











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los equinodermos constituyen uno de los grupos animales más conspicuos de los océanos (Lawrence, 1987) y generalmente son dominantes en términos de abundancia y diversidad en las comunidades bentónicas, donde juegan un rol ecológico de gran relevancia al ocupar diversos niveles de las redes tróficas y actuar como depredadores tope (Blankley, 1984; Reyes-Bonilla et al., 2005). En particular, en aguas antárticas y en el Océano Atlántico, los asteroideos se encuentran entre los grupos taxonómicos bentónicos más abundantes (Fraysse et al., 2018, Mah & Blake, 2012; Moreau, 2019).
Las estrellas de mar presentan diferentes patrones de desarrollo que incluyen desde desarrollo indirecto a directo. El estadio larval puede ser pelágico o bentónico, con nutrición planctotrófica o lecitotrófica (Chia, 1974; McEdward & Miner, 2001; Mileikovsky, 1971). La mayoría de los asteroideos de diversos hábitats poseen larvas pelágicas planctotróficas o lecitotróficas que se dispersan ampliamente (Chia & Walker, 1991; Hyman, 1955). Sin embargo, un número más reducido de especies poseen larvas lecitotróficas no pelágicas (bentónicas) incubadas por los progenitores (larva retenida), e incluso el estadio de larva puede encontrarse ausente y el embrión se desarrolla de forma directa.
Estas estrategias han sido ampliamente discutidas, especialmente respecto a la importancia de la incubación y el cuidado parental (Gillespie & McClintock, 2007). Por ejemplo, Leptasterias polaris (Müller & Troschel, 1842) según Hamel y Mercier (1995) o Himmelman et al. (1982) y Asterina phylactica Emson & Crump, 1979 según Strathmann et al. (1984) protegen a sus embriones debajo de su cuerpo. Por otro lado, Leptasterias hexactis (Stimpson, 1862) según Chia (1968), Leptasterias ochotensis (Brandt, 1851) según Kubo (1951), Anasterias antarctica (Lütken, 1857) en Fraysse et al., (2020) o en Pérez et al. (2015) y Anasterias minuta Perrier, 1875 en Gil y Zaixso (2007) o en Salvat (1985) mencionan que preparan una cámara de incubación arqueando sus brazos. En Ctenodiscus australis Loven in Lütken, 1871 y en el género Leptychaster (Lieberkind, 1926; Rivadeneira et al., 2017) se observó que la incubación se realizaba entre las espinas de la cara aboral mientras que en especies de la familia Pterasteridae la incubación se lleva a cabo en la cámara nidamental aboral (Fraysse et al., 2020; Hyman, 1955; McClary & Mladenov, 1989). Dentro de las especies incubadoras internas se incluyen a Leptasterias groenlandica (Steenstrup, 1857), que mantiene sus embriones en el estómago (Lieberkind, 1920) y un número de especies del género Patiriella que incuban sus crías en el espacio intragonadal (Byrne, 1996; Byrne, 2005; Dartnall et al., 2003; Komatsu et al., 1990).
El interés científico por los asteroideos antárticos, particularmente por sus patrones de desarrollo se remonta a fines del siglo XIX tras la expedición pionera del R/V Challenger (Thomson, 1876; Thomson, 1885), donde se registró una elevada incidencia de especies que presentan desarrollo no pelágico. Posteriormente, las observaciones realizadas en equinodermos y otros invertebrados bentónicos llevaron a fundar la idea de que el desarrollo no pelágico era el modo de desarrollo dominante entre de los animales marinos bentónicos de aguas frías, como estrategia adaptativa a las condiciones de los ambientes Ártico, Antártico y región subantártica (Murray, 1895), conduciendo a la formulación de la “Regla de Thorson” (Mileikovsky, 1971). Sin embargo, con la ampliación de los casos de estudio esta regla fue perdiendo sustento (Clarke, 1992; Pearse et al., 1991; Pearse et al., 2009; Pearse & Bosch, 1994) y en la actualidad es ampliamente aceptada la noción de que la mayor incidencia de especies con desarrollo no pelágico en aguas antárticas no es consecuencia de una simple adaptación a las condiciones polares (Dell, 1972) sino que se corresponde con la historia evolutiva de cada grupo, siendo consecuencia de la presión selectiva generada por el aislamiento luego de eventos vicariantes que condujeron a la proliferación específica (Pearse et al., 2009).
