











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Diferentes factores ecológicos, biológicos, químicos y físicos influyen en el éxito de eclosión de los nidos, por lo que algunos animales han desarrollado diferentes estrategias para aumentar la sobrevivencia de las crías. Por ejemplo, las tortugas marinas seleccionan determinados lugares para anidar, mostrando preferencias sobre las diferentes zonas y pendientes en la playa utilizadas para la construcción de los nidos.
Dicha selección depende, probablemente, de un conjunto de características como el oxígeno que fluye entre el sustrato, la salinidad que alcanza el suelo, la humedad relativa, la temperatura, el tipo y densidad de la arena, las luces artificiales que se encuentran en la localidad, la pendiente y la extensión de la playa. Estos factores son considerados determinantes para que el proceso reproductivo ocurra exitosamente, ya que son fundamentales en la preservación del nido y la eclosión de las crías (Zurita et al., 1993; Bolongaro et al., 2010).
El Caribe de Costa Rica representa uno de los sitios de mayor importancia para la anidación, alimentación, copulación, y desove de algunas especies de tortugas marinas (Chacón, 1999). Entre las que destaca la tortuga baula (Dermochelys coriacea) cuya anidación ha sido reportada en tres áreas protegidas, a saber, el Refugio Nacional de Vida Silvestre Gandoca-Manzanillo (REGAMA), la Reserva Forestal Pacuare-Matina (RFPM) y en el Parque Nacional Tortuguero (PNT) (Eckert, 1999). Se considera que esta colonia de baulas es la tercera en magnitud para la región del gran Caribe, y la cuarta en importancia a nivel mundial (Chacón-Chaverri, 2004a).
Por su parte, las tortugas carey anidan en las playas del PNT, el REGAMA y el Parque Nacional Cahuita (PNC). La población más representativa de esta especie en el Gran Caribe anida en la costa Mexicana, en la Península de Yucatán con más de 1000 hembras anidadoras/año (Meylan, 1999; Chacón-Chaverri, 2004b).
Actualmente, las zonas de anidación de tortugas marinas son vulnerables como consecuencia de las presiones naturales y antrópicos a los que están expuestos los ambientes costeros y que actúan sobre la morfología de la costa. Esto afecta los procesos de ovoposición por la disminución del espacio disponible y las constantes modificaciones de las playas, generando impactos negativos sobre el estado de estas poblaciones (Bolongaro et al., 2010).
Estas condiciones así como la cacería y la extracción de huevos, han provocado que las tortugas baula y carey se encuentren incluidas dentro de la categoría de peligro crítico, según la UICN (2013). En tanto, que en el Reglamento a la Ley de Conservación de Vida Silvestre, Decreto N° 32633-MINAE, son consideradas en peligro de extinción así como en el apéndice I de la Convención sobre el Comercio Internacional de Especies Silvestres de Fauna y Flora (CITES,2013).
Estas condiciones han sido otorgadas como consecuencia de la mortalidad de adultos y huevos por distintas causas como lo son el saqueo de nidos, la pérdida de nidadas por inundación o la erosión de las playas, la depredación natural e infecciones por microorganismos, la sobrepesca comercial, la captura accidental, la destrucción de los hábitats de alimentación y la contaminación de los mares (Bell et al., 2003).
En Gandoca, Chacón (1999) estudió los patrones de anidación de la tortuga baula. Sin embargo, la estructura de la playa ha cambiado en relación con la pendiente, el ancho y la erosión de playa. Por lo que estas situaciones pueden haber cambiado. Por lo tanto, el presente trabajo pretendió actualizar el patrón de anidación en las tortugas marinas Dermochelys coriacea (tortuga baula) y Eretmochelys imbricata (tortuga carey) en el sector Gandoca, del Refugio Nacional de Vida Silvestre Gandoca Manzanillo (REGAMA), Limón, Costa Rica con el fin de proponer medidas de manejo.
