











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Las especies del género Atherinella, pejerreyes o “sardinas” conforman un grupo cuya monofilia aún debe revisarse (sensuCampanella et al., 2015), pertenecen a la Familia Atherinopsidae sensuSaeed et al., 1994 con una distribución biogeográfica restricta a América del Norte, Central y Sur. Son habitantes de las aguas marinas del Pacífico, Atlántico y Caribe, así como de ambientes de aguas dulces y salobres. En Costa Rica el género Atherinella cuenta en la actualidad con 35 especies, la mayoría son marinos, pero también registra 6 especies de agua dulce que habitan los ríos del país desde los 0 a los 540 m.s.n.m. (Bussing, 2015, Angulo et al., 2013).
A continuación, se describe el primer registro fósil de un ejemplar del género Atherinella, procedente de la localidad de Las Juntas de Abangares, en la provincia de Guanacaste, el cual fue colectado por Héctor Flores Albertazzi (Fig. 1).
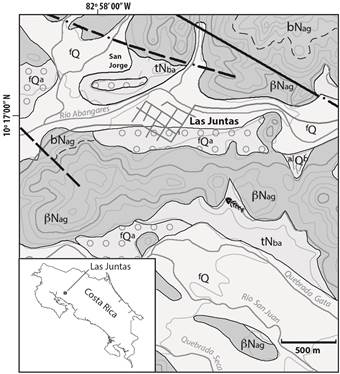
Fig. 1: Mapa geológico y de ubicación del afloramiento de la localidad fosilífera en la margen norte de la Quebrada Gata en las Juntas de Abangares. Simbología: βNag basaltos, andesitas basálticas del Mioceno Superior-Plioceno y bNag brechas andesíticas del Plioceno Superior del Grupo Aguacate; tNba unidad de tobitas con fósiles del Plioceno Inferior de la Formación Bagaces; fQa paleoterrazas fluviales con arenas y gravas laterizadas del Pleistoceno; fQ depósitos fluviales y alQb abanicos aluviales del Holoceno. Redibujado y modificado de Žáček et al., 2010.
Geología
La Formación Bagaces en la hoja cartográfica Juntas de acuerdo a Žáček et al., 2010 y Kycl et al., 2010 está formada por intercalaciones de ignimbritas ácidas y tobas que a nivel local contienen fósiles de origen marino, estas capas basculan en general 30° al NE, sin embargo, la inclinación debido al diastrofismo cambia con frecuencia.
Las secuencias tobáceas de la Formación Bagaces se pueden observar en las canteras próximas al pueblo de Palmita, al NW del Río Cañamazo y en el SW de la ladera del cerro Lomas Nancital y se extienden hacia el norte de la ciudad de Las Juntas, aflorando también a largo del valle del río San Juan.
En la sección inferior del afloramiento de la Palmita abundan las frondas carbonizadas, braquiópodos del género Glottidia y en la sección superior se registra gran cantidad de fauna bivalvos como Argopecten sp., Pinna sp. y Tellina sp., y restos de cangrejos.
Por su parte, la litofacies, así como los braquiópodos y moluscos presentes en dichas secuencias, sugieren ambientes marinos sublitorales y permiten correlacionar estos afloramientos con la unidad sedimentaria Tobitas Lomas Barbudal (sensuAguilar y Alvarado, 2004) de la Formación Bagaces.
La unidad de Tobitas (sensuŽáček et al., 2010) aflorantes al SSE en la ciudad de Las Juntas, lugar de donde proviene el ejemplar del presente estudio, se inter-estratifica con la secuencia ignimbrítica característica de la Formación Bagaces y presenta estratos decimétricos con laminación planar milimétrica, conformando una unidad sedimentaria de piroclastitas subacuáticas con restos de frondas y peces.
Paleontología sistemática
Subdivisión Teleostei Müller Serie Atherinomorpha Greenwood et al.
Orden Atheriniformes Rosen
Suborden Atherinopsoidei Campanella et al.