El objetivo del presente trabajo fue revisar la ocurrencia, distribución batimétrica, composición específica y riqueza de especies de asteroideos que habitan en los archipiélagos Orcadas del Sur, Shetland del Sur y Archipiélago de Palmer, ubicados en la Península Antártica. A su vez, detallar aspectos de los patrones de desarrollo de las especies halladas.
Materiales y métodos
Sitio de muestreo: El presente estudio se llevó a cabo durante las Campañas Antárticas de Verano (CAV) 2012 y 2013, a bordo del Buque Oceanográfico “Puerto Deseado” (Fig. 1). La zona de estudio comprendió la Península Antártica, entre 60° 27’ 36’’ S & 44° 23’ 60’’ W y 65° 22’ 31’’ S & 66° 22’ 55’’ W. Se realizaron en total 28 lances de pesca, mediante la utilización de una red piloto, en tres archipiélagos: Orcadas del Sur (OS), Shetland del Sur (SS) y Archipiélago de Palmer (AP) (Fig. 2). Los muestreos de los individuos fueron cualitativos. Además, en cada estación se registraron los parámetros temperatura, salinidad y profundidad (Tabla 1) mediante una sonda oceanográfica CTD (conductividad eléctrica, temperatura y la presión del agua de mar).

Fig. 1 Fotografías A) del Buque Oceanográfico “Puerto Deseado” (Foto: Manuel Soria) y B) de la zona de muestreo en la Península Antártica (Foto: Mariel Ojeda). / Fig. 1. Photographs of A. the oceanographic vessel ARA “Puerto Deseado” (Photo: Manuel Soria) and the B. sampling area of Antarctic Peninsula (Photo: Mariel Ojeda).
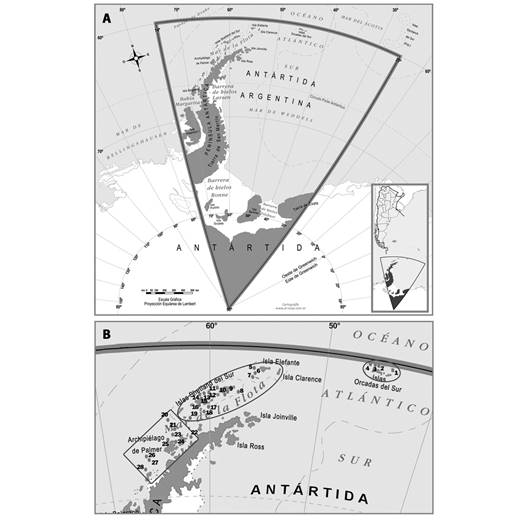
Fig. 2 Mapa de la Península Antártica, el recuadro negro indica el área de estudio. A. Sector Antártico Argentino. B. En los tres archipiélagos analizados se indican las estaciones de muestreo con puntos negros: Orcadas del Sur (1-4), Shetland del Sur (5-19) y Archipiélago de Palmer (20-28). / Fig. 2. Map of the Antarctic Peninsula, the black box indicates the study area A. Argentine Antarctic Sector. B. In the three archipelagos analysed, sampling stations are indicated with black dots: South Orkney (1-4), South Shetland (5-19) and Palmer Archipelago (20-28).
Tabla 1 Parámetros físicos del agua de mar (temperatura y salinidad) y profundidad máxima de los sitios de muestreo en Orcadas del Sur (OS), Shetland del Sur (SS) y Archipiélago de Palmer (AP), ubicados en la Península Antártica. / Table 1. Physical seawater parameters (temperature and salinity) and maximum depth of the sampling sites in the South Orkney (SO), South Shetland (SS) and Palmer Archipelago (PA), located on the Antarctic Peninsula.
| Archipiélago | Profundidad (m) | Temperatura (°C) | Salinidad (‰) | ||||
| Min | Max | Min | Max | Min | Max | ||
| Orcadas del Sur | 80.0 | 120.0 | 1.50 | 3.70 | 34.08 | 34.14 | |
| Shetland del Sur | 60.2 | 977.0 | 1.57 | 2.42 | 33.90 | 34.15 | |
| Archipiélago de Palmer | 170.0 | 562.0 | 1.56 | 3.13 | 32.90 | 33.98 | |
Tratamiento de las muestras: Cada estrella de mar recolectada fue fotografiada y disecada. Se midió el radio mayor (mm), se registraron las características macroscópicas de las gónadas de cada individuo, y se examinó la presencia de embriones.
