modalidad de gota colgante (hanging drop) en placas para cristalización (ComboPlates™, Greiner). Se evaluaron los kits de cristalización: Structure Screen, Morpheus y ProPlex (Molecular Dimensions), para un total de 292 condiciones de cristalización evaluadas. Se mezcló 1 µL de disolución concentrada de proteína con 1 µL de solución de cristalización y se incubó a 20 °C. Las gotas fueron periódicamente inspeccionadas utilizando un estereoscopio MEIJI EMZ-TR (Meiji Techno Corporation).
Resultados
Clonaje y sobreexpresión de los fragmentos recombinantes del FVIII
El análisis del clonaje y de la secuencia de los plásmidos de expresión conteniendo los conectores FVIIIa1, FVIIIa2 y FVIIIa3, y sus variantes, se confirmó mediante secuenciación de ADN luego de comparar la secuencia teórica de cada uno de los plásmidos codificantes in silico con el electroferograma de secuenciación respectivo. Como ejemplo, en las figuras 2 y 3 se muestra la secuenciación de ADN y la traducción in silico del FVIIIa3.
Posteriormente, los primeros experimentos de sobreexpresión de los fragmentos se llevaron a cabo en medio de cultivo LB con la cepa BL21 (DE3) de E. coli a 37 °C, y se evaluaron los resultados después de la inducción a las 2, 4, 6, 8 y 12 h. Se comprobó que a tiempos superiores a las seis horas de inducción no había un incremento proporcional en los niveles de sobreexpresión, y además se comenzaban a detectar productos de degradación del conector. También se hicieron experimentos de expresión en las cepas de E. coli BL21, pLysS y pLysE, para determinar si con ellas aumentaba la cantidad del fragmento recombinante obtenido. Sin embargo, la sobreexpresión de los conectores a1, a3 y sus respectivas variantes tuvieron rendimientos similares en las tres cepas, y en todas ellas el fragmento se expresaba soluble (datos no mostrados). Consecuentemente, en los experimentos de sobreexpresión que se realizaron de forma rutinaria para estos conectores se trabajó con la cepa E. coli BL21(DE3) pLysS.
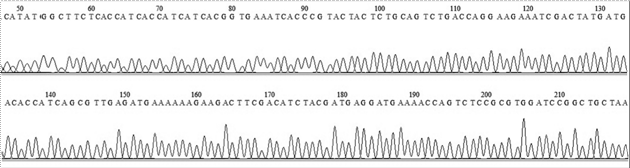
Figura 2: Electroferograma de la secuenciación del FVIIIa3. Se muestra únicamente el rango de nucleótidos donde se encuentra la secuencia del gen que codifica para el conector del factor FVIIIa3. La secuenciación fue realizada por la empresa Macrogen y se visualizó utilizando el software BioEdit (versión 7.1.9).

Figura 3: Traducción in silico de la secuencia nucleotídica a los residuos aminoacídicos correspondientes del FVIIIa3. Se presenta en letra minúscula la secuencia de nucleótidos (tripletas) que codifican para el conector interdominio FVIIIa3 y en letra mayúscula los aminoácidos respectivos para cada codón. La traducción se realizó utilizando la herramienta ExPASy (http://web.expasy.org/translate/).
Extracción, purificación e identidad de los fragmentos del FVIII
Una vez extraídas las proteínas por medio de la técnica de congelación/descongelación, se procedió a trabajar con la fracción soluble obtenida. Los fragmentos se purificaron por cromatografía de afinidad por metal con Ni-NTA, eluyendo de la resina de afinidad con 500 mM de imidazol, obteniéndose un considerable grado de pureza (figura 4).
