











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Los estudios sobre la fenología de plantas son fundamentales para comprender la dinámica de los recursos de las comunidades (Bullock & Solis-Magallanes, 1990), por ejemplo, la actividad de los polinizadores, dispersores de semillas, así como el período de reproducción de muchos animales depende directa o indirectamente de la producción de flores y frutos en una comunidad vegetal (Smith-Ramírez & Armesto, 1994; Terborgh, 1986).
En este sentido y en relación con la actividad apícola, el conocimiento de la riqueza y la distribución temporal de la vegetación en los alrededores de los apiarios es muy importante, ya que provee al apicultor una herramienta básica e indispensable para planificar su actividad (Primost et al., 2013). Esta información permite, por un lado, programar el traslado de las colmenas en función de mayores volúmenes de floración, con la finalidad de optimizar el aprovechamiento de esta o bien para la producción de mieles de determinados orígenes botánicos. Por otro lado, la oferta de floración de un área permite establecer relaciones con los recursos utilizados por la abeja melífera (Apis mellifera L.) e inferir acerca de su comportamiento recolector y preferencias alimenticias (Fagúndez et al., 2016).
En las distintas regiones de Argentina se han desarrollado diversos estudios sobre calendarios apícolas, es así que, para el Chaco Serrano, que pertenece a la ecorregión Chaco Seco (Burkart, et al., 1999), se encuentra lo realizado por Burgos (2012); en el Chaco Húmedo por Cabrera et al. (2013), Salgado (2006) y Salgado et al. (2014). En el Chaco Seco por Salgado (2006) y Salgado et al. (2014); en la Pampa se destacan los realizados por Aramayo et al. (1993), Gurini y Basilio (1995), Tellería (1993) y Tellería (1995); en el Espinal por Andrada (2003) y Naab et al. (2001). En el Monte de Llanuras y Mesetas estos estudios fueron desarrollados por Forcone (2003), Naab y Tamame (2007) y Tamame (2011); en los Bosques Patagónicos los llevados a cabo por Forcone y Kutscher (2006), y en la Estepa Patagónica por Forcone y Muñoz (2009). Para el Dpto. Diamante (Entre Ríos) donde convergen distintas ecorregiones, generando ecotonos, se encuentra lo realizado por Fagúndez et al. (2016). De los antecedentes citados, se observa que la principal oferta de floración corresponde a especies nativas, salvo en la ecoregión de la Pampa (Gurini & Basilio, 1995; Telleria, 1993), y en la provincia de Santa Cruz, en la transición entre la Estepa y Bosque Patagónico (Forcone & Kutscher, 2006; Forcone & Muñoz, 2009), donde se observa que la vegetación exótica adquiere mayor relevancia. En todos los trabajos mencionados, las familias que presentan mayor riqueza de especies son Asteraceae y Fabaceae. En cuanto a los periodos de floración, éstos se extienden durante toda la temporada apícola con determinadas particularidades para cada sector, presentándose entre octubre y diciembre los picos de floración.
Los bosques subtropicales de montaña como la Selva de las Yungas en el noroeste argentino se caracterizan por una gran diversidad de recursos. En relación con las características fenológicas, al ser un área con un marcado clima estacional, la fenología de las especies vegetales también es estacional. Más del 70 % de las especies y la mayor parte de los individuos arbóreos pierden su follaje durante la estación seca, entre junio y octubre. Estos rasgos convierten a las Yungas en uno de los ecosistemas forestales más estacionales de Sudamérica. También es estacional la floración de los árboles, que mayormente se da en primavera antes de que comiencen las lluvias (Brown & Malizia, 2004). Si bien se conoce la estacionalidad de la floración para estos bosques, no existen trabajos sobre calendarios apícolas en estos ecosistemas que brinden a los apicultores de estas áreas la información base para el desarrollo de esta actividad productiva. Además, la nutrición de las abejas está basada en néctar y polen principalmente y, por lo tanto, la cantidad disponible de estos recursos es un factor limitante para el desarrollo de la colonia. Es por ello que el presente trabajo tiene como objetivo evaluar la disponibilidad de los recursos tróficos a través de un calendario de floración y establecer su utilización a partir de la relación con el contenido polínico en el polen corbicular.