Taxonomía, distribución batimétrica y área de distribución geográfica: Las identificaciones taxonómicas fueron realizadas en base a Bernasconi (1962), Bernasconi (1970), Clark y Downey (1992), Fisher (1940), Koehler (1912), Mah (2020), Mah et al. (2014), Sladen (1889), Tablado (1982), y Verril (1914) y posteriormente controlados con Registro de Especies Marinas Antárticas (Clarke & Johnston, 2003; De Broyer et al., 2022), y Global Biodiversity Information Facility (GBIF) (2022) para garantizar la eliminación de sinonimias o errores ortográficos, así como para comparar el área de distribución y ocurrencia conocida con la registrada en este estudio.
Riqueza de especies, comparación entre archipiélagos: Debido a los desiguales esfuerzos de muestreo y al uso de artes heterogéneas para recolectar los individuos, elegimos datos binarios (presencia/ausencia) para construir la matriz de similitud de las especies. Se utilizó el índice de Sörensen para construir la matriz de datos (Clarke et al., 2006). Se realizó una agrupación jerárquica para analizar las similitudes en la composición de las especies entre las diferentes áreas; se obtuvo mediante la técnica de agrupación por enlace de grupos y se representó gráficamente en un dendrograma, utilizando el software Primer V6.0 (Anderson et al., 2008). Se eligió un valor del 60 % de similitud como umbral para agrupar las regiones.
Patrones de desarrollo: El patrón de desarrollo de cada especie recolectada fue revisado basándonos en antecedentes bibliográficos (Bernasconi, 1970; Hyman, 1955; Janies, 1995; McClary & Mladenov, 1990; Pearse & Bosch, 1994).
Resultados
Parámetros ambientales: Los parámetros físicos del agua de mar (temperatura y salinidad) y la profundidad máxima y mínima de las estaciones de muestreo localizadas en los tres archipiélagos antárticos se detallan en la Tabla 1. Se observa que tanto la temperatura como la salinidad fueron similares entre los tres archipiélagos.
Taxonomía, distribución batimétrica y área de distribución geográfica: Se registraron un total de 436 asteroideos, distribuidos en 28 estaciones de muestreo (100 % de ocurrencia) ubicadas en tres archipiélagos. Se determinaron 21 especies de estrellas de mar pertenecientes a nueve familias y 16 géneros (Tabla 2). Por otro lado, algunos individuos se identificaron a nivel de género (Tabla 2) mientras que seis morfotipos no pudieron ser determinados.
Las especies identificadas fueron Odontaster validus Koehler, 1906 (n = 44), Bathybiaster loripesSladen, 1889 (n = 39), Odontaster meridionalis (E. A. Smith, 1876) (n = 32), Glabraster antarctica (E. A. Smith, 1876) (n = 30), Psilaster charcoti (Koehler, 1906) (n = 29), Solaster regularis subarcuatus Sladen, 1889 (n = 27), Cheiraster gerlachei Ludwig, 1903 (n = 21), Pteraster stellifer Sladen, 1882 (n = 18), Cycethra verrucosa mawsoni A.M. Clark, 1962 (n = 17), Cryptasterias turqueti (Koehler, 1906) (n = 14), Lysasterias perrieri (Studer, 1885) (n = 10), Bathybiaster loripes obesus Sladen, 1889 (n = 7), Cuenotaster involutus (Koehler, 1912) (n = 7), Pteraster affinis Smith, 1876 (n = 6), Diplasterias brucei (Koehler, 1907) (n = 5), Acodontaster elongatus (Sladen, 1889) (n = 4), Acodontaster conspicuus (Koehler, 1920) (n = 3), Perknaster aurorae (Koehler, 1920) (n = 3), Solaster regularis Sladen, 1889 (n = 1), Lophaster gaini Koehler, 1912 (n = 1) y Macroptychaster accrescens (Koehler, 1920) (n = 1).
La distribución batimétrica de los asteroideos fue de 6 a 987 m de profundidad, ajustándose el rango batimétrico de todas las especies encontradas con las previamente reportadas. El área de distribución geográfica conocida para las especies se obtuvo mediante GBIF (2022) para los tres archipiélagos. Las fuentes de información utilizadas fueron: “NMNH Extant Specimen Records (USNM, US); MARINe/PISCO: Intertidal: MARINe Long-Term Monitoring Surveys: Sea Stars and Katharina; The echinoderm collection (IE) of the Muséum national d’Histoire naturelle (MNHN - Paris); NOAA AFSC North Pacific Groundfish Observer; Environmental Monitoring database (MOD) DNV; Marine Nature Conservation Review (MNCR). Se extendió el área de distribución geográfica de seis especies: Cryptasterias turqueti en Orcadas del Sur y Shetland del Sur, Cycethra verrucosa mawsoni y Perknaster aurorae en Shetland del Sur, y Pteraster stellifer y Solaster regularis subarcuatus en el Archipiélago de Palmer.