Área de estudio
El sector Gandoca del Refugio Nacional de Vida Silvestre Gandoca Manzanillo (REGAMA), Limón, Costa Rica que fue creado en Julio de 1985 mediante el Decreto Ejecutivo 16614MAF01 como área silvestre protegida y declarada como sitio RAMSAR, el 11 de diciembre de 1995; formando parte del Área de Conservación La Amistad Caribe (ACLA-C) (Weitzner y Fonseca, 2000) (Figura 1). El REGAMA comprende un área de 5013 hectáreas continentales y 4.436 hectáreas marinas (SINAC 2013). Se ubica en los 9°59.972' latitud norte y 82°60.530' longitud oeste con una extensión de 8,85 km desde Punta Mona hasta la desembocadura del río Sixaola (Matthew 2010), de los cuales, 7,4 km son áreas disponibles para la anidación y 1.1 km corresponden a zonas alrededor de las desembocaduras de ríos y quebradas (Chacón,1999).
El régimen de precipitaciones oscila entre los 2500 y 3100 mm anuales, y las temperaturas promedio varían entre 25°C y 27°C. El clima presenta dos estaciones lluviosas definidas, una entre noviembre y marzo y la segunda entre junio y agosto. La tasa de precipitación anual en este sector sur del REGAMA es menor, alrededor de 2500 mm en tanto que en la sección norte alcanza hasta 3100 mm. El microclima se caracteriza por lluvias nocturnas y períodos soleados durante el día (Herrera, 1984). Las mareas son mixtas, principalmente diurnas, con una amplitud de 30 hasta 50 cm. Las corrientes fluyen de noroeste a sureste, con pequeños remolinos en dirección opuesta (Cortés, 1994).
Dentro de las asociaciones vegetales que prevalecen se encuentra la transición entre palmares pantanosos y bosque húmedo tropical o palmar mixto (Coll et al., 2001). La playa se caracteriza por su alta energía y pendientes, con corrientes marinas fuertes, aguas profundas muy cerca de la costa y las formaciones coralinas presentes en el sector de Punta Mona (Chacón, 1999). Sin embargo, esta playa se ha modificado drásticamente en los últimos años, aumentó la pendiente de la misma y los signos de erosión son cada vez más marcados que podrían ser en respuesta al aumento del nivel del mar en el Mar Caribe (Figura 2). Además, debido al aprovechamiento forestal en la cuenca del río Sixaola y las quebradas de la zona se da el arrastre y acumulación de madera y desechos orgánicos a lo largo de la playa (Programa de Conservación de Tortugas Marinas, Caribe Sur, Costa Rica, 2006).

Materiales y métodos
La playa se dividió en cinco sectores denominados A, B, C, D y E, el primer sector inicia en Punta Mona y finaliza en la desembocadura del río Sixaola. Cada sector fue marcado con mojones separados a una distancia de 50 m. Se realizaron dos recorridos nocturnos, cado uno con una duración de cuatro horas, el primero se desarrolló entre las 20:00 y las 24:00 horas; en tanto que el segundo ocurrió entre las 24:01y las 4:00 a. m. Se registraron las observaciones directas y los rastros de las tortugas marinas que anidaron entre febrero y agosto del 2011 y 2012.
El registro consistió en anotar el sector, mojón y posición de la playa donde anido la tortuga, siendo alta, media y baja; la fase lunar así como presencia o ausencia de lluvia durante el proceso de anidación.
El análisis de los datos consistió en una prueba t de comparación de medias para una p < 0.05, con la previa comprobación de la normalidad y homogeneidad de varianza de los datos. Para los análisis estadísticos de los datos de anidación durante los dos años se utilizó el programa Statgraphics Centurion XV, Info Stat (versión 2013I) y Excel Microsoft office Excel 2007 para la elaboración de las figuras.
Resultados
Durante el 2011 y 2012, en la playa del sector Gandoca del REGAMA se observó la presencia de tres especies de tortugas marinas anidando, a saber: Dermochelys coriácea o tortuga baula que presentó 1005 registros; Eretmochelys imbricata o tortuga Carey con 47 registros; y Chelonia mydas o tortuga verde con solamente 6 registros.
El periodo de anidación de la tortuga baula ocurre, principalmente, durante los meses de marzo hasta finales de julio. Este patrón fue coincidente para ambos años analizados. Los períodos donde la anidación presentó el menor número de eventos se presentó entre enero, febrero y agosto, con un importante aumento durante el mes de marzo, con un punto máximo durante el mes de mayo del 2011 en el que se registraron 209 individuos; y en abril del 2012 con el registro de 229 individuos (Figura 3).