Materiales y métodos
Área de estudio: El área de estudio se sitúa en la localidad de El Fuerte, ubicado en el valle conformado por los cerros Centinela y la serranía de Santa Bárbara en el sur del Departamento Santa Bárbara al Suroeste de la Provincia de Jujuy (24°15'43'' S & 64°24'50'' W). La vegetación natural se ve representada por diferentes pisos boscosos, desde el chaqueño serrano (en las partes bajas), pasando al chaco húmedo de transición (Chaco/Yungas), posteriormente al húmedo de Yungas y por último los pastizales de altura. La temperatura media anual es de 21.5 ºC y la media de junio es de 15 °C. El área presenta un clima subtropical serrano-templado con estación seca donde las precipitaciones están concentradas en el verano y oscilan entre 800-1 050 mm anuales en promedio (Braun-Wilke, 2013). La principal actividad económica es la forestación además de la producción ganadera de subsistencia. Durante los inicios de la década del 2000, un fuerte impulso de la actividad apícola en Jujuy a partir de la red Cluster del Noroeste Argentino, permitió el desarrollo y crecimiento de esta actividad en la región, nucleado principalmente por la Escuela Agrotécnica N°12.
El apiario donde se llevó a cabo los muestreos pertenece a la Escuela Agrotécnica N° 12 que forma parte de la Red de Escuelas con desarrollo apícola, iniciada en 2002, como una iniciativa de fortalecimiento de la actividad apícola y que se transformaron en verdaderos referentes del desarrollo apícola, con docentes capacitados, apiarios demostrativos y activa participación en el proceso de transferencia tecnológica a través de jornadas, cartillas, asistencia directa, trabajos de investigación en conjunto con instituciones como la Universidad Nacional de Jujuy (UNJu) y el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), entre otros (Bedascarrasbure, 2009).
Curvas de floración: La metodología empleada para el seguimiento de la fenología de la floración fue la propuesta por Anderson y Hubritch (1940), la que considera el aspecto visual que produce la floración de las distintas especies en el paisaje. Las fenofases relevadas fueron: a- inicio de floración, b- floración plena, c- fin de floración-comienzo de fructificación, registradas una vez al mes en una planilla elaborada para tal fin con excepción de enero debido a las dificultades en acceso al área de estudio por cortes de camino a consecuencia de las lluvias estivales. Los registros fenológicos fueron como mínimo de cinco individuos por especie.
Vegetación disponible y utilización: Relación vegetación-polen: Con el fin de conocer la relación entre la vegetación de los alrededores del apiario en la localidad de El Fuerte (recursos tróficos disponibles para las abejas) y el espectro polínico de las cargas corbiculares (utilización del recurso), se calculó la asociación (A), sobrerrepresentación (O) y subrepresentación (U) de los distintos tipos polínicos a través del Índice de Davis (1984), que fue adaptado con la finalidad de determinar la relación entre la presencia de un taxón en la muestra polínica y la vegetación local. En este trabajo se presenta una adaptación del índice, relacionando los tipos polínicos encontrados en las muestras de polen corbicular con el recuso disponible en cada mes relevado. Las fórmulas empleadas son: A = B0 /(P0 + P1 + B0), U = P1/(P1 + B0), O = P0/(P0 + B0). Donde, B0 es el número de veces en los que el tipo polínico está presente en la muestra y en el registro de vegetación, P0 representa el número de veces en donde el tipo polínico se encuentra presente pero el taxón vegetal está ausente y P1 es igual al número de veces donde el taxón vegetal se encuentra presente pero el tipo polínico está ausente.
Los valores del índice de asociación varían entre 0 y 1; donde A = 1 indica que el tipo polínico y el taxón vegetal están siempre presentes y si A = 0 alguno de los dos elementos se encuentra ausente en todas las muestras analizadas.
Resultados
Relevamiento de la vegetación. Determinación taxonómica, hábito y estatus: El relevamiento de la vegetación realizada permitió determinar un total de 47 especies y 9 géneros, pertenecientes a 27 familias botánicas. Las principales familias representadas a partir de su riqueza específica fueron: Asteraceae (15), Fabaceae (6), Verbenaceae (4), Solanaceae (3). Con dos especies le siguen: Anacardiaceae, Lythraceae, Malvaceae, Rhamnaceae y Sapindaceae. Las restantes 18 familias estuvieron representadas por solo una especie. En la Fig. 1 se representa el valor porcentual de las principales familias.
En la Fig. 2 se presenta la distribución por estatus de las especies relevadas, observándose la presencia de un amplio porcentaje de especies nativas (80 %).