El rendimiento promedio de recuperación del fragmento recombinante FVIIIa1 fue de 0,5 mg de proteína a partir de 1 litro de cultivo bacteriano, mientras el del FVIIIa3 fue de 0,4 mg. Los productos obtenidos fueron analizados por espectrometría de masas en modo negativo, identificando la presencia del fragmento FVIIIa1 y el fragmento FVIIIa3 según correspondía y confirmando su pureza, al observar un pico de masa de 6404.55 Da y de 6284.44 Da, respectivamente (datos no mostrados).
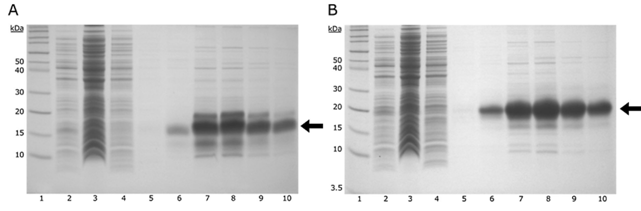
Figura 4: Análisis por SDS-PAGE de la expresión heteróloga y la purificación del conector FVIIIa1 (A) y FVIIIa3 (B). Se evaluaron los niveles de proteína expresados durante las seis horas de inducción tras añadir 0.55 mM IPTG al cultivo bacteriano. El carril 1 corresponde al marcador de peso molecular (Novex Sharp). En el carril 2 se muestra una alícuota de las células totales; en el carril 3 se visualiza la fracción de proteínas no unidas a la resina de afinidad Ni-NTA; en los carriles 4 y 5 se presentan alícuotas de lavados sucesivos con 30 mM de imidazol; mientras que en los carriles 6 a 10 se muestran las fracciones de eluciones sucesivas de la resina con 500 mM de imidazol, donde se evidencia el fragmento de interés purificado (indicado con una flecha).
Comprobación de la secuencia de los conectores FVIIIa1 y FVIIIa3 mediante digestión enzimática
Además, la identidad de los fragmentos FVIIIa1 y FVIIIa3 purificados se confirmó por espectrometría de masas a partir de fragmentos generados mediante digestión con la endoproteinasa Asp-N o tripsina, respectivamente. Las masas obtenidas se compararon con las masas teóricas según PeptideMass de ExPASy (Wilkins et al., 1999) y con mMass 3.9.0 (Strohalm et al., 2008). Se comprobó la presencia de picos con relaciones m/z correspondientes a las masas teóricas de los fragmentos monocargados generados (figuras 5 y 6).
Expresión y purificación del fragmento FVIIIa2
En ensayos preliminares se expresó el fragmento FVIIIa2 en las diferentes cepas de E. coli indicadas anteriormente y a temperaturas de 28, 30, 34 y 37 °C durante seis horas de inducción. Los mejores resultados se obtuvieron con la cepa BL21(DE3) al inducir a 30 °C durante seis horas. Sin embargo, la mayor parte del fragmento se encontró en la fracción insoluble, por ello, el sedimento obtenido tras la centrifugación se incubó en tampón Tris/NaCl que contenía concentraciones crecientes de úrea (2, 4, 6 y 8 M). Al analizar las fracciones correspondientes por SDS-PAGE, se observó que el fragmento de interés se extraía mayoritariamente en 4 y 6 M de úrea. La purificación del fragmento FVIIIa2 se realizó unificando los extractos obtenidos de las disoluciones de 4 y 6 M de úrea y empleando la resina Ni-NTA agarosa. Las fracciones correspondientes se evaluaron mediante SDS-PAGE al 12% (datos no mostrados). El rendimiento promedio de recuperación del conector FVIIIa2 fue de ~0.2 mg de proteína a partir de 1 litro de cultivo bacteriano, y las muestras se debieron mantener en tampón Tris/NaCl con 2 M de úrea. No se continuó trabajando con este fragmento debido a que las altas concentraciones de úrea interfieren en los ensayos de cristalización.