Riqueza de especies, comparación entre archipiélagos: En relación a la riqueza de especies, los Archipiélagos de Palmer y Shetland del Sur presentaron la máxima riqueza, con 12 especies en cada sitio, mientras que en Orcadas del Sur se registró la menor riqueza, con ocho especies (Tabla 2).
El análisis de conglomerados sugirió dos grupos regionales con una composición faunística similar (Fig. 3). Uno de los clústeres estuvo compuesto por Shetland del Sur y Orcadas del Sur con una similitud de 64 %, y el segundo grupo correspondió al Archipiélago de Palmer.
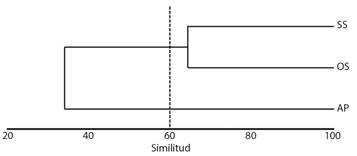
Fig. 3 Agrupación jerárquica (promedio de grupos) de la fauna de asteroideos. Se eligió un valor del 60 % de similitud (distancia de Sörensen) como umbral para agrupar las regiones. Referencias: Shetland del Sur (SS), Orcadas del Sur (OS) y Archipiélago de Palmer (AP). / Fig. 3. Hierarchical clustering (group average) of the Asteroidea fauna. A value of 60 % of similarity (Sörensen distance) was chosen as a threshold to group regions. References: South Shetland (SS), South Orkney (OS) and Palmer Archipelago (PA).
Patrones de desarrollo: En cuanto al estudio de los patrones de desarrollo, el 28 % de las especies encontradas en la Península Antártica presentaron cuidado parental (17 % con larva retenida y 11 % con desarrollo directo) mientras que la mayoría presentaron desarrollo pelágico con valores de 55 % y 17 % para las que poseen larva pelágica lecitotrófica y larva pelágica planctotrófica, respectivamente (Tabla 2, Fig. 3). En los tres archipiélagos el patrón de desarrollo mayoritario fue el pelágico con nutrición lecitotrófica. El mayor porcentaje de especies con cuidado parental mediante retención de las larvas fue hallado en Orcadas del Sur (29 %), mientras que especies con desarrollo directo (Pterasteridae, 17 %) sólo se hallaron en el Archipiélago de Palmer (Tabla 2, Fig. 4).
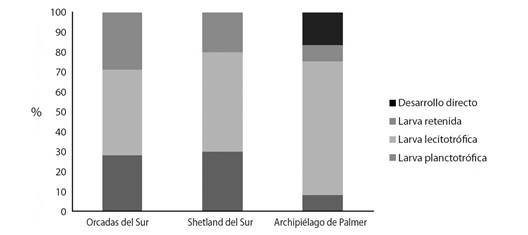
Fig. 4 Frecuencia relativa de especies con diferentes patrones de desarrollo registrados en Orcadas del Sur, Shetland del Sur y Archipiélago de Palmer, ubicados en la Península Antártica. / Fig. 4. Relative frequency of species with different developmental patterns recorded in South Orkney, South Shetland and Palmer Archipelago, located on the Antarctic Peninsula.
Discusión
En el presente trabajo logramos fortalecer la información disponible sobre distribución batimétrica, composición específica, ocurrencia y riqueza de asteroideos hallados en tres archipiélagos Antárticos, incorporando información obtenida durante las Campañas Antárticas de Verano 2012 y 2013 a lo ya descrito para la Península Antártica. Así mismo, discutimos aspectos de los patrones de desarrollo de las especies halladas.
Taxonomía, distribución batimétrica y área de distribución geográfica: Un total de 21 especies de asteroideos pertenecientes a nueve familias fueron recolectadas en las áreas de muestreo. La familia Asteriidae fue la que registró mayor número de especies en Orcadas del Sur, mientras que en Shetland del Sur y Archipiélago de Palmer fueron las familias Odontasteridae y Astropectinidae, respectivamente. Valiéndonos de la base de datos GBIF (2022), se amplió el área de distribución geográfica de seis especies.