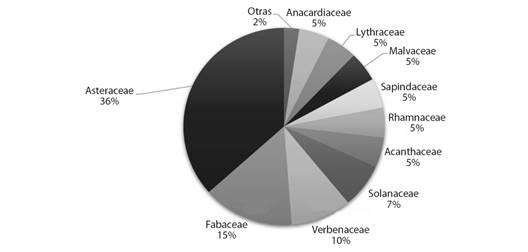
Fig. 1 Distribución porcentual de las familias botánicas relevadas en El Fuerte-Departamento Santa Bárbara, Jujuy. / Fig. 1. Percentage distribution of the botanical families surveyed in El Fuerte- Santa Barbara Department, Jujuy.
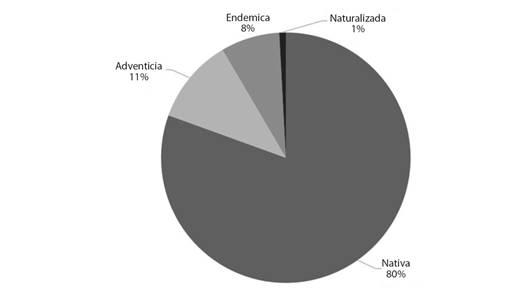
Fig. 2 Representación porcentual por estatus del relevamiento de especies vegetales realizado en El Fuerte-Departamento Santa Bárbara, Jujuy. / Fig. 2. Percentage representation by status of the survey of plant species carried out in El Fuerte-Santa Bárbara Department, Jujuy.
Registros fenológicos. Curvas de floración: En la Tabla 1 se presentan los períodos de floración de las especies identificadas en el área de estudio, en la que se indican las fenofases: inicio, plenitud y fin de floración.
Tabla 1 Registros de fenología de floración de las especies registradas en los alrededores del apiario en la localidad de El Fuerte (Departamento Santa Barbara, Jujuy). Table 1. Records of flowering phenology of the species recorded in the surroundings of the apiary in the town of El Fuerte (Santa Barbara Department, Jujuy)
| Familia | Nombre Científico | Hábito | 2014-2015 | 2015-2016 | ||||||||||
| S | O | N | D | F | M | S | O | N | D | F | M | |||
| Acanthaceae | Justicia mandonii (Lindau) Wassh. & C. Ezcurra | Arbusto | ||||||||||||
| Stenandrium dulce (Cav.) Nees | Hierba | |||||||||||||
| Amaranthaceae | Amaranthus hybridus L. | Hierba | ||||||||||||
| Anacardiaceae | Schinus sp. | Árbol | ||||||||||||
| Schinus myrtifolius (Griseb.) Cabrera | Arbusto o arbolito | |||||||||||||
| Apocynaceae | Mandevilla laxa (Ruiz & Pav.) Woodson | Enredadera o liana | ||||||||||||
| Asteraceae | Baccharis salicifolia (Ruiz & Pav.) Pers. | Arbusto | ||||||||||||
| Barnadesia odorata | Arbusto | |||||||||||||
| Bidens andicola Kunth | Hierba | |||||||||||||
| Fleischmannia prasiifolia (Griseb.) R.M. King & H. Rob. | Subarbusto | |||||||||||||
| Jungia pauciflora Rusby | Hierba o subarbusto | |||||||||||||
| Mikania cordifolia (L. f.) Willd. | Enredadera o liana | |||||||||||||
| Onoseris alata Rusby | Hierba | |||||||||||||
| Senecio hieronymi Griseb. | Subarbusto | |||||||||||||
| Senecio rudbeckiiefolius Meyen & Walp. | Arbusto | |||||||||||||
| Stevia sp. | Hierba | |||||||||||||
| Stevia vaga Griseb. | Subarbusto | |||||||||||||
| Stevia yaconensis Hieron. | Hierba | |||||||||||||
| Taraxacum officinale F.H. Wigg. | Hierba | |||||||||||||
| Vernonanthura squamulosa (Hook. & Arn.) H. Rob. | Arbusto | |||||||||||||
| Zinnia peruviana (L.) L. | Hierba | |||||||||||||
| Berberidaceae | Berberis commutata Eichler | Arbusto | ||||||||||||
| Boraginaceae | Cynoglossum amabile Stapf & J.R. Drumm. | Hierba | ||||||||||||
| Brassicaceae | Descurainia erodiifolia (Phil.) Prantl ex Reiche | Hierba | ||||||||||||
| Bromeliaceae | Tillandsia tenuifolia L. | Hierba Epífita | ||||||||||||
| Euphorbiaceae | Croton sp. | Hierba | ||||||||||||
| Fabaceae | Collaea argentina Griseb. | Subarbusto | ||||||||||||
| Desmodium sp. | ||||||||||||||
| Parasenegalia visco (Lorentz ex Griseb.) Seigler & Ebinger | Árbol | |||||||||||||
| Senna sp. | ||||||||||||||
