










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgronomía Costarricense
versão impressa ISSN 0377-9424
Agron. Costarricense vol.37 no.1 San Pedro de Montes de Oca Jan./Jun. 2013
Efecto de la aplicación de los fungicidas propineb y mancozeb sobre el estado nutricional de plántulas de banano en medio hidropónico
Effect of fungicides propineb an mancozeb on the nutritional status of banana plants growing under hydroponic culture
Effect of fungicides propineb an mancozeb on the nutritional status of banana plants growing under hydroponic culture
*Dirección para correspondencia:
Resumen
Se evaluó el efecto de la aplicación reitera-da y/o alterna de los fungicidas propineb y man-cozeb sobre el estado nutricional y el crecimiento de plántulas de banano procedentes de cultivo in vitro. Para ello, se realizó un experimento en cultivo hidropónico con 2 diferentes soluciones nutritivas, con altos y bajos contenidos de Mg y Zn, alto contenido de Mn y niveles óptimos de los demás nutrimentos en ambas soluciones. Los tratamientos, aplicados foliarmente, fueron: testigo, propineb en agua, propineb en emulsión agua:aceite, propineb alternado con mancozeb, mancozeb, y mancozeb + Zn (todos en emulsión). Las variables evaluadas fueron: peso de materia seca (g) y la concentración y absorción de Mg, Mn y Zn en hojas, pseudotallo y raíz. La mayor concentración de Zn en hojas y pseudotallo se obtuvo con los tratamientos que incluyeron pro-pineb, independientemente del nivel de Zn en la solución, que duplica los valores encontrados en el testigo. La concentración de Zn en la raíz fue afectada únicamente por la cantidad de este ele-mento presente en la solución nutritiva. La con-centración y absorción total de Mg se incrementó de manera significativa en todas las partes de la planta al aumentar la cantidad de este elemen-to en la solución nutritiva, mientras que el Mn mostró un comportamiento inverso; por lo que la relación Mg:Mn en los tejidos (especialmente en la raíz) se vio afectada. Ninguno de estos 2 elementos se vio influenciado por la aplicación de los productos fungicidas. El mayor peso de biomasa aérea y radical se obtuvo a partir de la solución nutritiva con bajos niveles de Mg y Zn. La concentración de Ca en los todos tejidos dis-minuyó en un 50% cuando la concentración de Mg se incrementó 8 veces en la solución.
Palabras clave: Nutrición en banano, propineb, mancozeb, micronutrientes, magnesio.
Abstract
The effect of reiterative and/or alternate application of propineb and mancozeb fungicides on the nutritional status and growth of banana seedlings produced in vitro culture was evaluated. The greenhouse experiment was conducted under hydroponic culture with 2 different nutrient solutions, with high and low contents of Mg and Zn, high content of Mn and optimal amounts in the other mineral nutrients in both solutions. The foliar treatments were: a control, propineb in water, propineb in water:oil-emulsion, propineb alternated with mancozeb, mancozeb, and mancozeb + Zn (all of them applied as emulsion). Variables evaluated were: dry matter (g) and concentration and absorption of Mg, Mn and Zn in leaves, pseudostem and roots. Highest Zn concentration in leaves and pseudostem was obtained with treatments including propineb (twice that found in the control treatment), regardless of Zn amounts in the nutrient solution. The concentration of Zn in the root was affected only by the level of this element in the nutrient solution. The concentration and total absorption of Mg in all plant tissues was significantly increased when the amount of this element increased in the nutrient solution, while Mn showed the opposite behavior, the reason why the Mg:Mn ratio in the tissues (specially in the root) was affected. Neither of these 2 elements was influenced by the foliar application of fungicides. The highest dry weight of the aerial and root biomass was obtained with the nutrient solution with low levels of Mg and Zn. Ca concentrations in all tissues were halved when Mg concentration increased eight-fold in the solution.
Keywords: Banana nutrition, propineb, mancozeb, micronutrients, magnesium.
Introducción
El aporte de microelementos por parte de algunos fungicidas elaborados a base de metales ha sido reportado como un efecto secundario de las aplicaciones de estos productos (Poh et ál. 2011). En el caso concreto de los fungicidas pertenecientes al grupo de los ditiocarbamatos, entre ellos el mancozeb y propineb, ha sido reconocida su capacidad para suplir Mn y Zn a las plantas deficientes en estos elementos (Deckers et ál. 1997, Agrios 2004).
Sin embargo, es escasa la información existente en cuanto a la influencia que tiene la aplicación repetida de estos productos y su consecuente aporte de microelementos, en la absorción de otros nutrientes esenciales para la planta, a pesar de que se ha documentado una interacción antagónica en la absorción entre elementos catiónicos con una misma valencia (Marschner 1995). Por ejemplo, en el caso del Mg es conocido su antagonismo con otros cationes divalentes (como el Mn), que al estar presentes en altas concentraciones en el medio de crecimiento pueden inhibir la absorción de este nutrimento (Merhaut 2007).
Al respecto, Quartin et ál. (2001) mencionan que altas concentraciones de Mn en el medio de crecimiento inhibe la absorción de Mg por parte de las raíces de algunos cultivares de trigo.
En Costa Rica, las plantaciones de banano se establecen principalmente en la Vertiente Atlántica, en donde es posible distinguir 2 regiones con marcadas diferencias edáficas debido al origen de los suelos. En la sección Este del Río Reventazón predominan los suelos con alto contenido de bases, formados a partir de aluviones de origen sedimentario y depositados por inundaciones frecuentes (Arias et ál. 2010).
Por otro lado, en la sección Oeste de la cuenca de este río predominan los suelos con propiedades ándicas, formados a partir de materiales volcánicos y donde los contenidos de bases intercambiables son mucho más bajos (López y Solís 1991, Méndez y Bertsch 2012). De acuerdo con López y Solís (1991), además de los bajos niveles de Mg presentes en el suelo, los desbalances con otros elementos como el K, han reportado con frecuencia, bajos contenidos foliares de este elemento en las plantaciones de banano de esa región. Una situación similar ocurre con el caso del Zn, el cual con frecuencia se presenta en bajos contenidos tanto en el suelo como en la planta.
Por lo tanto, con la ejecución del presente trabajo se pretende obtener información preliminar que permita identificar el efecto de varios ciclos de aplicación, de los fungicidas propineb y mancozeb sobre la nutrición de plántulas de banano (Musa sp.) en solución nutritiva, y más concretamente en lo que respecta al aporte de los elementos Zn y Mn y su repercusión en la relación Mg:Mn.
Materiales y Métodos
El experimento se realizó en el Centro de Investigaciones Agronómicas de la Universidad de Costa Rica (CIA-UCR) con plántulas de banano provenientes de cultivo in vitro, establecidas en solución nutritiva dentro de un invernadero durante enero a marzo del 2012.
Selección y preparación del material
Con el fin de garantizar la homogeneidad del material vegetal, las plántulas fueron trasplantadas en 2 recipientes de 20 l de capacidad y se sometieron a un período de aclimatación en medio hidropónico durante 3 semanas, con un cambio de solución nutritiva cada 7 días. La concentración de los elementos en la solución durante este período se indica en el Cuadro 1.
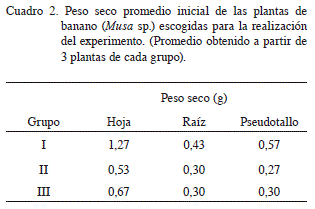
Al finalizar el período de aclimatación se seleccionaron las plantas que presentaron un mayor grado de uniformidad entre sí (Figura 1), y se establecieron 3 grupos (según su tamaño) que corresponderían posteriormente a cada uno de los bloques del experimento. De cada grupo se tomaron 3 plantas, a las cuales, después de permanecer por un período de 48 horas en una estufa a 80ºC, se les determinó el peso seco. En el Cuadro 2 se presenta el peso seco promedio inicial para las diferentes partes de la planta de cada uno de los grupos.