Familia Atherinopsidae Fowler
Subfamilia Menidiinae Schultz
Tribu Membradini Chernoff
Género Atherinella Steindachner Atherinella sp. indet.
Material: Un ejemplar casi completo, molde y contra-molde depositados en la Colección de fósiles de la Sección de Geología, Departamento de Historia Natural del Museo Nacional de Costa Rica, bajo el código CFM-5290. Recuperado en las coordenadas 10°16’28” N y 84°57’17” W, hoja 3246 IV Juntas por Flores Albertazzi.
Descripción: El fósil con una longitud total de 49 mm, está constituido por un molde y un contramolde en el que no se preservaron elementos óseos, solamente sus improntas, La región craneal tiene 9 mm de diámetro y la aleta caudal 14 mm de largo (Figuras 2, 3 y 5).

Fig. 2: Atherinella sp. indet., A: vista superior corresponde con el molde del lado izquierdo y B: vista inferior corresponde con el contra molde del lado derecho, la longitud del fósil es de 49 mm.
En la región craneal se observan las oquedades de las orbitas oculares, notablemente grandes que alcanzan los 3 mm de longitud. De igual manera se distingue la región donde se ubicaba el opercular con una longitud de 5 mm.
Se preservaron las 2 aletas pectorales, 2 aletas ventrales, la aleta anal y la aleta caudal. No se observa la impronta de ninguna de las aletas dorsales.
El arco pectoral derecho se encuentra dislocado y está representado por las improntas del cleitro con una longitud total de 6 mm y la aleta pectoral con 5 mm de ancho, constituida por 6 largos radios visibles y en el extremo izquierdo 7 radios distales cortos y delgados, no se distinguen improntas del coracoides, radiales u otras estructuras de la aleta. El arco pectoral izquierdo también se encuentra dislocado y se ubica en el costado derecho por delante de la impronta de la aleta anal, donde se observa la aleta pectoral constituida por 11 radios y la impronta del cleitro de manera superpuesta (Fig. 3).
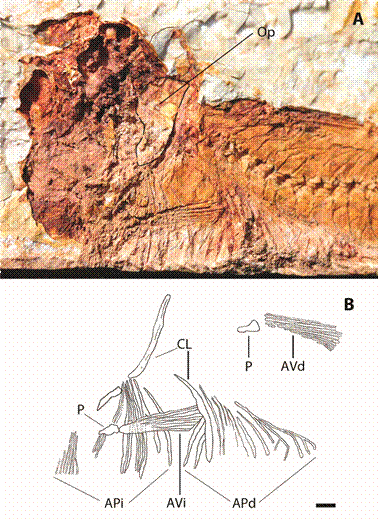
Fig. 3: Atherinella sp. indet.: A. detalle de mitad anterior del espécimen, en la región del cráneo se detallan: OP probable región del opercular y B. esquema de las extremidades, CL cleitro, AP aletas pectorales, P pelvis, AV aletas ventrales, la i y la d indican la correspondiente lateralidad izquierda o derecha. Escala visual 1 mm.
Se preservaron las improntas de ambas aletas pélvicas, donde se definen los radios de las respectivas aletas, la izquierda unida a la pelvis correspondiente y la derecha separada de su pelvis. La longitud de la impronta de la pelvis derecha alcanza los 3,5 mm y su aleta 6 mm, la impronta de la aleta pélvica izquierda alcanza los 5,5 mm y su pelvis 3,4 mm.
La aleta anal está bien preservada y tiene una longitud de 12 mm, ésta se muestra plegada con un total de 24 radios observables, la mitad anterior presenta los primeros 10 radios relativamente largos con longitudes variables entre los 4,0 y 2,6 mm, mientras la porción posterior presenta 14 radios con longitudes de entre 1,5 y 0,5 mm, ver Fig. 4.