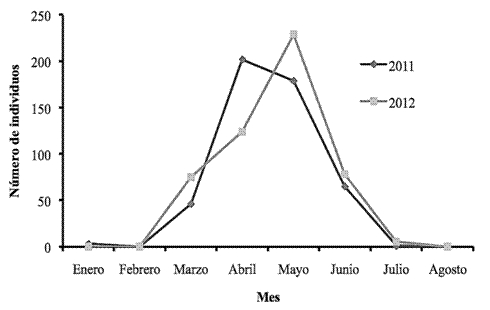
Figura 3: Tortugas baulas observadas mensualmente durante el período 2011-2012 en el sector Gandoca, REGAMA, Limón, Costa Rica.
Las tortugas carey fueron registradas anidando entre los meses de abril y julio en el 2011, con un punto máximo en mayo y junio. Mientras que, para el año 2012, se tuvieron registros desde mayo hasta agosto, en este último se obtuvo el mayor número de individuos registrados con un total de 8, en tanto las tortugas verdes se avistaron principalmente durante el mes de marzo (5 individuos) y en agosto (1 individuo) de este mismo año.
Durante el periodo de estudio, se registraron un total de 226 nidos, de los cuales 197 (87, 17%) corresponden a nidadas de la tortuga baula y los 29 restantes a la tortuga carey, mientras que la tortuga verde no se tuvo registros de que elaborara algún nido. Debe mencionarse que 152 (67,25 %) nidos fueron registrados en el 2011 mientras que, durante el 2012, solo se registraron 74; mostrándose una disminución considerable en la cantidad de nidos durante el segundo año.
Para ambas especies estudiadas, se observó preferencia en la posición de la playa que utilizan para construir y ovopositar los nidos. En el caso particular de la tortuga baula, muestra preferencia para emplear con mayor frecuencia la parte media, mientras que la tortuga carey utilizó, principalmente, la parte baja o sea, más cercana a la línea de pleamar para anidar (t (223)= 17.26, p ≤ 0.05) (Figura 4).
El sector de la playa en la zona de estudio que la tortuga baula utilizó con mayor frecuencia fue el sector C. Éste fue donde recurrió para la elaboración de los nidos, para un total de registros de 130. Seguidamente empleó los sectores B y D.
En el caso de las tortugas carey, la mayor cantidad de nidos fueron construidos en el sector E, el cual fue usado exclusivamente por esta especie. Mientras que el sector A fue empleado por las tortugas baula y carey en proporciones similares (Figura 5). A pesar de lo anteriormente mencionado, no se encontró diferencias significativas en el uso de los sectores para ninguna de las especies (t (6)=2.21, p ≤ 0.05).
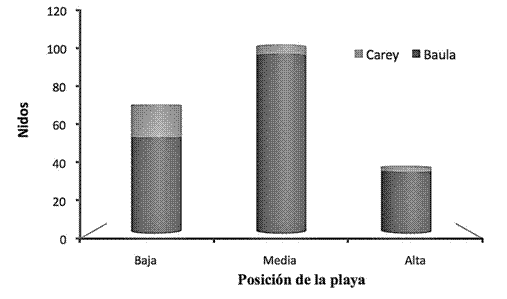
Figura 4: Nidos construidos según la posición de la playa por las tortugas carey y baula durante el periodo 20112012 en el sector Gandoca, REGAMA, Limón, Costa Rica.
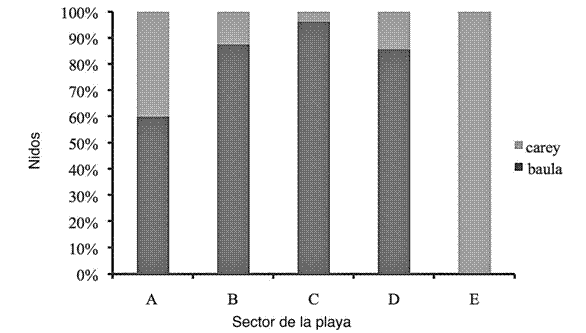
Figura 5: Nidos construidos según los sectores de la playa por las tortugas carey y baula durante el periodo 20112012 en el sector Gandoca, REGAMA, Limón, Costa Rica.
La cantidad de nidos varió según la fase lunar, tanto para las tortugas baula como para la carey. Presentaron el mismo comportamiento para el cuarto menguante, que fue el período donde se registró la máxima cantidad de nidos ovopositados. Mientras que durante la luna llena y nueva, se registraron el número más bajo de nidadas observadas (Figura 6). No obstante, no se determinó diferencias significativas entre el número de nidos construidos según la fase lunar (t(8)= 2.64, p ≤ 0.05).