Una vez seleccionado el material que se utilizaría en el experimento se instalaron 3 plántulas por pote de 4 l de capacidad recubiertos con bolsas plásticas negras. Los potes fueron colocados sobre mesas de trabajo a una distancia inicial de 40 cm entre cada uno de ellos y se cubrieron con tapas de estereofón, las cuales sirvieron de soporte para colocar las plántulas sobre la solución nutritiva.
Aplicación de tratamientos
En total se aplicaron 12 tratamientos, producto de la combinación de 6 tratamientos foliares (Cuadro 3) con 2 diferentes concentraciones de los elementos Mg y Zn en la solución nutritiva (25 y 0,05 mg.l-1 de Mg y Zn respectivamente en una solución; 200 y 0,3 mg.l-1 de Mg y Zn en la otra solución). En ambos casos el Mn se mantuvo en niveles relativamente altos (24 mg.l-1).
La concentración de los demás elementos durante el tiempo en que se aplicaron los tratamientos, fue la misma que se presentó en el Cuadro 1. La solución nutritiva fue preparada a partir de sales solubles y únicamente para el caso del Fe, se utilizó una fuente quelatada de Fe-EDTA para evitar incompatibilidad con los otros elementos.
Durante las 2 primeras semanas de aplicación de tratamientos la solución nutritiva se cambió cada 7 días; después de este tiempo fue necesario realizar los cambios de solución cada 5 días debido a la mayor demanda de agua que presentaron las plantas conforme crecían.
Las aspersiones foliares se realizaron basándose en un volumen equivalente a 25 l.ha-1 de mezcla. El área de aplicación fue de 0,6 m2 (6 potes por tratamiento: 3 repeticiones x 2 soluciones) lo que correspondió con un volumen de 1,5 ml de solución. Para lograr una aspersión uniforme con un volumen tan bajo de mezcla se utilizó un aerógrafo marca Badger modelo 200, con base en experiencias previas. Se calibró el aparato para conocer la descarga por minuto y así ajustar el tiempo de aplicación que permitió una cobertura uniforme del follaje con el volumen de mezcla requerido. En total se realizaron 8 ciclos de aplicación con una separación de 4 días entre cada uno de ellos.
A partir del sexto ciclo, fue necesario reajustar el volumen de aplicación debido a la expansión del área foliar de las plantas, al ser necesario aplicar 3,5 ml de solución para los 6 potes correspondientes de cada tratamiento foliar.
Los fungicidas utilizados se caracterizan por contener en sus moléculas de ingrediente activo las siguientes cantidades de elementos menores: mancozeb entre 20 y 22% de Mn y 2,5% de Zn, y el propineb alrededor de 16% de Zn (Lo et ál. 1996, Weingerl y Kerin 2000).
Para el caso del tratamiento foliar 3 (prop/ man) se alternó el uso de propineb y mancozeb de ciclo por medio, por lo que se realizaron 4 aplicaciones de cada producto. En el tratamiento 5 (man+Zn), las aplicaciones con una fuente adicional de Zn se realizaron de ciclo por medio y se utilizó un quelato con aminoácidos con una concentración de 6,8% de este elemento, a una dosis equivalente a 0,5 l.ha-1 de producto comercial (según recomendación de la etiqueta del producto).
Diseño experimental y análisis de datos
En el experimento se empleó un diseño de bloques completos al azar y se conformaron 3 bloques, dentro de los cuales se distribuyó de manera aleatoria los 12 tratamientos (producto del factorial 6 x 2 mencionado anteriormente). La conformación de los bloques se basó en el tamaño y la homogeneidad de las plantas al momento de iniciar la aplicación de tratamientos, según lo expuesto en el Cuadro 2. Cada unidad experimental estuvo constituida por un pote con 3 plantas.
Semanalmente se realizaron evaluaciones visuales del estado en el que se encontraban las plantas con el fin de identificar la aparición de síntomas de deficiencias nutricionales o bien, síntomas por posibles toxicidades o “quemas” debido a aplicación reiterada de los productos foliares.
Para evaluar la concentración y la absorción total de los elementos, 4 días después del último ciclo de aplicación se cosecharon todas las plantas y se separaron en raíz, pseudotallo y láminas foliares. A cada una de estas partes se le determinó el peso fresco y luego de que permanecieron en una estufa a 80°C por 48 h se les determinó el peso seco. Los análisis para determinar la concentración de nutrientes en los diferentes tejidos fueron realizados en el Laboratorio de Suelos y Foliares del CIA-UCR, mediante una digestión del material vegetal con HNO3 y analizándolo por medio de un equipo de Espectroscopía de Emisión Atómica con plasma (ICP), por medio de los procedimientos establecidos para este tipo de determinaciones descritos por Díaz y Hunter (1978) y Henríquez et ál. (1995).
El análisis estadístico se hizo mediante el programa SigmaPlot 11.0; con el cual se realizó un análisis de varianza (p≤0,05) para determinar la existencia de diferencias significativas y una prueba de Diferencia Mínima Significativa (DMS) de Fisher (p≤0,05) para realizar la separación estadística de medias.
Resultados y Discusión
Concentración y absorción de Zn
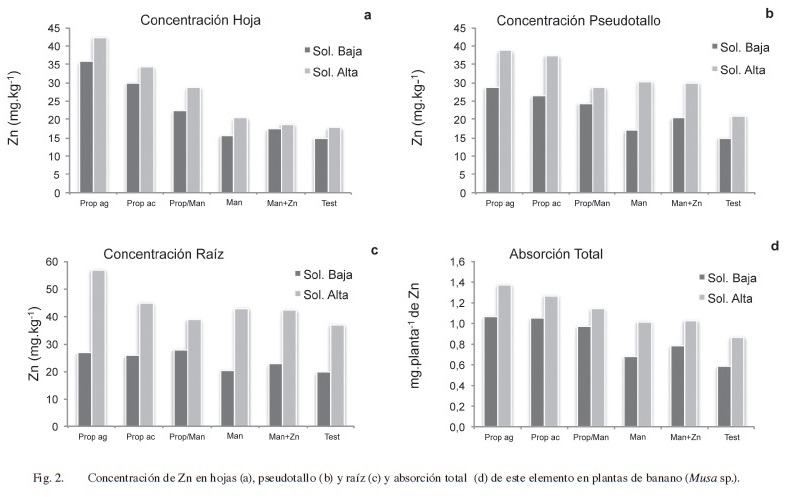
En la hoja, no hubo interacción (p<0,05) de los tratamientos foliares con la solución nutritiva, lo cual indica que el efecto de las aspersiones fue similar, independientemente del nivel de Zn en la solución. Hubo diferencias significativas (p<0,05) entre los tratamientos que incluyeron propineb en aplicaciones continuas (8 ciclos), que pasaron de un promedio de 16 mg.kg-1 en el tratamiento testigo a 32 y 39 mg.kg-1 de Zn, o sea el doble, en los tratamientos de propineb en aceite y en agua, respectivamente. No hubo diferencias significativas (según prueba DMS p<0,05) entre estos 2 últimos tratamientos (Figura 2a). Los tratamientos que incluyeron mancozeb no difirieron del testigo (p<0,05) en la concentración foliar de Zn. No obstante, a pesar de que cuando este producto se alternó con propineb (prop/man) se presentó un incremento en el contenido de Zn, este aumento no alcanzó a ser significativo.
A nivel de pseudotallo, al igual que lo ocurrido en las hojas, con las aplicaciones de propineb tanto en agua como en aceite, se obtuvieron las mayores concentraciones de Zn. En el caso de los tratamientos propineb alternado con mancozeb (prop/man) y mancozeb con Zn foliar (man+Zn) se encontraron diferencias significativas (p<0,05), y estas fueron únicamente con respecto al testigo. Además, se hallaron diferencias significativas (p<0,05) entre las 2 soluciones nutritivas, obteniéndose mayores concentraciones de Zn con la solución que contenía niveles más elevados de este elemento, independientemente del tratamiento foliar aplicado (Figura 2b).