| Vachellia aroma Gillies ex Hook. & Arn. | Arbusto o arbolito | |||||||||||||
| Vachellia caven (Molina) Seigler & Ebinger | Arbusto o arbolito | |||||||||||||
| Iridaceae | Herbertia tigridioides (Hicken) Goldblatt | Hierba | ||||||||||||
| Lamiaceae | Cantinoa mutabilis (Rich.) Harley & J.F.B. Pastore | Hierba | ||||||||||||
| Loasaseae | Caiophora clavata Urb. & Gilg | Hierba | ||||||||||||
| Lythraceae | Heimia salicifolia (Kunth) Link | Arbusto o subarbusto | ||||||||||||
| Cuphea sp. | Hierba | |||||||||||||
| Malvaceae | Modiolastrum malvifolium (Griseb.) K. Schum. | Hierba | ||||||||||||
| Sida rhombifolia L. | Hierba o subarbusto | |||||||||||||
| Malpigiaceae | Heteropterys dumetorum (Griseb.) Nied. | Liana | ||||||||||||
| Myrtaceae | Blepharocalyx salicifolius (Kunth) O. Berg | Arbusto o árbol | ||||||||||||
| Papaveraceae | Argemone subfusiformis G.B. Ownbey | Hierba | ||||||||||||
| Ranunculaceae | Clematis sp. | Liana | ||||||||||||
| Rhamnaceae | Condalia buxifolia Reissek | Arbusto o árbol | ||||||||||||
| Colletia spinossisima J.F. Gmel. | Arbusto | |||||||||||||
| Rosaceae | Cydonia oblonga | Árbol | ||||||||||||
| Rutaceae | Zanthoxylum coco Gillies ex Hook. f. & Arn. | Árbol | ||||||||||||
| Sapindaceae | Allophylus edulis (A. St.-Hil., A. Juss. & Cambess.) Hieron. ex Niederl. | Arbusto o árbol | ||||||||||||
| Serjania sp. | ||||||||||||||
| Solanaceae | Cestrum sp. | Arbusto | ||||||||||||
| Solanum sp. | ||||||||||||||
| Verbenaceae | Aloysia gratissima (Gillies & Hook. ex Hook.) Tronc. | Arbusto | ||||||||||||
| Duranta serratifolia (Griseb.) Kuntze | Arbusto o árbol | |||||||||||||
| Glandularia cabrerae (Moldenke) Botta | Hierba | |||||||||||||
| Glandularia scrobiculata (Griseb.) Tronc. | Hierba | |||||||||||||
| Vitaceae | Cissus tweediana (Baker) Griseb. | Liana | ||||||||||||
La intensidad del color gris indica los estados:  INICIO,
INICIO,  PLENITUD,
PLENITUD,  FINALIZACION. / The intensity of the gray color indicates the states: START, FULLNESS, END.
FINALIZACION. / The intensity of the gray color indicates the states: START, FULLNESS, END.
En las Fig. 3 y Fig. 4 se representa la oferta de floración durante los periodos apícolas estudiados y la variación de la distribución por hábito de las especies registradas en el sitio. Se observa en el inicio del periodo 2014-2015 (setiembre y octubre de 2014) una moderada oferta de floración representada en forma equitativa por el estrato arbóreo arbustivo y herbáceo. Avanzado en la primavera se observa un pico de floración en noviembre. Hacia diciembre 2014 y febrero 2015 se presenta una baja disponibilidad de recursos florales, reiterándose un pico de floración a fines de la temporada (marzo 2015). Mientras que para el periodo 2015-2016 el inicio presenta una buena oferta de floración que va disminuyendo hasta diciembre. Hacia fines de la época estival (marzo) la oferta de floración tiene un incremento, presentando un pico al igual que el periodo anterior a fines de la temporada (marzo 2016). En ambos periodos, el incremento de la oferta de floración hacia el final de la temporada se caracteriza por la mayor disponibilidad de especies herbáceas.