Riqueza de especies y comparación entre archipiélagos: La riqueza de especies reportada en nuestro estudio para los Archipiélagos de Palmer y Shetland del Sur fue igual para ambos y mayor que en Orcadas del Sur, siendo en esta última 40 % menor. Los valores de riqueza hallados para los diferentes archipiélagos son consistentes con los resultados arrojados por el análisis de conglomerados, sugiriendo dos grupos regionales con una composición faunística similar, Shetland del Sur y Orcadas del Sur con una similitud de 64 %, y otro grupo representado por el Archipiélago de Palmer. Estos resultados podrían responder a las ecorregiones bénticas que cada uno de los archipiélagos ocupa. El Archipiélago de Palmer y Shetland del Sur se incluyen y comparten la ecorregión bentónica Península Antártica, mientras que Orcadas del Sur pertenecen a la ecorregión bentónica Orcadas del Sur (Douglass et al., 2014). La ecorregión bentónica Península Antártica se caracteriza por presentar una plataforma poco profunda y productiva ubicada al oeste de la Península Antártica con una baja duración de la capa de hielo marino y fondos marinos “cálidos” en comparación con otras zonas de la plataforma antártica e incluye los ecosistemas insulares de las Shetland del Sur, mientras que la ecorregión Orcadas del Sur está representada por los ecosistemas insulares de las islas Orcadas del Sur y los montes submarinos y mesetas del Arco de Scotia, muchos de los cuales subyacen a la Zona Frontal de la Corriente Circumpolar Antártica Meridional (Douglass et al., 2014).
Patrones de desarrollo: Con respecto a las especies con desarrollo no pelágico halladas (larva retenida o desarrollo directo), éstas pertenecen sólo a dos familias. Esta radiación específica podría ser consecuencia del aislamiento tras eventos vicariantes, lo cual concuerda con la hipótesis de Pearse et al. (2009) sobre la selección de este tipo de patrón de desarrollo.
Nuestros resultados indican que los asteroideos con desarrollo pelágico lecitotrófico fueron mayoritarios, seguidos por los de desarrollo no pelágico. Si bien en trabajos de revisión bibliográfica recientes, se ratificó que las especies que presentan cuidado parental son mayoritarias en aguas Antárticas (Moreau, 2019), el bajo porcentaje de especies con desarrollo no pelágico aquí reportadas podría explicarse por el esfuerzo de muestreo, siendo que según Pearse et al., (2009) las especies que presentan larvas pelágicas son más abundantes en aguas antárticas. Sí bien los clados de asteroideos con desarrollo no pelágico son numerosos, la mayoría de las especies son muy poco abundantes (pocas especies muy abundantes con desarrollo pelágico; muchas especies poco abundantes con desarrollo no pelágico). Pearse et al. (1991) y Poulin et al. (2002) sugieren que esta diferencia en los patrones de abundancia se debe a factores ecológicos: las especies con desarrollo pelágico colonizan y prosperan en zonas poco profundas perturbadas por el hielo, mientras que las de desarrollo no pelágico predominan en hábitats más estables y profundos, donde la competencia interespecífica es más intensa.
Conectividad Antártida-subantártida: Es ampliamente difundido que la capacidad de dispersión de larvas y/o embriones, contribuye a explicar sus patrones de distribución geográfica en la región antártica y subantártica (Fraysse et al., 2018; Moreau, 2019). Si bien la mayor capacidad de dispersión se observa en especies con desarrollo pelágico, existe evidencia que especies con desarrollo no pelágico poseen amplia capacidad de dispersión, como es el caso de D. brucei, que puede ser hallada tanto en aguas antárticas como en el Arco de Scotia, Banco Burdwood e Islas Malvinas (Kim & Thurber, 2007). Todas las especies reportadas en el presente trabajo (72 % con desarrollo pelágico y 28 % con desarrollo no pelágico) han sido halladas a ambos lados de la Corriente Circumpolar Antártica (CCA), el 62 % fueron registradas recientemente en la región subantártica por nuestro equipo de trabajo (Fraysse, 2021; Fraysse et al., 2018), mientras que el 38 % restante también presenta registros en dicha región (GBIF, 2022). La Corriente de Malvinas, rama en dirección sur-norte de la CCA, podría facilitar la dispersión de larvas de la región antártica a la región subantártica, y particularmente para las especies no pelágicas se sugieren capacidades inusuales de dispersión, como el “rafting”.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