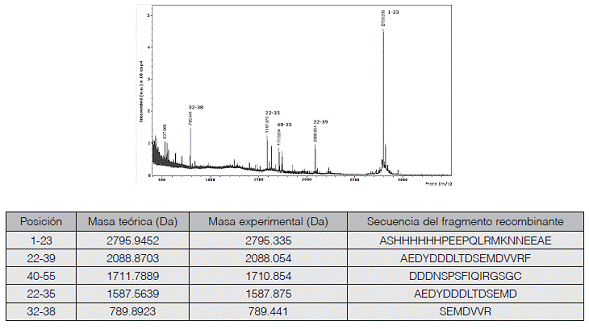
Figura 5 Verificación de la identidad del fragmento recombinante FVIIIa1. El conector FVIIIa1 purificado fue digerido con la endoproteinasa Asp-N y los péptidos resultantes se analizaron por espectrometría de masas en modo reflector con polaridad negativa. En el espectro mostrado se identifican los picos correspondientes a las masas que se indican en la tabla correspondiente, donde aparecen las masas teóricas calculadas con PeptideMass (www.expasy.org/tools/ peptide-mass.html) (Wilkins et al., 1999) y con mMass 3.9.0 (Strohalm et al., 2008). Se verificó la secuencia completa del fragmento FVIIIa1 a partir de la posición 1 (Ala) hasta la 55 (Cys) del fragmento recombinante.
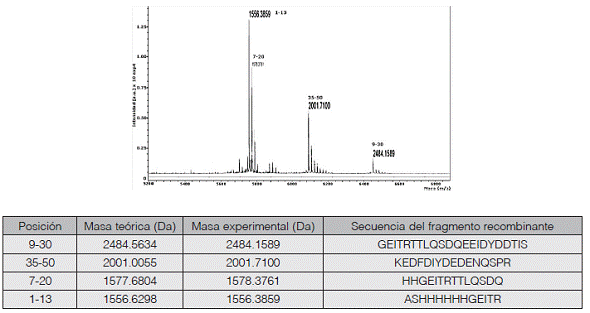
Figura 6 Verificación de la identidad del fragmento FVIIIa3. La proteína recombinante purificada fue digerida con tripsina, y los péptidos resultantes se analizaron por espectrometría de masas en modo reflector con polaridad positiva. En el espectro mostrado se identifican los picos correspondientes a las masas que se indican en la tabla correspondiente, donde aparecen las masas teóricas calculadas con PeptideMass y con mMass.
Cristalización de los complejos proteicos
En la búsqueda de las condiciones de cristalización de los fragmentos purificados FVIIIa1, FVIIIa3 y sus variantes -acomplejados con trombina humana-, se evaluaron 296 tampones de cristalización distintos con el objetivo de encontrar condiciones que favorezcan la formación de cristales proteicos de calidad. A partir del complejo FVIIIa1 (R372Q, G373P)·Trombina se observó crecimiento de cristales en la condición 1-33 de Structure Screen a los ocho días (figura 7). Uno de los cristales obtenidos mide aproximadamente 44 µm de largo por 6 µm de ancho (figura 7A), y al repetir la condición para aumentar el número de cristales se obtuvieron varios con dimensiones de 20 µm x 14 µm y con un mayor grosor (figura 7B), lo que podría permitir recolectar un mejor conjunto de datos de difracción.

Figura 7: Cristales obtenidos de la variante del FVIIIa1 (R372Q, G373P) en complejo con trombina. A. Cristal de proteína obtenido en la condición 1-33 de Structure Screen (Molecular Dimensions) conteniendo 0.2 M de MgCl2·6 H2O, 0.1 M de Tris pH 8.5 y 30% p/v de PEG 4000. B. La repetición de la misma condición permitió obtener cristales más pequeños, pero más gruesos. Imágenes capturadas con la cámara HDCE-II, empleando el programa ScopeImage 9.0, en un estereoscopio MEIJI EMZ-TR (Meiji Techno Corporation).