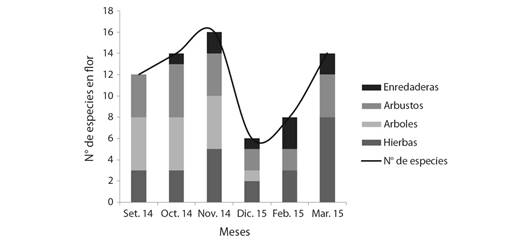
Fig. 3 Curva de floración para la localidad de El Fuerte. Periodo setiembre 2014-marzo 2015. / Fig. 3. Flowering curve for the town of El Fuerte. Period September 2014-March 2015.
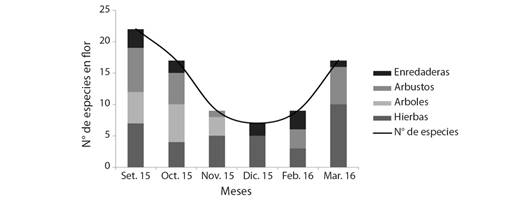
Fig. 4 Curva de floración para la localidad de El Fuerte. Periodo setiembre 2015-marzo 2016. / Fig. 4. Flowering curve for the locality of El Fuerte. September 2015-March 2016 period.
Para complementar la información, en la Fig. 5 se observa que, frente a una baja disponibilidad de especies en flor, la abeja utiliza una mayor diversidad de recursos polínicos (Riqueza polínica), siendo en condiciones opuestas menor la riqueza utilizada.
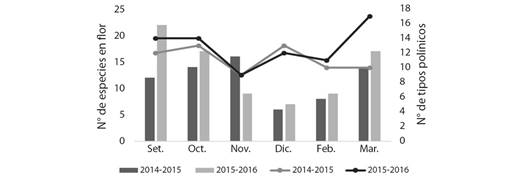
Fig. 5 Distribución de la riqueza polínica y el N° de especies en flor identificadas durante los periodos de estudio en la localidad de El Fuerte (Departamento Santa Bárbara, Jujuy). / Fig. 5. Distribution of pollen richness and the number of flower species identified during the study periods in the town of El Fuerte (Santa Bárbara Department, Jujuy).
Relación vegetación-polen: Se calculó el índice de asociación de Davis, utilizando los datos de presencia-ausencia de los 64 taxones presentes en los registros de vegetación y polen. De acuerdo con los valores obtenidos, cinco de ellos presentan una asociación media a alta entre la especie vegetal disponible y la presencia de ésta en el espectro polínico de la muestra de polen corbicular recolectada. Estos recursos son: Vachellia aroma, B. salicifolius, Cantinoa sp., Vernonanthura sp. y Zanthoxylum coco (Tabla 2).
Tabla 2 Índices de asociación (A), Subrepresentación (U) y Sobrerrepresentación (O) adaptados de Davis (1984) para la localidad de El Fuerte (Departamento Santa Bárbara, Jujuy). Table 2. Association (A), Underrepresentation (U) and Overrepresentation (O) indices adapted from Davis (1984) for the town of El Fuerte (Santa Bárbara Department, Jujuy)
| Especies | Índices | Especies | Índices | ||||
| A | U | O | A | U | O | ||
| Allophylus edulis | 0.17 | 0.67 | 0.75 | Glandularia cabrerae | 0 | 1 | 0 |
| Aloysia gratissima | 0 | 1 | 0 | Glandularia scrobiculata | 0 | 1 | 0 |
| Amaranthus hybridus | 0.17 | 0.5 | 0.8 | Heimia salicifolia | 0 | 1 | 0 |
| Anadenanthera colubrina | 0 | 0 | 1 | Herbertia tigridioides | 0 | 1 | 0 |
| Apiaceae | 0 | 0 | 1 | Heteropterys dumetorum | 0 | 1 | 0 |
| Argemone subfusiformis | 0 | 1 | 0 | Jungia pauciflora | 0 | 1 | 0 |
| Baccharis sp. | 0.22 | 0.67 | 0.6 | Justicia mandonii | 0 | 1 | 0 |
| Barnadesia odorata | 0 | 1 | 0 | Lithraea molleoides | 0 | 0 | 1 |
| Berberis commutata | 0 | 1 | 0 | Mandevilla laxa | 0 | 1 | 0 |
| Bidens andicola | 0 | 1 | 0 | Mikania cordifolia | 0 | 1 | 0 |
| Bignoniaceae | 0 | 0 | 1 | Modiolastrum malvifolium | 0 | 1 | 0 |
| Blepharocalyx salicifolius | 0.57 | 0.2 | 0.33 | Onoseris alata | 0 | 1 | 0 |
| Brassicaceae | 0.25 | 0 | 0.75 | Parapiptadenia excelsa | 0 | 0 | 1 |
| Caiophora clavata | 0 | 1 | 0 | Parasenegalia visco | 0 | 1 | 0 |