En la raíz no hubo diferencias entre los tratamientos asperjados y únicamente se hallaron diferencias significativas entre las 2 soluciones con diferentes niveles de Zn y Mg (p<0,05). Se obtuvo una concentración promedio de Zn de 43,8 mg.kg-1 en los casos donde la solución contenía los niveles más elevados de Zn, en comparación con la concentración promedio de 23,8 mg.kg-1obtenida cuando la solución contenía niveles bajos de Zn.
En las Figuras 2b y 2c se puede observar que el efecto de la concentración de Zn en el tejido por causa de las diferencias en la solución nutritiva, es más marcado en la raíz que en el pseudotallo.
Respecto a la absorción total de Zn, en la Figura 2d se observa que hubo diferencias significativas (p<0,05) entre los diferentes tratamientos asperjados y entre las soluciones nutritivas, sin embargo el comportamiento de los tratamientos asperjados presentó una tendencia similar en ambas soluciones, por lo que no se halló una interacción entre tratamientos (p<0,05). Entre los tratamientos foliares, los que contienen propineb presentaron las mayores cantidades de Zn absorbido, sin diferencias significativas entre sí. Sin embargo solo los tratamientos que recibieron las aplicaciones consecutivas de este producto en agua y aceite durante los 8 ciclos que duró el ensayo presentaron valores estadísticamente diferentes con respecto al resto de tratamientos.
Los niveles más altos de Zn en el medio propiciaron una mayor absorción de este elemento en términos totales por parte de la planta, independientemente de la aspersión que se realizara, lo cual refleja que la aplicación del producto foliar no restringió la absorción radical de Zn, si este estuvo disponible en el medio, sin embargo su translocación al resto de la planta no fue homogénea y por el contrario tendió a acumularse en el tallo y en mayor medida en la raíz.
Estos resultados reflejan la capacidad que tiene el propineb de aportar Zn al ser utilizado de manera reiterada en varios ciclos de aplicación, independientemente de si la concentración en la solución nutritiva fue alta o baja. Esta capacidad de aporte de Zn por parte del propineb ha sido también reportada por Tiancang et ál. (2008), quienes al evaluar la efectividad de este producto para controlar el hongo Diplocarpon mali en árboles de manzana, observaron un incremento considerable en el contenido foliar de Zn de 20,9 a 75 mg.kg-1 luego de haber realizado 2 ciclos de aplicación.
El propineb es un fungicida que contiene un 16% de Zn (con base en la formulación del producto comercial) mientras que en el mancozeb tiene una concentración de apenas 2,5% de este elemento (Lo et ál. 1996, Weingerl y Kerin 2000). La diferencia en el contenido de Zn de estos 2 fungicidas es una de las principales razones por la cual se presentaron las diferencias observadas entre tratamientos foliares en las hojas y pseudo-tallo. De acuerdo con Fernández y Eichert (2009) entre los factores concernientes a las propiedades físico-químicas de la solución, la concentración externa de solutos es una de las que más influencia tiene en la tasa de penetración a través de la superficie foliar.
Por esta misma razón, se podría explicar el hecho de que al añadir una fuente adicional de Zn con una concentración más baja de este elemento (6,8%) y aplicada de manera alterna en los ciclos, no se reflejara significativamente con cambios en la concentración de Zn en la hoja, y solo de manera leve en el pseudotallo. Se debe anotar sin embargo, que la aplicación de dicho tratamiento se hizo de esta forma con el fin de simular de la mejor manera posible las prácticas que se realizan en plantaciones comerciales, y siguiendo la dosis recomendada del distribuidor de la fuente de Zn adicional.
A pesar de que es conocido el efecto que tienen los aceites agrícolas y los agentes surfactantes en el mejoramiento de la penetración de moléculas a través de la cutícula de las hojas, debido al aumento en la permeabilidad que sufre la misma ante la presencia de este tipo de sustancias (Stock y Holloway 1993, Fernández y Eichert 2009); para efectos de absorción de Zn en el presente ensayo, el agente utilizado para la aplicación de los productos (agua o emulsión agua:aceite) no presentó influencia significativa en los tratamientos con propineb. Sin embargo, es importante resaltar nuevamente que el experimento fue realizado en un ambiente controlado en ausencia de precipitación, condición en la cual se ha justificado el uso de aceites y surfactantes para mejorar la acción de los productos aplicados foliarmente.
Translocación del Zn
El Zn ha sido descrito en diversas ocasiones como un elemento de movilidad intermedia dentro de la planta, sin embargo su tasa de translocación puede variar según la especie y las condiciones propias del momento (disponibilidad, demanda, interacción con otros elementos y compuestos) (Bukovac y Wittwer 1957, Lindsay 1972, Kochian 1991).
Los resultados muestran que parte del Zn absorbido a través de las hojas fue transportado al pseudotallo de la planta, ya que en este tejido fue posible observar diferencias significativas (p<0,05) con los tratamientos que llevaron aplicaciones consecutivas de propineb. Además en el caso del tratamiento man+Zn, se observó también un ligero aumento, con respecto al tratamiento testigo, en la concentración de Zn en el pseudotallo (Figura 2b), esto a pesar de que en la hoja no hubieron diferencias, lo cual hace pensar que parte del Zn aplicado foliarmente fue movilizado. Estos resultados contrastan con lo citado por Vargas y Solís (1999) y por Arias et ál. (2006) quienes mencionan que la movilidad del Zn en plantas de banano es muy limitada.
Basándose en los resultados de este experimento, se puede decir que además de la movilización acrópeta del Zn vía xilema, la cual se refleja en el incremento en el contenido de Zn en la raíz y el pseudotallo al aumentar la concentración de este elemento en la solución, se dio una movilización de tipo basípeta, vía floema (de las hojas al pseudotallo) en el caso de los tratamientos foliares que contenían propineb aplicado de forma consecutiva.
Esta capacidad de transporte del Zn vía floema ha sido confirmada en otras especies; por ejemplo Haslett et ál. (2001) encontraron que en plantas de trigo hubo movilización de este elemento por el floema a partir de las hojas asperjadas con fuentes inorgánicas (ZnO y ZnSO4) y quelatadas hacia las partes inferiores de la planta, al igual que lo hicieron en su momento Hocking (1980) en tabaco y Volschenk et ál. (1999) en viñedos.
Fageria et ál. (2009) mencionan que la removilización de un nutriente dentro de la planta es esencial para que el efecto de la aplicación sea exitoso, ya que si este es inmovilizado después de su aplicación su efecto positivo estaría limitado únicamente a los tejidos asperjados con la solución.
A pesar de que en el pseudotallo se evidenció un aumento en la concentración de Zn al incrementarse la concentración de este elemento en la solución nutritiva, fue en la raíz donde la mayoría del Zn absorbido fue retenido, lo cual puede estar relacionado con la capacidad de este tejido de acumular minerales dentro de sus células (Marschner 1995). Las raíces suelen contener niveles más elevados de Zn que las partes aéreas, especialmente si las plantas crecen en medios con alta disponibilidad de este elemento. Además las plantas usualmente acumulan niveles elevados de Zn en las raíces si el suplemento de este es adecuado; si por el contrario, crecen en condiciones de deficiencia, el Zn es redistribuido y utilizado por los tejidos aéreos (Lindsay 1972), lo cual se puede ver reflejado en la Figura 2b y 2c, al observar que fue en la raíz donde mayor diferencia hubo en la concentración de Zn al pasar de niveles bajos en la solución nutritiva, a niveles relativamente altos de Zn.
Por lo tanto, basándose en los resultados obtenidos en este experimento se tiene que las aspersiones de productos que contienen Zn no suprimen la absorción de este elemento por parte de las raíces, y que por el contrario sirven como un complemento para suplir de Zn a la planta en el caso de que este no sea eficientemente translocado desde la raíz.