Discusión
Los factores que influyen en la selección de los sitios de anidación de las tortugas marinas parecen ser múltiples, y su importancia radica en el hecho de que determinan el éxito de la nidada, el adecuado desarrollo de los embriones, la emergencia y supervivencia de los neonatos.
Dentro de las especies de tortugas marinas que visitan el Caribe de Costa Rica, la baula es la que alcanza las mayores dimensiones y el mayor peso; por ende, el proceso de desplazarse sobre la playa representa un mayor esfuerzo energético para ella. Dada esta condición es de esperar, que ésta no se adentre mucho en la playa para anidar. En Gandoca, Costa Rica; las baulas ovopositaron mayormente en la zona media, de igual forma que lo observado por Quiñones et al., (2007) en Chocó, Colombia. Según Chacón (1999), el 32% de las nidadas fueron puestas en la berma, 37% en la zona de marea alta y 31% en la zona de marea baja y, a su vez, indica, para 1991, el 64% de los nidos fueron localizados entre las líneas de marea, con el subsecuente riesgo de pérdida.
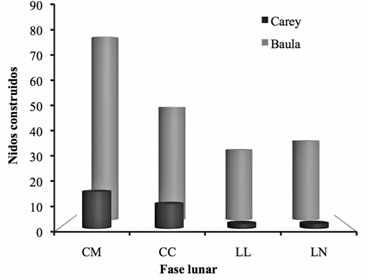
Figura 6: Nidos construidos según la fase lunar durante la construcción del nido por las tortugas carey y baula durante el periodo 2011-2012 en el sector Gandoca, REGAMA, Limón, Costa Rica. CM: cuarto menguante, CC: cuarto creciente, LL: luna llena y LN: luna nueva.
Con los cambios observados en la morfología de la playa, el anidamiento en la zona media de la playa contribuye con la conservación de la especie en el corto plazo, debido a que el aumento en el nivel del mar en el Caribe podría reducir las áreas potenciales de anidación de las tortugas marinas. Preocupa el caso de la tortuga carey, que anida principalmente en la zona baja de la playa aumentando el riesgo de pérdida de nidos, el cual aumenta cada año dado los pronósticos del IPCC (2003). Es urgente atender esta última especie mediante planes de manejo que aseguren la protección de los nidos y para las dos, se debe discutir la posibilidad de manejar viveros fuera de la playa por la alta energía de estas que las hacen inestables.
La anidación en esta zona de la playa supone un menor esfuerzo para la hembra y un menor riesgo de quedar varada o bloqueada por los obstáculos presentes en la playa tales como los troncos de árboles caídos o arrastrados por las mareas, así como la vegetación. Sin embargo, los signos evidentes de erosión en la playa dificultan el ingreso a la zona media, obligando a la hembra a ovopositar en la zona baja o buscar otras playas.
Además, a esto se le suma la presencia de pendientes que le impiden avanzar hacia las áreas más elevadas y las zonas erosionadas que se encuentran a lo largo de la playa. Estas condiciones podrían afectar las nidadas, modificando la humedad relativa en el nido y causando un incremento en la mortalidad de las crías (Bolongaro et al., 2010). De igual manera, si los nidos se colocan cerca de la línea de la pleamar hay un riesgo inminente de ser inundados o arrastrados por las olas (Eckert, 1987).
Según Eckert (1999), las hembras de baula prefieren playas donde la plataforma continental sea corta, que tengan accesos abiertos, con costas de alta energía, fuerte corriente, oleaje alto y sin contacto con rocas o corales que puedan serle abrasivos durante el proceso de alcanzar la playa para anidar. Estas condiciones explican por qué la menor presencia de nidos en el sector A, el cual corresponde a la zona de Punta Mona donde se presentan formaciones coralinas así como la isla Pájaros que dificultan el ingreso de esta especie. Por el contrario, los sectores B, C y D corresponden a zonas que presentan algunas de las condiciones mencionadas por dicho autor siendo por tanto preferidaspara la anidación.