| Cantinoa mutabilis | 0.67 | 0.33 | 0 | Schinus myrtifolius | 0 | 1 | 0 |
| Carduus thoermeri | 0 | 0 | 1 | Schinus sp. | 0 | 1 | 0 |
| Cestrum sp. | 0 | 1 | 0 | Senecio sp. | 0.4 | 0.6 | 0 |
| Cissus sp. | 0.4 | 0.5 | 0.33 | Senna sp. | 0 | 1 | 0 |
| Clematis sp. | 0 | 1 | 0 | Serjania sp. | 0 | 1 | 0 |
| Collaea argentina | 0 | 1 | 0 | Sida rhombifolia | 0 | 1 | 0 |
| Colletia spinossisima | 0 | 1 | 0 | Solanum sp. | 0.43 | 0.25 | 0.5 |
| Condalia buxifolia | 0 | 1 | 0 | Stenandrium dulce | 0 | 1 | 0 |
| Croton sp. | 0.09 | 0.88 | 0.75 | Stevia sp. | 0 | 1 | 0 |
| Cucurbitaceae | 0 | 0 | 1 | Stevia vaga | 0 | 1 | 0 |
| Cuphea sp. | 0 | 1 | 0 | Stevia yaconensis | 0 | 1 | 0 |
| Cydonia oblonga | 0 | 1 | 0 | Taraxacum officinale | 0 | 1 | 0 |
| Cynoglossum amabile | 0 | 1 | 0 | Tilandsia tenuifolia | 0 | 1 | 0 |
| Descurainia erodiifolia | 0 | 1 | 0 | Vachellia aroma | 0.5 | 0 | 0.5 |
| Desmodium sp. | 0 | 1 | 0 | Vachellia caven | 0 | 1 | 0 |
| Duranta serratifolia | 0 | 1 | 0 | Vernonanthura | 0.5 | 0.5 | 0 |
| Elephantopus mollis | 0 | 0 | 1 | Zanthoxylum coco | 0.5 | 0 | 0.5 |
| Fleischmannia prasiifolia | 0 | 1 | 0 | Zinnia peruviana | 0 | 1 | 0 |
Cabe destacar, que existe una sobrerepresentación (O = 1) de polen correspondiente a taxones como Anadenanthera colubrina, Berberis sp., Carduus thoermeri, Elephantopus mollis, Lithraea molleoides, Parapiptadenia excelsa, Apiaceae y Cucurbitaceae. Es decir, hay ausencia de los individuos de la especie (en la vegetación) en el radio de influencia al apiario donde se realizaron los registros y colecciones de los recursos florales disponibles. Por otra parte, se registró la ausencia dentro de los recursos polínicos utilizados (espectro polínico) o Subrepresentación (U) de diversos taxa en su mayoría arbustivos y herbáceos como Aloysia gratissima, Barnadesia odorata, Caiophora clavata, Collaea argentina, Colettia spinosissima y Descurainia erodifolia, entre otros, que si se encuentran en la comunidad vegetal circundante al apiario.
Discusión
A partir del relevamiento realizado, se determinó que al igual a otras áreas apícolas de Argentina, las familias botánicas que presentaron mayor riqueza de especies son Asteraceae y Fabaceae (Cabrera et al., 2013; Fagúndez et al., 2016; Forcone & Muñoz, 2009; Naab & Tamame, 2007; Salgado et al., 2014). Estos resultados reflejan la importancia de estas familias tanto a nivel global como regional. En el caso de las Asteraceae, es la familia de fanerógamas con mayor diversidad a nivel mundial, representa 8-10 % de la flora global y abarca alrededor de 22 750 taxones específicos e infraespecíficos (del Vitto & Petenatti, 2009; Zuloaga et al., 2014). Además, es de distribución cosmopolita, ocupando hábitats diversos, desde el nivel del mar hasta el límite altitudinal de la vegetación. En Argentina, también es la más numerosa de las familias de plantas superiores con el mayor porcentaje de endemismos, al igual que en la provincia de Jujuy en la que constituye el 16 % de su flora (Cabrera, 1961; Katinas et al., 2007; Zuloaga, 1999; Zuloaga et al., 1999).