Concentración y absorción de Mg
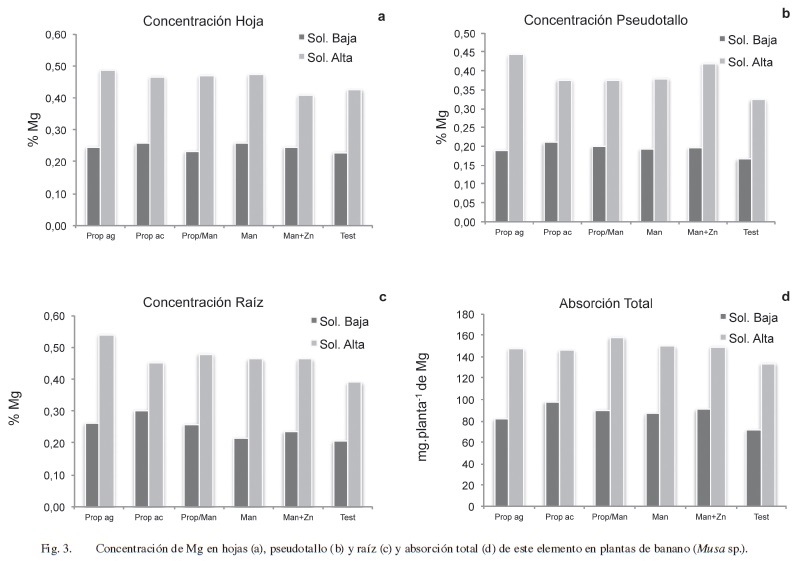
Con base en los resultados obtenidos, no se encontró efecto alguno de la aplicación de los 2 fungicidas evaluados (propineb y mancozeb) sobre el comportamiento del Mg en la planta, ya que no hubo diferencias significativas (p<0,05) entre los diferentes tratamientos foliares en concentración ni en la absorción de este nutrimento (Figura 3).
Los resultados de este ensayo concuerdan con los encontrados por Fontes et ál. (1999) quienes al realizar aplicaciones foliares con fungicidas que contienen Zn determinaron que se dio un incremento en la concentración de este elemento en hojas de papa sin que hubiera algún efecto en el comportamiento del Mg. De igual forma Kowalenko e Ihnat (2010) encontraron que al realizar aplicaciones foliares de Zn y Mn en coliflor se incrementó la concentración de estos elementos en el tejido, sin que la absorción de algún otro nutrimento se viera afectada.
Sin embargo al variar la concentración de Mg en la solución nutritiva de 25 a 200 mg.l-1 sí fue posible observar un incremento significativo (p<0,001) en la concentración de este elemento en las 3 partes de la planta evaluadas, que pasa de concentraciones promedio (con base en la cantidad de materia seca) de 0,26 a 0,45% en las hojas, de 0,19 a 0,39% en el pseudotallo y de 0,25 a 0,47% en la raíz (Figura 3a,b,c).
En términos de absorción total de Mg, se observó que, a nivel general hubo un incremento de hasta un 70% en la cantidad total de este elemento absorbido por las plantas de banano al aumentar 8 veces la concentración de Mg en la solución nutritiva (Figura 3d). Marschner (1995) menciona que al incrementar el suplemento de Mg más allá de los niveles requeridos para un adecuado crecimiento es común que se dé un incremento en la cantidad de Mg almacenado en las vacuolas, el cual actúa como un “buffer” y como un criterio de compensación de cargas y osmorregulación de la vacuola.
Al usar una concentración moderadamente alta de Mn en la solución nutritiva (24 mg.l-1) y niveles de Mg (Mg bajo con una relación Mg:Mn 1:1 y Mg alto con una relación Mg:Mn 8:1) se esperaba observar un efecto antagónico diferenciado entre estos 2 elementos, según la relación Mg:Mn, al agregar de manera reiterada cantidades adicionales de Mn y Zn provenientes del mancozeb y propineb respectivamente; esto debido a que tanto el Mn+2 como el Zn+2 son cationes divalentes que pueden competir con el Mg+2 por los sitios de intercambio en las partículas del complejo coloidal del suelo y en las membranas biológicas a lo interno de la planta (Merhaut 2007). Sin embargo, en este caso, no se observó efecto alguno sobre el Mg por parte de estos 2 elementos al ser aplicados foliarmente.
A pesar de que las plantas requieren cantidades mucho menores de estos micronutrientes en comparación con el Mg, algunos estudios han mostrado su capacidad de disminuir la absorción y acumulación del Mg cuando estos se encuentran disponibles en cantidades lo suficientemente elevadas (Gunes et ál. 1998, Mills y Bones 1996); sin que esto signifique necesariamente la aparición de síntomas por deficiencia de Mg (Merhaut 2007).
Por ejemplo, Heenan y Campell (1981) encontraron que el Mn podía competir eficientemente con el Mg, debido a la interacción efectiva del Mn a nivel de la membrana plasmática de las raíces y por el bloqueo de los sitios de transporte del Mg. Por su parte Kaya et ál. (2001) encontraron que a concentraciones extremadamente altas de Zn (30 mg.l-1) la acumulación de Mg en plantas de tomate (frutos y hojas) fue inhibida, en comparación con plantas sometidas a concentraciones normales de Zn (0,5 mg.l-1).
En plantas de banano Turner y Barkus (1983) reportaron el antagonismo entre estos 2 elementos al evaluar las plantas que crecían en medios con altas concentraciones de Mn. Estos autores encontraron que se dio un incremento de 7 veces en la absorción del elemento Mn, mientras que la absorción de Mg se vio significativamente reducida.
Por lo tanto, respaldándose con los resultados obtenidos en este ensayo y con lo reportado por los autores anteriormente citados, se podría esperar que la disminución en la absorción de Mg debido al efecto antagónico del Zn y del Mn se presente principalmente cuando estos 2 últimos elementos se encuentran disponibles en grandes cantidades para ser absorbidos por la raíz, y no así por medio de la hojas, ya que usualmente las cantidades aplicadas y la capacidad de absorción foliar son mucho menores.
Sin embargo, es importante anotar que, a largo plazo, es probable que el uso reiterado durante muchos ciclos de aplicación de fungicidas pueda llegar a afectar la absorción de otros elementos catiónicos como el Mg, especialmente si en el suelo se presentan condiciones que facilitan la solubilidad del Mn, situación que ocurre con frecuencia en muchos suelos bananeros de Costa Rica. Según Arias (2002) en plantaciones bananeras es frecuente que se apliquen entre 8 y 10 kg.ha-1.año-1 de Mn foliar debido al uso reiterado de productos como el mancozeb; esta dosis es mucho mayor a la recomendada para corregir deficiencias de Mn en banano. Moreira (1999) señala que el Mn suplido foliarmente es absorbido de manera eficiente, lo cual puede conducir a una acumulación progresiva si es aplicado de manera constante.
Efecto sobre la concentración de Mn y la relación Mg:Mn a nivel de planta
En términos de concentración en los diferentes tejidos analizados (hoja, pseudotallo y raíz) y de absorción total de Mn, no fue posible observar diferencias estadísticamente significativas (p<0,05) producto de la aplicación de los diferentes tratamientos foliares, esto a pesar de que se observó una tendencia a aumentar la concentración en el tejido foliar del tratamiento con aplicaciones reiteradas de mancozeb.
Sin embargo, tal y como se mencionó anteriormente, la competencia entre el Mg y Mn ha sido ampliamente documentada (Marschner 1995, Merhaut 2007), encontrándose no solamente una disminución en la absorción de Mg al haber altas concentraciones de Mn en el medio de crecimiento, sino que también se puede dar una disminución en la absorción de Mn si el Mg está presente en altas concentraciones.