La zona de preferencia para la anidación de la tortuga carey se ubicó en la parte baja de la playa, al igual que lo reportado por Bolongaro et al., (2010) en la playa Chenkan en el Golfo de México. Sin embargo, autores como Medina-Cruz et al.,(2010) y Chacón-Chaverri (2004 a) señalan que las tortugas carey seleccionan zonas cercanas a la vegetación terrestre o bajo ella para anidar, explicando dicho comportamiento con la presencia de raíces que contribuye a que la arena esté menos compacta, propiciando un sustrato adecuado para la construcción de los nidos, en comparación, con las zonas desprovistas de vegetación en donde el grado de compactación es mayor (Horrocks y Scott, 1991; Chen y Cheng, 1995).
A su vez, algunos autores sugieren que debe considerar que cuando el nido se coloca en la vegetación, el riesgo asociado está representado por la penetración de raíces a la cámara del mismo y la destrucción de los huevos. También puede influir sobre el éxito de emergencia de las crías, ya que algunas podrían quedar atrapadas en las raíces o la vegetación (Wood y Bjorndal, 2000). Otro aspecto es la posibilidad de desorientación de las crías en su camino hacia el mar (Kamel y Mrosovsky, 2004) debido a que deben recorrer una distancia mayor aumentando la posibilidad de no alcanzar las olas o bien, sean susceptibles a la depredación (Mrosovsky, 1983; Horrocks y Scott,1991). Estos factores podrían contrarrestar el efecto de la compactación del terreno en áreas sin vegetación y propiciar que las zonas bajas sean beneficiosas para los neonatos durante la emergencia y su recorrido hacia el mar.
Por otra parte, el sector utilizado en mayor proporción por las tortugas carey fue el sector E, que se encuentra en las cercanías de la desembocadura del río Sixaola. Este presenta escombros, troncos y árboles que son arrastrados por las corrientes del río hasta la playa; y propicia cambios en el panorama de la playa, dificultando el acceso de las tortugas hacia el interior (Chacón, 1999). Por tanto, la parte baja es la que está accesible para que esta especie construya sus nidos.
Sin embargo, se debe mencionar que Bolongaro et al. (2010) discute sobre los riesgos de la elaboración de los nidos en la parte baja de la playa, indicando que tienen una menor probabilidad de sobrevivencia por el exceso de humedad, lo cual aumenta la mortalidad de huevos por asfixia (Whitmore y Dutton, 1985). Sin embargo, en el presente trabajo no se comprobó el éxito de eclosión de estos nidos.
Por su parte, la luna es considerada como un generador de energía indirecta que ejerce atracción sobre los cuerpos sólidos y líquidos, determinando las mareas oceánicas e influyendo en las actividades fisiológicas de las plantas y animales (Alonso et al., 2002). Se ha reportado, que, en muchas especies animales, la luna llena y nueva incrementa perceptiblemente la tasa de actividad física, el metabolismo, la agresividad y la conducta sexual (Lieber, 1984). Sin embargo, en este estudio se observó que en las fases de luna llena y luna nueva se registró el menor número de nidos. Esto, probablemente se explicada por la cantidad de luz reflejada en la playa, durante la luna llena, que produce un incremento de la cantidad de horas luz durante la noche y la intensidad lumínica mayor, aumentando el grado de exposición de las tortugas y, por ende, el aumento de las probabilidades de que los huevos o la tortuga sea depredada (Higuera-Moros et al., 2002).
Por el contrario, durante luna nueva, la cantidad de luz es escasa, lo que podría dificultar la capacidad de las tortugas para orientarse en la playa. O bien, podrían estar ocurriendo procesos de copula en el mar (Lieber, 1984), ya que esta suele ocurrir en la superficie de aguas someras cercanas a las playas de anidación (Carr et al., 1978). Mientras que en cuarto creciente y cuarto menguante la cantidad de luz no es ni excesiva ni escasa, brindando a estas especies un escenario ideal para la anidación. Se considera que, el ciclo reproductivo es sincrónico con las fases lunares (Márquez et al., 1976; Casas, 1978); para el caso de especies que presentan arribadas, estás ocurren durante el cuarto menguante (Briceño, 1980). Lo cual coincide, parcialmente, con lo reportado para la baula y la carey en el sector Gandoca del REGAMA y que, aunque no mantienen comportamiento de arribadas, el mayor número de nidos se elaboró durante esta fase lunar.
Sin embargo, en el Pacífico Mexicano, el número de hembras baula que ingresan para anidar aumenta considerablemente durante las noches más oscuras; es decir, durante luna nueva (Márquez et al., 1981). En el Pacífico Sur de Costa Rica durante la temporada del 2011 los eventos de anidación de varias especies, incluidas la baula y la carey fueron menores durante la luna llena, en tanto, las otras fases lunares no mostraron ningunas tendencia durante los meses de estudio (Viejobueno et al., 2012).