Discusión
Sobreexpresión heteróloga de los conectores ácidos FVIIIa1, FVIIIa2 y FVIIIa3
Al iniciar la presente investigación se partió de la hipótesis de que los interconectores de los dominios A1-A2, A2-B y B-A3 del factor VIII de la coagulación son importantes tanto para la activación del cofactor como para la actividad del FVIII dentro del complejo X-asa (CorralRodríguez et al., 2011, Newell & Fay 2007; Yuan et al., 2005), lo que se evidencia por el alto grado de conservación entre diferentes especies de mamíferos (figura 8) tanto para el conector FVIIIa1 que incluye los residuos Pro330-Arg372 (figura 8A), como el FVIIIa2 con los residuos Asp712Arg740 (figura 8B) y el FVIIIa3 que contiene la región Glu1649-Arg1689 (figura 8C). Estos conectores preceden a los sitios de corte reconocidos por la trombina que son relevantes fisiológicamente, Arg372-Ser373, Arg740-Ser741 y Arg1689-Ser1690, respectivamente.

Figura 8 Alineamientos múltiples de los conectores del FVIII de diferentes especies de mamíferos. Se presentan las secuencias de las regiones FVIIIa1 (A), FVIIIa2 (B) y FVIIIa3 (C). La numeración corresponde al FVIII humano maduro. Los residuos estrictamente conservados en cada uno de los conectores se muestran con fondo gris oscuro; otros residuos bien conservados o reemplazados conservativamente están sombreados en gris claro. Las regiones ricas en aminoácidos de carácter ácido o aromático se muestran dentro de un recuadro y el sitio de corte por la trombina se indica con un triángulo invertido. En el panel B se presenta la secuencia del extremo C-terminal de la hirudina (Hir-C) debajo de las secuencias del conector FVIIIa2 topológicamente equivalentes. En C, nótese que el conector FVIIIa3 presenta dos regiones diferenciadas ricas en aminoácidos de carácter ácido. Los alineamientos se generaron con T-Coffee (Notredame et al., 2000; Poirot et al., 2003).
La activación del FVIII mediada por la trombina es fundamental para los procesos procoagulantes, de ahí la importancia de estudiar las interacciones trombina-FVIII. Resultados publicados sobre la interacción de los residuos Glu666-Glu672 del conector a2 del FV con el exositio I de la trombina (Corral- Rodriguez et al., 2011) evidencian la importancia de regiones ricas en residuos ácidos y aromáticos en estas interacciones. Por otra parte, otros investigadores han reportado que los residuos ácidos Glu720-Asp725 del conector FVIIIa2 (Newell & Fay, 2008) son importantes para la activación del FVIII mediada por la trombina. Esta información previa, junto con la característica de que cada uno de los conectores del FVIII contiene regiones ricas en residuos ácidos que les confieren cargas netas negativas, sugiere que estos conectores interaccionan con los exositios básicos I y II de la trombina para presentar los sitios de corte y activación del cofactor en el centro activo de la proteasa.
Estas observaciones nos llevaron a estudiar la formación de complejos de los conectores ácidos del FVIII con la trombina. Para cumplir este objetivo, se clonaron, sobreexpresaron y purificaron los tres conectores del FVIII, así como sus variantes puntuales. Aunque los constructos iniciales utilizados en estos estudios se optimizaron de forma similar, se observaron diferencias significativas en la solubilidad de los fragmentos recombinantes. Por ejemplo, los conectores a1 y a3 y sus respectivas variantes se lograron extraer con buen rendimiento en la fracción soluble a partir de la sobreexpresión en células de E. coli, probablemente debido a que son cadenas polipeptídicas relativamente cortas, carentes de elementos de estructura secundaria y con un notable carácter polar (45-46% de los residuos). Mientras que el FVIIIa2 se extrajo en tampones conteniendo 4-6 M de úrea, probablemente este fragmento recombinante tiende a agregar debido a su menor porcentaje de residuos polares (38% vs 45-46%).