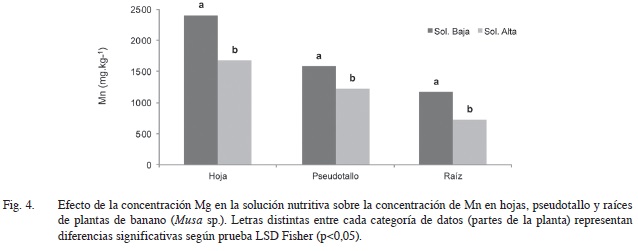
En el presente ensayo, la absorción de Mn se vio afectada significativamente (p<0,05) por la concentración de Mg en la solución nutritiva, ya que al aumentar los niveles de este último elemento en el medio de crecimiento la concentración de Mn disminuyó en todos los tejidos de la planta analizados. En la lámina foliar con mayor magnitud, en términos absolutos, se evidenció esta diferencia al pasar de una concentración promedio de 2398 mg.kg-1 cuando los niveles de Mg eran bajos (25 mg.l-1) a 1676 mg.kg-1 cuando se incrementó la cantidad de Mg en la solución (200 mg.l-1) (Figura 4). Sin embargo en términos porcentuales la raíz fue la parte de la planta que mayor reducción en la concentración de Mn tuvo al incrementar los niveles de Mg en la solución, presentándose una disminución del 38% con respecto a la condición encontrada cuando la cantidad de Mg era baja en la solución.
A pesar de esa disminución en el contenido de Mn, visualmente no se encontró ningún síntoma de posible deficiencia de este elemento, ya que aún en altas concentraciones de Mg en la solución nutritiva los niveles de Mn a nivel foliar se mantuvieron por encima de 1500 mg.kg-1 de materia seca, lo que corresponde a concentraciones muy superiores a las reportadas como el mínimo requerido para plantas de banano, que se estima en aproximadamente entre 23 y 25 mg.kg-1 (Arias 2002, Soto 2008).
Por el contrario, con esas concentraciones de Mn en la lámina foliar se han reportado la aparición de síntomas por toxicidad con este elemento. Vargas (2001) encontró síntomas por toxicidad de Mn en plantas de plátano (Musa AAB cv. Falso cuerno) cultivadas en suelo cuando la concentración de Mn foliar superó los 1400 mg.kg-1. Sin embargo en este experimento tampoco se observaron síntomas por toxicidad con Mn, lo cual puede deberse a la capacidad que tienen las plantas de banano para tolerar altos contenidos de Mn (Vargas 2001).
Al respecto Clarkson (1988) citado por Kochian (1991) menciona que la absorción de Mn no parece tener un mecanismo de control definido como ocurre en el caso de los macronutrientes, estimándose en algunas ocasiones que la absorción de Mn puede ser entre 100 y 1000 veces mayor que la necesidad real de las plantas, sin que se presenten alteraciones en el funcionamiento fisiológico de la planta.
Los resultados obtenidos en este ensayo concuerdan con lo reportado por otros autores, por ejemplo Elamin y Wilcox (1986a), quienes encontraron que al incrementar los niveles de Mg en la solución nutritiva la concentración de Mn en tallos y raíces descendió al tiempo que los síntomas por toxicidad de Mn disminuyeron en plantas de melón, y se obtuvo un incremento en la producción de biomasa. Estos mismos autores (Elamin y Wilcox 1986b) que trabajan con plantas de sandía encontraron resultados similares a los anteriores.
Arias (2002) menciona que al aplicar Mg en el cultivo de banano se encontró una disminución entre el testigo y la mayor dosis de Mg aplicada (300 kg.ha-1.año-1) de 120 mg.kg-1 en la concentración de Mn foliar (en la tercer hoja). Turner y Barkus (1980) al trabajar con plantas de banano, encontraron una disminución en la cantidad de materia seca producida al someterse la planta a niveles de Mg bajos y de Mn altos, aunque su efecto no fue tan contundente como el que se observó al tener las plantas con bajos niveles de K.
Por su parte, Le Bot et ál. (1990) trabajó con plantas de tomate y trigo para derminar que al incrementar los niveles de Mg no solo hubo disminución en la absorción de Mn, sino que aún al presentarse altas concentraciones de Mn ante la presencia también de altos niveles de Mg en el tejido, se dio una disminución en los problemas de toxicidad causados por el Mn. Por esta razón estos autores concluyen que la relación Mg:Mn es un mejor indicador del estado nutricional del Mn en la planta, que el dato de concentración de Mn por sí mismo.
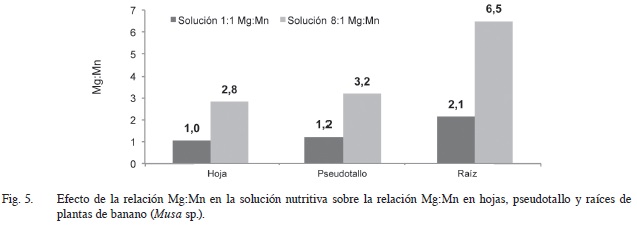
En la Figura 5 se muestra como al pasar de una relación 1:1 de Mg:Mn en la solución nutritiva a una proporción 8:1 se incrementa también el valor de dicho cociente a nivel de tejido. No obstante, este incremento no se da de manera proporcional en todas las partes de la planta, ya que es más acentuado en la raíz en donde la relación Mg:Mn pasa de 2,1 a 6,5 al incrementarse la concentración de Mg en 8 veces en la solución. Estos resultados están directamente asociados tanto al aumento en la absorción de Mg al incrementar su concentración en la solución nutritiva como a la disminución en la absorción de Mn en los diferentes órganos de la planta (Figuras 3 y 4).
La mayor variación en la relación Mg:Mn en la raíz indica que al incrementar la concentración de Mg en el medio de crecimiento gran parte del Mn fue retenido en este tejido, lo cual se puede asociar con la capacidad que tiene este órgano para “retener” elementos cuando se encuentran en grandes cantidades, y de esta forma mantener cierto equilibrio en las demás partes de la planta de manera que no se vean afectados procesos fisiológicos fundamentales. Humphries et ál. (2007) han reportado que en ocasiones la tolerancia a altas concentraciones de Mn está relacionada con la reducción en el transporte de Mn desde la raíz hasta el tallo, que incrementó la retención de Mn en la raíz. También, Andrew y Hegarty (1969) al comparar entre varias especies de leguminosas concluyeron que la tolerancia a la toxicidad de Mn fue atribuible parcialmente a la capacidad de retener el exceso de Mn en el sistema radical.
A nivel fisiológico, Marschner (1995) indica que en diversos estudios de activación enzimática llevados a cabo in vitro se ha encontrado que el Mn ha sido reemplazado por el Mg, o viceversa. El Mn está involucrado en diversas funciones bioquímicas, principalmente como activador de diversas enzimas envueltas en la respiración, en la síntesis de aminoácidos y ligninas y concentración de hormonas, además de su participación en reacciones de Redox en la cadena de transporte de electrones y en el desdoblamiento de la molécula de agua en el proceso fotosintético (Reacción de Hill), procesos en los cuales puede ser inhibido por la acción del Mg (Humphries et ál. 2007).
También, hay ocasiones en las que el exceso de Mn puede inhibir la acción del Mg, tal y como ocurre con el caso de la activación de la rubisco (RuBP), en el cual el Mn puede desplazar el Mg e inhibir la reacción de carboxilación de esta enzima, y como resultado una disminución en la tasa neta fotosintética. De allí la importancia de mantener un equilibrio entre estos elementos especialmente a nivel foliar. Esta manera de regular el balance entre estos elementos se puede ver reflejado en la Figura 5, en la que se muestra como en las hojas, a pesar de que en la solución nutritiva el incremento fue de 8 unidades, la relación Mg:Mn se incrementó apenas en 1,8 unidades (de 1 a 2,8).
Efecto sobre la producción de biomasa y en la absorción de otros nutrimentos
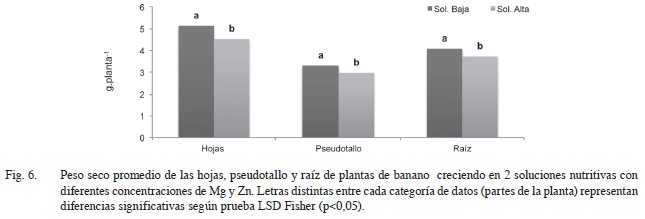
En términos de producción de biomasa, no se encontraron diferencias significativas entre los tratamientos foliares en el peso seco aéreo y radical de las plantas de banano tanto en el caso de los tratamientos sometidos a bajas concentraciones de Mg y Zn , como a los que se encontraban a concentraciones altas de estos elementos.
Entre soluciones nutritivas sí se presentaron diferencias significativas (p<0,05), obteniéndose el mayor peso seco en el caso de la solución con bajos niveles de Mg y Zn, y fue en la parte aérea (principalmente en las hojas) donde esa diferencia fue mayor (Figura 6).
Una razón por la que se pudo dar esta diferencia en la cantidad de materia seca producida es debido a la interacción (antagonismo) con otros elementos que se dio al incrementar las concentraciones de Mg y Zn en la solución nutritiva.
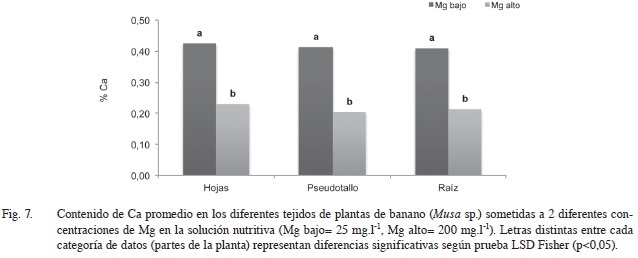
En la Figura 7 se evidencia que la absorción de Ca se redujo prácticamente a la mitad (de 0,41% a 0,21% en promedio en relación con la cantidad de materia seca) en todos los tejidos de la planta al incrementarse el contenido de Mg en la solución nutritiva. Esta relación antagónica entre estos 2 elementos ha sido frecuentemente reportada, y de acuerdo con Merhaut (2007) las altas concentraciones de Ca en la rizósfera inhiben la absorción de Mg y viceversa, presentándose una disminución en el crecimiento de las plantas en ambos casos.
La disminución en la cantidad de materia seca al disminuir la concentración de Ca en los tejidos es probable que se deba al rol estructural que tiene el pectato de Ca en la lámina media de la pared celular y a su participación activa en los procesos de extensión y división celular (Marschner 1995).
Además la concentración de Zn y Mg en la solución nutritiva no interfirió con los tratamientos aplicados foliarmente, ya que tanto en concentraciones altas como bajas de estos elementos se observó el mismo efecto de los productos asperjados vía foliar.
Conclusiones
El uso reiterado del fungicida propineb incrementó la concentración de Zn en la parte aérea de las plantas de banano, independientemente de si es aplicado en agua o en emulsión.
No se encontraron diferencias significativas en el efecto de la aplicación de tratamientos foliares sobre la concentración de Mn, sin embargo es probable que estos resultados se deban a la alta concentración de este elemento en la solución nutritiva. Tampoco hubo efecto en la concentración de Mg en ninguna de las partes analizadas de la planta, por lo que se podría decir que no hubo un efecto antagónico de los elementos Zn y Mn (presentes en el propineb y mancozeb) sobre el elemento Mg al ser asperjados foliarmente.
Al elevarse la concentración de Mg en la solución nutritiva sí se observó un efecto antagónico con el Mn y el Ca, ya que su concentración se vio disminuida considerablemente en todos los tejidos de la planta al incrementarse los niveles de Mg en la solución nutritiva. Además, al aumentar la relación Mg:Mn en la solución nutritiva, se incrementó la relación Mg:Mn en los tejidos, principalmente en la raíz, foliarmente el cambio fue muy moderado lo que parece indicar que la raíz contribuyó a atenuar ese cambio.
Finalmente, es importante anotar que debido a las condiciones de invernadero y solución nutritiva en las que se realizó el ensayo, muestran resultados que deben ser considerados como preliminares y por lo tanto no deben ser extrapolados directamente a campo, donde las condiciones climáticas presentan grandes variaciones que afectan la efectividad de los productos y la absorción de los nutrientes.
Literatura citada
AGRIOS G. 2004. Plant Pathology. 5 ed. Elsevier Academic Press. New York, USA. 922 p. [ Links ]
ANDREW C., HEGARTY M. 1969. Comparative responses to manganese excess of eight tropical and four temperate pasture legume species. Australian Journal of Agricultural Research 20(4):687-696. [ Links ]
ARIAS F. 2002. Análisis y comentario: el manganeso (Mn) en la nutrición del cultivo de banano (Musa AAA). CORBANA 28(55):83-96. [ Links ]
ARIAS F., MATA R., ALVARADO A., SERRANO E., LAGUNA J. 2010. Caracterización química y clasificación taxonómica de algunos suelos cultivados con banano en las llanuras aluviales del Caribe de Costa Rica. Agronomía Costarricense 34(2):175-195. [ Links ]
ARIAS F., SERRANO E., GUZMAN M. 2006. Absorción y translocación de Zinc asperjado al follaje de plantas jóvenes de banano (Musa AAA. CV. Grande Naine), desarrolladas en cultivo hidropónico. CORBANA 32(59):79-85. [ Links ]
BUKOVAC M., WITTWER S. 1957. Absorption and mobility of foliar applied nutrients. Plant Physiology 32(5):428-435. [ Links ]
DECKERS T., DAEMEN E., LEMMENS K., MISSOTTEN C. 1997. Influence of foliar applications of Mn during summer on the fruit quality of Jonagold. Acta Horticulturae 448:467-474. [ Links ]
DÍAZ R., HUNTER A. 1978. Metodologías de muestreo de suelos, análisis químico de suelos y tejido vegetal y de investigación en invernadero. CATIE. Turrialba, Costa Rica, 62 p. [ Links ]
ELAMIN O., WILCOX G. 1986a. Effect of magnesium and manganese nutrition on muskmelon growth and manganese toxicity. Journal of the American Society for Horticultural Science 111(4):582-587. [ Links ]
ELAMIN O., WILCOX G. 1986b. Effect of magnesium and manganese nutrition on watermelon growth and manganese toxicity. Journal of the American Society for Horticultural Science 111(4):588-593. [ Links ]
FAGERIA N.K., BARBOSA M.P., MOREIRA A., GUIMARAES M. 2009. Foliar fertilization of crops plants. Journal of Plant Nutrition 32:1044-1064. [ Links ]
FERNÁNDEZ V., EICHERT T. 2009. Uptake of hydrophilic solutes through plant leaves: current state of knowledge and perspectives of foliar fertilization. Critical Reviews in Plant Sciences 28(1):36-68. [ Links ]
FONTES P.C.R., MOREIRA M.A., FONTES R.L.F., CARDOSO A.A. 1999. Effects of zinc fungicides and different zinc fertilizer application methods on soluble and total zinc in potato plant shoots. Communications in Soil Science and Plant Analysis 30(13-14):1847-1859. [ Links ]
GUNES A., ALPASLAN M., INAL A. 1998. Critical nutrient concentrations and antagonistic and synergistic relationships among the nutrients of NFT-grown young tomato plants. Journal of plant nutrition 21(10):2035-2047. [ Links ]
HASLETT B.S., REID R.J., RENGEL Z. 2001. Zinc mobility in wheat: uptake and distribution of zinc applied to leaves or roots. Annals of Botany 87(3):379-386. [ Links ]
HEENAN D., CAMPBELL L. 1981. Influence of potassium and manganese on growth and uptake of magnesium by soybeans ( (L.) Merr. Cv. Bragg). Plant and Soil 61(3):447-456. [ Links ]
HENRÍQUEZ C., BERTSCH F., SALAS R. 1995. Fertilidad de suelos: Manual de laboratorio. ACCS. San José, Costa Rica. 64 p. [ Links ]
HOCKING P.J. 1980. The composition of phloem exudate and xylem sap from tree tobacco ( Grah.). Annals of Botany 45:633-643. [ Links ]
HUMPHRIES J.M., STANGOULIS J.C.R., GRAHAM R.D. 2007. Manganese, pp. 351-374. In: A. Barker y D. Pilbeam (eds.). Handbook of Plant Nutrition. CRC Press. [ Links ]
KAYA C., BURTON M., HIGGS D. 2001. Responses of tomato cvs., grown to fruit–harvest stage under zinc stress in glasshouse conditions. Journal of Plant Nutrition 24(2):369-382. [ Links ]
KOCHIAN L.V. 1991. Mechanisms of micronutrient uptake and translocation in plants, pp. 229-296. In: J.J. Mortvedt, F.R. Cox, L.M. Shuman and R.M. Welch (eds.). Micronutrients in Agriculture, ed. 2. Soil Science Society of America. [ Links ]
KOWALENKO C., IHNAT M. 2010. Effects of soil limestone and foliar manganese and zinc applications on field cauliflower growth and element contents. Canadian Journal of Soil Science 90(1):177-188. [ Links ]
LE BOT J., GOSS M., CARVALHO M.J., BEUSICHEM M.L., KIRKBY E. 1990. The significance of the magnesium to manganese ratio in plant tissues for growth and alleviation of manganese toxicity in tomato (Lycopersicon esculentum) and wheat (Triticum aestivum) plants. Plant and Soil 124(2):205-210. [ Links ]
LINDSAY W. 1972. Zinc in soils and plant nutrition. Advances in Agronomy 24:147-186. [ Links ]
LO C.C., HO M.H., HUNG M.D. 1996. Use of high-performance liquid chromatographic and atomic absorption methods to distinguish Propineb, Zineb, Maneb, and Mancozeb fungicides. Journal of Agricultural and Food Chemistry 44(9):2720-2723. [ Links ]
LOPEZ M., SOLIS P. 1991. Contenidos e interacciones de los nutrimentos en tres zonas bananeras de Costa Rica. CORBANA 15(36):25-32. [ Links ]
MARSCHNER H. 1995. Mineral nutrition of higher plants. Academic Press. Cambridge, UK. 889 p. [ Links ]
MÉNDEZ J.C., BERTSCH F. 2012. Guía para la interpretación de la fertilidad de los suelos de Costa Rica. ACCS, San José, Costa Rica. 108 p. [ Links ]
MERHAUT D. 2007. Magnesium, pp. 146-181. In: A. Barker y D. Pilbeam (eds.). Handbook of Plant Nutrition. CRC Press. [ Links ]
MILLS H.A., BONES J.B. 1996. Plant Analysis Handbook II. Athens, GA: MicroMacro Publishing, Inc. 422 p. [ Links ]
MOREIRA R.S. 1999. Banana: Teoria e practica de cultivo. 2 ed. Fundaçao Cargill, São Pablo, Brasil. 335 p. [ Links ]
POH B.L., GEVENS A., SIMONNE E., SNODGRASS C. 2011. Estimating copper, manganese and zinc micronutrients in fungicide applications. UF/IFAS, University of Florida. Consultado el 24-07-2012. Disponible en http://edis.ifas.ufl.edu/hs1159. [ Links ]
QUARTIN V.M.L., ANTUNES M.L., MURALHA M.C., SOUSA M.M., NUNES M.A. 2001. Mineral imbalance due to manganese excess in triticales. Journal of plant nutrition 24(1):175-189. [ Links ]
SOTO M. 2008. Bananos. Técnicas de Producción, Manejo Postcosecha y Comercialización. Litografía e Imprenta LIL. San José, Costa Rica. 1090 p. [ Links ]
STOCK D., HOLLOWAY P.J. 1993. Possible mechanisms for surfactant-induced foliar uptake of agrochemicals. Pesticide Science 38:165-177. [ Links ]
TIANCANG Z., HUA Z., LILI H., HUANMIN X., DONGZI Z., JINGJING C. 2008. Efficacy of propineb for controlling leaf blotch caused by Marssonina coronaria and its effect on zinc content in apple leaves. Acta Phytophylacica Sinica 35(6):519-524. [ Links ]
TURNER D.W., BARKUS B. 1980. Plant growth and dry-matter production of the ‘Williams’ banana in relation to supply of potassium, magnesium and manganese in sand culture. Scientia Horticulturae 12(1):27-45. [ Links ]
TURNER D.W., BARKUS B. 1983. The uptake and distribution of mineral nutrients in the banana in response to supply of K, Mg and Mn. Fertilizer Research 4:89-99. [ Links ]
VARGAS A. 2001. Descripción y observaciones de la toxicidad de manganese en plantas de plátano (Musa AAB, cv. “Falso Cuerno”) cultivadas en domos de reciente construcción. CORBANA 27(54):133-144. [ Links ]
VARGAS A., SOLIS P. 1999. Síntomas de deficiencia y contenido de macro y micronutrimentos en las plantas de banana (Musa AAA) bajo condiciones de carencia inducida en cultivo hidróponico. CORBANA 24(51):21-42. [ Links ]
VOLSCHENK C.G., HUNTER J.J., LE ROUX D.J., WATTS J.E. 1999. Effect of graft combination and position of application on assimilation and translocation of zinc in grapevines. Journal of Plant Nutrition 22:115-119. [ Links ]
WEINGERL V., KERIN D. 2000. Distribution of zinc in vineyard areas treated with zinc containing phytopharmaceuticals. Acta Chimica Slovenica 47(4):453-468. [ Links ]
ANDREW C., HEGARTY M. 1969. Comparative responses to manganese excess of eight tropical and four temperate pasture legume species. Australian Journal of Agricultural Research 20(4):687-696. [ Links ]
ARIAS F. 2002. Análisis y comentario: el manganeso (Mn) en la nutrición del cultivo de banano (Musa AAA). CORBANA 28(55):83-96. [ Links ]
ARIAS F., MATA R., ALVARADO A., SERRANO E., LAGUNA J. 2010. Caracterización química y clasificación taxonómica de algunos suelos cultivados con banano en las llanuras aluviales del Caribe de Costa Rica. Agronomía Costarricense 34(2):175-195. [ Links ]
ARIAS F., SERRANO E., GUZMAN M. 2006. Absorción y translocación de Zinc asperjado al follaje de plantas jóvenes de banano (Musa AAA. CV. Grande Naine), desarrolladas en cultivo hidropónico. CORBANA 32(59):79-85. [ Links ]
BUKOVAC M., WITTWER S. 1957. Absorption and mobility of foliar applied nutrients. Plant Physiology 32(5):428-435. [ Links ]
DECKERS T., DAEMEN E., LEMMENS K., MISSOTTEN C. 1997. Influence of foliar applications of Mn during summer on the fruit quality of Jonagold. Acta Horticulturae 448:467-474. [ Links ]
DÍAZ R., HUNTER A. 1978. Metodologías de muestreo de suelos, análisis químico de suelos y tejido vegetal y de investigación en invernadero. CATIE. Turrialba, Costa Rica, 62 p. [ Links ]
ELAMIN O., WILCOX G. 1986a. Effect of magnesium and manganese nutrition on muskmelon growth and manganese toxicity. Journal of the American Society for Horticultural Science 111(4):582-587. [ Links ]
ELAMIN O., WILCOX G. 1986b. Effect of magnesium and manganese nutrition on watermelon growth and manganese toxicity. Journal of the American Society for Horticultural Science 111(4):588-593. [ Links ]
FAGERIA N.K., BARBOSA M.P., MOREIRA A., GUIMARAES M. 2009. Foliar fertilization of crops plants. Journal of Plant Nutrition 32:1044-1064. [ Links ]
FERNÁNDEZ V., EICHERT T. 2009. Uptake of hydrophilic solutes through plant leaves: current state of knowledge and perspectives of foliar fertilization. Critical Reviews in Plant Sciences 28(1):36-68. [ Links ]
FONTES P.C.R., MOREIRA M.A., FONTES R.L.F., CARDOSO A.A. 1999. Effects of zinc fungicides and different zinc fertilizer application methods on soluble and total zinc in potato plant shoots. Communications in Soil Science and Plant Analysis 30(13-14):1847-1859. [ Links ]
GUNES A., ALPASLAN M., INAL A. 1998. Critical nutrient concentrations and antagonistic and synergistic relationships among the nutrients of NFT-grown young tomato plants. Journal of plant nutrition 21(10):2035-2047. [ Links ]
HASLETT B.S., REID R.J., RENGEL Z. 2001. Zinc mobility in wheat: uptake and distribution of zinc applied to leaves or roots. Annals of Botany 87(3):379-386. [ Links ]
HEENAN D., CAMPBELL L. 1981. Influence of potassium and manganese on growth and uptake of magnesium by soybeans ( (L.) Merr. Cv. Bragg). Plant and Soil 61(3):447-456. [ Links ]
HENRÍQUEZ C., BERTSCH F., SALAS R. 1995. Fertilidad de suelos: Manual de laboratorio. ACCS. San José, Costa Rica. 64 p. [ Links ]
HOCKING P.J. 1980. The composition of phloem exudate and xylem sap from tree tobacco ( Grah.). Annals of Botany 45:633-643. [ Links ]
HUMPHRIES J.M., STANGOULIS J.C.R., GRAHAM R.D. 2007. Manganese, pp. 351-374. In: A. Barker y D. Pilbeam (eds.). Handbook of Plant Nutrition. CRC Press. [ Links ]
KAYA C., BURTON M., HIGGS D. 2001. Responses of tomato cvs., grown to fruit–harvest stage under zinc stress in glasshouse conditions. Journal of Plant Nutrition 24(2):369-382. [ Links ]
KOCHIAN L.V. 1991. Mechanisms of micronutrient uptake and translocation in plants, pp. 229-296. In: J.J. Mortvedt, F.R. Cox, L.M. Shuman and R.M. Welch (eds.). Micronutrients in Agriculture, ed. 2. Soil Science Society of America. [ Links ]
KOWALENKO C., IHNAT M. 2010. Effects of soil limestone and foliar manganese and zinc applications on field cauliflower growth and element contents. Canadian Journal of Soil Science 90(1):177-188. [ Links ]
LE BOT J., GOSS M., CARVALHO M.J., BEUSICHEM M.L., KIRKBY E. 1990. The significance of the magnesium to manganese ratio in plant tissues for growth and alleviation of manganese toxicity in tomato (Lycopersicon esculentum) and wheat (Triticum aestivum) plants. Plant and Soil 124(2):205-210. [ Links ]
LINDSAY W. 1972. Zinc in soils and plant nutrition. Advances in Agronomy 24:147-186. [ Links ]
LO C.C., HO M.H., HUNG M.D. 1996. Use of high-performance liquid chromatographic and atomic absorption methods to distinguish Propineb, Zineb, Maneb, and Mancozeb fungicides. Journal of Agricultural and Food Chemistry 44(9):2720-2723. [ Links ]
LOPEZ M., SOLIS P. 1991. Contenidos e interacciones de los nutrimentos en tres zonas bananeras de Costa Rica. CORBANA 15(36):25-32. [ Links ]
MARSCHNER H. 1995. Mineral nutrition of higher plants. Academic Press. Cambridge, UK. 889 p. [ Links ]
MÉNDEZ J.C., BERTSCH F. 2012. Guía para la interpretación de la fertilidad de los suelos de Costa Rica. ACCS, San José, Costa Rica. 108 p. [ Links ]
MERHAUT D. 2007. Magnesium, pp. 146-181. In: A. Barker y D. Pilbeam (eds.). Handbook of Plant Nutrition. CRC Press. [ Links ]
MILLS H.A., BONES J.B. 1996. Plant Analysis Handbook II. Athens, GA: MicroMacro Publishing, Inc. 422 p. [ Links ]
MOREIRA R.S. 1999. Banana: Teoria e practica de cultivo. 2 ed. Fundaçao Cargill, São Pablo, Brasil. 335 p. [ Links ]
POH B.L., GEVENS A., SIMONNE E., SNODGRASS C. 2011. Estimating copper, manganese and zinc micronutrients in fungicide applications. UF/IFAS, University of Florida. Consultado el 24-07-2012. Disponible en http://edis.ifas.ufl.edu/hs1159. [ Links ]
QUARTIN V.M.L., ANTUNES M.L., MURALHA M.C., SOUSA M.M., NUNES M.A. 2001. Mineral imbalance due to manganese excess in triticales. Journal of plant nutrition 24(1):175-189. [ Links ]
SOTO M. 2008. Bananos. Técnicas de Producción, Manejo Postcosecha y Comercialización. Litografía e Imprenta LIL. San José, Costa Rica. 1090 p. [ Links ]
STOCK D., HOLLOWAY P.J. 1993. Possible mechanisms for surfactant-induced foliar uptake of agrochemicals. Pesticide Science 38:165-177. [ Links ]
TIANCANG Z., HUA Z., LILI H., HUANMIN X., DONGZI Z., JINGJING C. 2008. Efficacy of propineb for controlling leaf blotch caused by Marssonina coronaria and its effect on zinc content in apple leaves. Acta Phytophylacica Sinica 35(6):519-524. [ Links ]
TURNER D.W., BARKUS B. 1980. Plant growth and dry-matter production of the ‘Williams’ banana in relation to supply of potassium, magnesium and manganese in sand culture. Scientia Horticulturae 12(1):27-45. [ Links ]
TURNER D.W., BARKUS B. 1983. The uptake and distribution of mineral nutrients in the banana in response to supply of K, Mg and Mn. Fertilizer Research 4:89-99. [ Links ]
VARGAS A. 2001. Descripción y observaciones de la toxicidad de manganese en plantas de plátano (Musa AAB, cv. “Falso Cuerno”) cultivadas en domos de reciente construcción. CORBANA 27(54):133-144. [ Links ]
VARGAS A., SOLIS P. 1999. Síntomas de deficiencia y contenido de macro y micronutrimentos en las plantas de banana (Musa AAA) bajo condiciones de carencia inducida en cultivo hidróponico. CORBANA 24(51):21-42. [ Links ]
VOLSCHENK C.G., HUNTER J.J., LE ROUX D.J., WATTS J.E. 1999. Effect of graft combination and position of application on assimilation and translocation of zinc in grapevines. Journal of Plant Nutrition 22:115-119. [ Links ]
WEINGERL V., KERIN D. 2000. Distribution of zinc in vineyard areas treated with zinc containing phytopharmaceuticals. Acta Chimica Slovenica 47(4):453-468. [ Links ]
*Correspondencia a:
Juan Carlos Méndez. Centro de Investigaciones Agronómicas, Universidad de Costa Rica. San José, Costa Rica. Autor para correspondencia. Correo electrónico: juancarlos.mendez@ucr.ac.cr
Floria Bertsch. Centro de Investigaciones Agronómicas, Universidad de Costa Rica. San José, Costa Rica.
Oscar Castro.Laboratorio de Fitopatología, Universidad de Costa Rica. San José, Costa Rica.
1. Autor para correspondencia. Correo electrónico: juancarlos.mendez@ucr.ac.cr
Oscar Castro.Laboratorio de Fitopatología, Universidad de Costa Rica. San José, Costa Rica.
1. Autor para correspondencia. Correo electrónico: juancarlos.mendez@ucr.ac.cr
* Centro de Investigaciones Agronómicas, Universidad de Costa Rica. San José, Costa Rica.
** Laboratorio de Fitopatología, Universidad de Costa Rica. San José, Costa Rica.
** Laboratorio de Fitopatología, Universidad de Costa Rica. San José, Costa Rica.
Recibido: 06/08/12 Aceptado: 20/11/12


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}