En el caso de la familia Fabaceae, también es una familia con un gran de número de especies distribuidas en todo el mundo, sobre todo en ambientes tropicales y subtropicales (Burkart et al., 1987), y en Argentina, se distribuye preferentemente en el centro y norte del país, representando el 6 % de la flora en la provincia de Jujuy. En esta familia la mayoría de las especies poseen flores hermafroditas y entomófilas en las cuales los himenópteros son los principales agentes de polinización (Burkart, 1952; Burkart et al., 1987; Figueroa-Fleming, 2014; Figueroa-Fleming & Etcheverry, 2017; Zuloaga et al., 1999).
El período de disponibilidad de recursos alimentarios del área circundante al apiario en la localidad de El Fuerte (Departamento Santa Bárbara, Jujuy), se extiende durante toda la temporada apícola (setiembre-marzo), situación que también se observa en otras áreas apícolas del país (Forcone, 2003; Forcone & Kutschker, 2006; Tellería, 1993). Al inicio de la temporada, en ambos períodos la oferta de floración esta entre moderada y alta y es representada principalmente por especies nativas de los diferentes estratos entre los que se encuentran: Allophylus edulis, Zanthoxylum coco, Vachellia aroma, Cestrum sp., Senecio rudbeckiiefolius y Vernonanthura squamulosa. En el caso particular del período 2015-2016, el número de especies en flor declina hacia mediados de verano. Esta tendencia en la marcha de floración se observa también en otras áreas subtropicales del noroeste argentino (NOA) como lo es el sector del chaco húmedo (Salgado et al., 2014). Además, es necesario destacar que durante estos meses se presentan bajos valores de precipitación que condicionarían la disminución de los recursos para el área estudiada. En relación a la máxima oferta (mayor número de especies en floración plena), para la temporada 2014-2015, noviembre registró disponibilidad de especies arbóreas como Allophylus edulis, Blepharocalyx salicifolius, Condalia buxifolia, Vachellia aroma; y arbustivas tales como: Baccharis salicifolia, Aloysia gratissima, Cestrum sp. y herbáceas como Descurainia erodifolia, Croton sp., Glandularia scrobiculata, Solanum sp., Argemone subsiformis, entre otras. La similitud temporal en relación con el pico de floración también se presenta en la zona del Espinal (Andrada, 2003), el Monte de Llanuras y Mesetas, en la provincia de la Pampa (Naab & Tamame, 2007), y en el noroeste de Santa Cruz (Forcone & Muñoz, 2009). Por otro lado, en ambos periodos hacia fines de la temporada (marzo) se observa un segundo pico de floración en el que el principal aporte está dado por las especies herbáceas. Este comportamiento en la comunidad vegetal permite a los productores apícolas en general llevar a cabo una segunda cosecha finalizando la temporada apícola (obs. pers.).
Si bien solo se cuenta con los registros de floración para el periodo apícola (setiembre a marzo), es en esta época donde se produce la mayor floración por la marcada estacionalidad en respuesta a las variaciones climáticas en el bosque de Yungas (Brown & Malizia, 2004). Sin embargo, en otras regiones de importancia apícola para el país como los son el Chaco y Entre Ríos, la floración se extiende durante todo el año (Cabrera et al., 2013; Fagúndez et al., 2016; Salgado et al., 2014), complementado en el caso de Entre Ríos, con la oferta de especies adventicias o naturalizadas.
La utilización de los recursos disponibles en los alrededores del apiario en la localidad de El Fuerte (Santa Bárbara, Jujuy) es baja. Esto se refleja en el valor del índice de similitud de Jaccard (Ij = 0.11), que relaciona en este caso, los recursos utilizados por A. mellifera y la flora disponible y registrada durante las temporadas estudiadas. Es decir que la abeja solo recolecta polen de algunas especies presentes en sus alrededores. Una situación similar se observó en tres apiarios de la localidad de Diamante, Entre Ríos (Fagúndez, 2011). En cambio, en otras regiones del país, Forcone (2002) y Andrada y Tellería (2005) observaron una mayor similitud entre el número de tipos polínicos recolectados y la oferta de la floración.