Al evaluar las proteínas recombinantes mediante SDS-PAGE, se observó que para todos los casos tanto del FVIIIa1 como del FVIIIa3 y sus respectivas variantes, cuyo peso molecular teórico está entre aproximadamente 6.4 y 6.3 kDa, migraron a la misma distancia aproximada que la banda de 20 kDa del marcador de peso molecular utilizado. Estudios han demostrado que los múltiples residuos ácidos presentes en una proteína son capaces de neutralizar parte de las cargas negativas (aún en geles desnaturalizantes), disminuyendo de manera considerable la migración en el gel de poliacrilamida (Shi et al., 2012, Rath et al., 2009). Debido a lo anterior, la identidad de estas proteínas se confirmó mediante espectrometría de masas analizando los fragmentos completos, así como cada uno de los picos generados por la digestión limitada. A partir de estos últimos se logró obtener la secuencia experimental de cada fragmento y se identificó una alta homología con el Factor VIII de la coagulación de Homo sapiens (Uniprot: P00451) con un porcentaje de confianza del 99%. Estos resultados de expresión, purificación e identificación confirman que es posible expresar de forma heteróloga estos fragmentos del FVIII, en cantidades y con un nivel de pureza adecuados para estudios bioquímicos y de cristalización.
Búsqueda de condiciones de cristalización de los conectores del FVIIIa1, FVIIIa3 y sus variantes respectivas, unidos a la trombina
La sobreexpresión en E. coli de los fragmentos FVIIIa1 (337-372), FVIIIa3 (1649-1689), así como de los mutantes FVIIIa1 (R372Q, G373P) y FVIIIa3 (R1689Q), permitió obtener suficiente cantidad de proteínas con una alta calidad y grado de pureza, lo cual es casi siempre el cuello de botella para poder realizar estudios de cristalización y cristalografía debido a la gran cantidad de proteínas que se requieren (Rupp, 2009).
Nuestros resultados previos con los experimentos de band shift de los conectores FVIIIa1 y FVIIIa3 que evidencian la interacción con la trombina (datos no mostrados), nos motivaron a buscar condiciones de cristalización de complejos de estos fragmentos del FVIII con la proteinasa, con el objetivo de obtener cristales de calidad, para posteriormente resolver las estructuras 3D de dichos complejos. Sin embargo, según nuestra experiencia con trabajos previos en la resolución de estructuras 3D (Hernández-Carvajal, 2013), se decidió modular el sitio de corte y activación de los conectores para favorecer la forma acomplejada con la trombina. Para ello se realizaron mutaciones dirigidas en las posiciones P1 y P1’ sobre los residuos de arginina y serina, los cuales fueron reemplazados por glutamina y prolina, respectivamente. Dado que la glutamina tiene una cadena lateral no cargada y la prolina dificulta a las proteasas serínicas procesar los enlaces Xxx-Pro, se espera que estos cambios favorezcan la formación de complejos más estables entre la trombina y sus sustratos.
Los cristales conseguidos están siendo almacenados para para realizar experimentos de difracción de rayos X en los sincrotrones ALBA (Barcelona, España) o ESRF (Grenoble, Francia), para evaluar el resultado de esta estrategia y tratar de resolver la estructura tridimensional del complejo.
Conclusiones
Se sobreexpresaron heterólogamente en E. coli y se purificaron los fragmentos del FVIII humano correspondientes a los conectores FVIIIa1, FVIIIa2 y FVIIIa3, así como las variantes FVIIIa1 (R372Q, G373P) y FVIIIa3 (R1689Q), evidenciando la capacidad de producir suficiente proteína para ensayos bioquímicos y de cristalización.
Se realizó la búsqueda de condiciones de cristalización, obteniéndose cristales del complejo Trombina·FVIIIa1 (R372Q, G373P) en la condición Structure Screen 1-33 que contiene 0.2 M de MgCl2·6 H2O, 0.1 M de Tris pH 8.5 y 30% p/v de PEG 4000.