Debe considerarse lo indicado por otro autores como Ackerman (1997), quien cuestiona que exista una relación entre el ciclo lunar y la actividad de anidación de las tortugas marinas. Mientras que Lohmannet al., (1997) y Witherington y Martin, (2003) sugieren que las tortugas anidan en un determinado período lunar, previendo que la eclosión de las crías ocurre en la fase de cuarto creciente o luna llena, para facilitar la ubicación de los neonatos respecto al océano durante la noche, siendo guiados por el resplandor de la luz de la luna en la espuma de las olas y la claridad misma de estas (Sánchez, 2007).
Además, se debe considerar el grado de influencia de luz artificial, el tipo e intensidad de esta en la iluminación de las playas. Dicho factor ha sido reportado como una elemento que influye en el número de hembras anidando o el número de anidaciones exitosas. Debido a que causa cambios en el comportamiento de anidación principalmente porque las luces artificiales son interpretadas por las tortugas como luz diurna de modo que las tortugas no salen a anidar, por ejemplo Médicci et al., 2009, en un estudio en Cipara, Venezuela encontró una diferencia en la proporción de salidas de tortugas baula y la construcción de nidos entre los años en los que la playa carecía y en los que presentaba electricidad, y presentó un disminución en el periodo en que la playa estaba iluminada. En el caso del sector de Gandoca del REGAMA, este factor es irrelevante para explicar la llegada del número de tortugas anidantes.
Durante el año 2012 en el REGAMA se registró una disminución en número de anidaciones en comparación con el año 2011. Chacón (1999), reportó una variación en el número de nidadas entre los años 1990 y 1997, en donde durante el año 1996 se da un aumento en el número de nidos construidos, pero en 1997 se da un notable descenso. La disminución en la cantidad de hembras anidantes o bien en el número de nidos elaborados en el REGAMA podría estar relacionada con diferentes factores ambientales como las precipitaciones, cantidad de escombros en la playa, o bien por características intrínsecas de las poblaciones como tasa de mortalidad o de reclutamiento de hembras. Después de su esfuerzo reproductivo, la hembra vuelve a la zona de alimentación, donde se mantiene reproductivamente inactiva durante un periodo variable de años, para acumular energía para la nueva época reproductiva, lo cual implica que no todos los años la cantidad de hembras anidantes sea alta, o bien se mantenga una proporción constante.
Por otra parte Según Pacheco, 2008, las tortugas baula tienen la capacidad de distinguir sitios con características topográficas inadecuadas, ejemplo por efecto de las lluvias, que las inducen a no desovar en esas partes de la playa, en el caso del REGAMA la playa está expuesta a fuertes oleajes y se dan procesos erosivos evidentes que hacen que muchos sectores de la playa no estén disponibles para anidar, esto podría contribuir en la disminución de nidadas que se presentó durante la temporada 2012, ya que en algunas playas sujetas a erosión se forman rompeolas y terrazas verticales que constituyen serias barreras físicas para las tortugas, de modo que se pueden ver obligadas a recorrer grandes trechos paralelos a la costa o bien regresan al mar para anidar en otros tramos más alejados (Bolongaro et al., 2010).
En conclusión, el pico de anidación para la tortuga baula fue entre marzo y finales julio, y para la carey entre abril y julio en el 2011 con un desplazamiento en el 2012 para mayo hasta agosto, además, se dan preferencias en la selección de la zona de playa, la carey anida en la parte baja cerca de la línea de costa y la baula en la parte media. Asimismo, ambas especies utilizan indistintamente los diferentes sectores de la playa para anidar, el mayor número de nidos se ubican en el sector C para la baula y el sector E para la carey. Por último, durante el cuarto creciente y menguante se registran el mayor número de anidaciones; sin embargo, las fases lunares no determinan la cantidad de anidamientos.
Por lo tanto, se recomienda remover los escombros que representen un obstáculo para el desplazamiento de las tortugas en la playa, así como desarrollar un plan de aprovechamiento de escombros de madera por vecinos de zona, para reducir los costos de remoción y aumentar la efectividad, también evaluar las condiciones de playa respecto a los procesos erosivos que podrían dificultar la anidación de las tortugas.