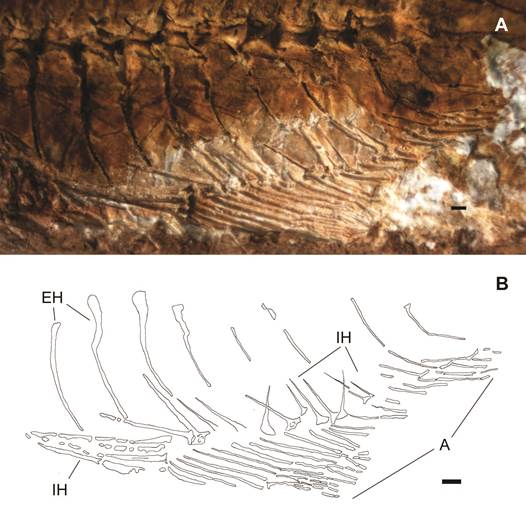
Fig. 4: Atherinella sp. indet., Aleta anal: A. detalle y B. esquema osteológico, EH espinas hemales, IH radios interhemales, AA aleta anal. Escala gráfica 0,5 mm.
Los primeros 3 radios inter-hemales (IH) son notablemente gruesos con longitudes de 4 mm, las improntas de los restantes 12 radios inter-hemales, son aciculares ensanchados en sus extremos distales, siendo los últimos conspicuamente delgados y cortos.
Discusión y conclusiones
El ejemplar del presente estudio posee una buena articulación de la mayor parte del esqueleto, la cabeza está dislocada y se observa en posición dorsal, desplazada hacia la parte media del cuerpo, ubicándose por sobre la aleta pélvica izquierda. De igual manera el arco pectoral izquierdo que incluye el cleitro probablemente junto con el supracleitro y la aleta pectoral está desplazado hacia atrás y abajo, superponiéndose la aleta pectoral sobre la aleta pélvica izquierda; la aleta pectoral derecha se dislocó completamente, llegándose a ubicar en el costado izquierdo del cuerpo justo entre las aletas anal y la aleta pélvica. La aleta pélvica derecha está dislocada y desplazada hacia la porción dorsal y hacia atrás, ligeramente adelantada con respecto a la aleta anal. En lo que respecta a la mayor parte del cuerpo desde la región post-craneal hasta la aleta caudal se encuentra de perfil o en norma lateral.
La forma en que se preservó el cuerpo supone muy poco o nada de transporte post-mortem, al parecer el individuo se hundió y se depositó en un fango donde se preservó, en el proceso la cabeza se disloca, pero no llegó a desprenderse del todo, al igual que las aletas pectorales que se desplazaron un poco sin separarse totalmente del cuerpo.
Lo que si llama la atención es la no preservación ósea, no se observa en ningún caso un resto óseo, no se encontraron dientes ni otolitos, solo moldes, probablemente debido a la acides del agua, el ambiente de oxidación y el sedimento mismo.
La determinación a nivel genérico se fundamenta en las siguientes características: la aleta anal presenta un número de radios que es acorde con el descrito para el género Atherinella; la longitud de los radios anteriores es notablemente más larga que los posteriores lo que evidencia la forma triangular alargada propia del género. La forma bifurcada de la aleta anal (Fig. 5B), el hecho de que las aletas pélvicas son relativamente cortas, la distribución topológica de todo el conjunto de aletas, además de las enormes cavidades orbitales (fig. 5 A), así como las dimensiones observadas del cuerpo, permite concluir que se trata del género Atherinella. La probable longitud del individuo en vida varió entre los 65 a 70 mm.

Fig. 5: Atherinella sp. indet., A. detalle de la región craneal del contra molde donde se observa las Or orbitas oculares y B. detalle de la AC aleta caudal, también se observa la AA aleta anal.
De momento con el material disponible y a falta de más ejemplares con elementos característicos y una mejor preservación, no es posible realizar una determinación a nivel específico del ejemplar, aunque el número de radios en la aleta anal que es de 24 se encuentra en el rango observado para dos de las especies actuales Atherinella milleri con número variable entre 23 a 27 y Atherinella sardina con numero variable entre 19 y 25 (cf. Bussing, 2015).