En cuanto al uso particular de los recursos, si bien los índices de asociación (A), Sobrerepresentación (O) y Subrepresentación (U) adaptados de Davis (1984), se calcularon a partir de datos de presencia y ausencia de los taxa en las muestras de polen corbicular y en los registros de vegetación, los valores obtenidos muestran una tendencia sobre su utilización. Los recursos Cantinoa sp. (0.66), B. salicifolius (0.57), Vachellia aroma (0.5), Zanthoxylum coco (0.5) y Vernonanthura sp. (0.5) son los que presentaron mayor valor de asociación, es decir que, frente a la presencia del recurso, éste es pecoreado por las abejas. Como se observa, la mayoría de ellos son especies arbóreas y su elección se podría explicar a partir de lo planteado por Ramalho (2004), quien demuestra que, las abejas sin aguijón del Bosque Tropical Atlántico, tienen preferencia por el estrato superior (arbóreo), con floración masiva, caracterizada por una alta densidad de flores pequeñas dispuestas en numerosas inflorescencias, característica que se observa en las especies identificadas en este trabajo, como recurso floral utilizado por A. mellifera. También se conoce que el costo energético en la obtención del recurso es un factor que influye en su selección, al igual que la cantidad de polen producida por las flores y la disposición del polen dentro de la flor, esto se refleja en cómo A. mellifera y Bombus prefieren forrajear en flores donde el polen es abundante y se puede obtener de forma rápida y sencilla (Pernal & Currie, 2002; Rasheed & Harder, 1997; de Sá-Otero et al., 2004). Sin embargo, en el espectro polínico aparecen numerosos recursos sobrerepresentados, es decir que no se encuentran en el radio de vegetación de influencia relevado (O = 1). Entre ellos se destacan por su mayor abundancia y/o frecuencia en las muestras polínicas: Gleditsia amorphoides, Anadenanthera colubrina y Parapiptadenia excelsa. Su presencia podría estar relacionada con la ampliación del área de forrajeo de la abeja, llegando hasta distancias que pueden superar los 10 km de la colmena (Beekman & Ratnieks, 2000; Roubik, 1992; Steffan-Dewenter & Kuhn, 2003; Visscher & Seeley, 1982), además de que todas ellas presentan amplios periodos de floración (Cazón, 2009; Digilio & Legname, 1966), criterio que también es considerado para su selección (Avni et al., 2009; Girón-Vanderhuck, 1995; Montoya-Pfeiffer, 2011). En contraposición se encuentran los recursos subrepresentados (U= 1), es decir aquellos que se encuentran disponibles en el área relevada pero que la abeja no recolecta. Entre éstos encontramos en su mayoría a especies herbáceas y arbustivas como Aloysia grattisima, Duranta serratifolia, Barnadesia odorata, Caiophora clavata, Collaea argentina, Colletia spinosissima, Descurainia erodifolia, Glandularia scrobiculata y Sida rhombifolia, Cestrum sp., entre otras. Esta situación podría atribuirse a factores de selección o atracción floral como la forma de flor (síndrome de entomofilia), cantidad de polen producida y disposición de los estambres en la flor (Pernal & Currie, 2002; Rasheed & Harder, 1997). Por ejemplo, en el caso de A. gratissima, C. spinosissima, G. scrobiculata, D. serratifolia y Cestrum sp., los estambres se encuentran incluidos en una corola tubular poco accesible para los visitantes florales. Por otro lado, se puede considerar que algunas de estas especies junto a Descurainia erodifolia y Sida rhombifolia presentan sus periodos de floración coincidente con especies arbóreas de floración masiva como B. salicifolius por ejemplo, situación que también podría estar influenciando en su elección (Ramalho, 2004).
Un aspecto importante a destacar es lo planteado por McLellan (1976) y Steffan-Dewenter y Kuhn (2003) en referencia a la riqueza de especies que usa la abeja frente a la disponibilidad de los mismos; dichos autores han documentado que las abejas visitan un mayor número de especies vegetales bajo condiciones de escasez de recursos en el entorno y un menor número de recursos en ambientes más abundantes, como se observa en El Fuerte en los periodos estudiados (Fig. 5).
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