Paleoecología y paleogeografía
Los lechos de tobitas heterolíticas donde se halló el pez, conforman una unidad sedimentaria de piroclastitas subacuáticas asociada a facies de estuario. La naturaleza estuarina de estas secuencias de tobitas, se evidencia en secuencias volcano-sedimentarias similares de la Formación Bagaces, aflorantes por ejemplo, en las Lomas de Barbudal a aproximadamente 72 km al NW del sitio del hallazgo, dónde Aguilar y Alvarado (2004) describen el registro fósil parautóctono de moluscos marinos y braquiópodos del género Glottidia, sugiriendo un ambiente estuarino influenciado por las mareas, y en los sedimentos aflorantes a 5 kilómetros al sur de la ciudad de Las Juntas, a lo largo del pueblo de Palmita, al NW del Río Cañamazo y en al SW de la ladera del cerro Lomas Nancital donde se registra una asociación de invertebrados marinos y el braquiópodo Glottidia sp. (Žáček et al., 2010 y Kycl et al., 2010).

Fig. 6: Atherinella sp. indet., reconstrucción basada en especímenes actuales del género. Dibujo realizado por Daniel Acosta Mora.
La diferencia paleoambiental más notoria entre las secuencias de Lomas Barbudal y las aflorantes al sur de la hoja Juntas con las tobitas de la localidad donde se recuperó el pez fósil o las aflorantes al este del Barrio San Jorge radica principalmente en el no registro de invertebrados marinos, lo que se puede interpretar como un ambiente con muy baja salinidad o predominantemente dulceacuícola, quizás un ambiente lagunar asociado a un antiguo estuario. Además, los sedimentos constituidos por piroclastitas subacuáticas se caracterizan por una fina laminación heterolítica donde se alternan niveles milimétricos a sub milimétricos de cenizas de color blanco, arcillas grises y limos muy finos férricos, lo que sugiere un ambiente de oxidación, con poca corriente, probablemente lagunar salobre a dulceacuícola.
El escenario paleogeográfico general sugiere una ingresión marina ya sea por elevación del nivel del mar o por subsidencia de la cuenca, tiempo en el cual se favoreció el establecimiento de un estuario afectado por una notable sedimentación piroclástica y el desarrollo de ambientes de aguas salobres y dulces tierra adentro. Luego, la posterior retrogradación del mar dio paso de nuevo a la deposición de piroclastos en ambiente sub-aéreos, lo que es concordante con el modelo expuesto por Aguilar y Alvarado (2004).
La importancia de un evento paleogeográfico de este tipo, donde primero se dio una ingresión marina y luego una caída del nivel del mar, esto último probablemente por un levantamiento del arco de islas, coadyuva a comprender los procesos de especiación y colonización del género Atherinella, inicialmente habitante de ambientes marinos y salobres que luego se adaptó a ambientes eminentemente dulceacuícolas, colonizando incluso, localidades bastante alejadas de las costas, tierra adentro.
Esta plasticidad ambiental del género Atherinella puede ser perfectamente explicado en razón de los procesos tectónicos que dieron origen a una especiación vicariante, algo similar a lo observado en especies dulce acuícolas actuales que divergieron fenotípicamente en razón de unas pocas decenas de años de sus ancestros oceánicos luego de un levantamiento tectónico importante (Lescak et al., 2015).
Edad
Teniendo en cuenta la fauna de invertebrados asociada a las secuencias de tobas y tobitas de Las Juntas, es posible correlacionar dichas secuencias con las de la unidad sedimentaria Tobitas Lomas Barbudal (sensu Aguilar y Alvarado, 2004 y Denyer et al., 2013) quienes sugieren una edad Mioceno Superior - Plioceno Inferior. Por correlación estratigráfica Žáček et al., 2010 y Kycl et al., 2010 les asignan a las tobas y tobitas del área de Las Juntas una edad Plioceno Inferior.
Distribución paleobiogeográfica
A pesar de que los tiempos de divergencia obtenidos a partir de las hipótesis filogenéticas con base en análisis de ADN, sugieren que el género Atherinella surgió en el Oligoceno Superior (sensu Campanella et al., 2015) y de la amplia distribución actual del género a lo largo de todo el continente, el hallazgo acá descrito, corresponde con el primer registro fósil conocido para el género Atherinella.

