











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La región oriental de Colombia, denominada Orinoquía, es una extensa zona de sabanas que ocupa el 22 % del territorio nacional, en la cual existe un amplio desconocimiento de su biodiversidad y los procesos que rigen sus dinámicas ecológicas (Instituto Alexander Von Humboldt, 2009; Rippstein et al., 2001). Según la clasificación biogeográfica de Morrone (2014) pertenece a la provincia de Sabana, de la subregión caribeña, de la Región Neotropical; es el mayor ecosistema de sabanas de América, donde existen bosques secos, húmedos, de galería, pastizales y humedales. Como se presenta en muchos ecosistemas del planeta, la Orinoquía está expuesta a una rápida reducción de la biodiversidad por acciones antrópicas (Instituto Alexander Von Humboldt, 2009; Morrone, 2001). Esta prospección de transformaciones en curso y futuras generan la necesidad de establecer medidas para la protección de las especies, para lo cual es esencial documentar la biodiversidad (Anderson, 2018; Larsen et al., 2018).
En los ecosistemas terrestres los insectos son el grupo con mayor éxito evolutivo (Purvis & Hector, 2000) por su abundancia, diversidad, amplio espectro de hábitats y roles funcionales que desarrollan (Llorente-Bousquets et al., 1996; Mattoni et al., 2000). Cuatro órdenes de insectos, entre ellos los dípteros, contribuyen con más del 50 % de la diversidad animal y por tanto son considerados megadiversos (Zhang, 2011a; Zhang, 2011b). Los dípteros son excelentes candidatos para documentar detalladamente sobre la heterogeneidad ecológica (Kitching et al., 2005; Smith & Mayfield, 2015) dado que participan en una gran variedad de interacciones bióticas y procesos ecosistémicos (Rivera-García & Viggers-Carrasco, 1991; Kitching et al., 2005; Smith & Mayfield, 2015), además que interactúan de forma positiva o negativa con las poblaciones humanas como polinizadores, vectores de enfermedades, o como competidores, siendo algunos de ellos plagas de los cultivos agrícolas.
La superfamilia Tephritoidea del orden Diptera incluye ocho familias de moscas y en sus especies existen hábitos alimenticios como saprófagía, depredadores de huevos y larvas de insectos, parasitoides y fitófagía. Estos últimos, conocidos como moscas de las frutas, han atraído gran atención por las pérdidas económicas que causan en la agricultura (Korneyev, 1999). En Colombia se han documentado 224 especies de Tephritoidea distribuidas en 58 géneros (Kameneva et al., 2017; Rodríguez et al., 2018; Wolff et al., 2016), la mayoría de ellos provenientes de recolectas esporádicas. Levantamientos sistemáticos de este grupo de insectos se han direccionado a las especies de importancia económica y sólo en siete de los 32 departamentos, la mayoría de ellos localizados en la Región Andina (Castañeda et al., 2010; Cruz et al., 2017; Ruiz-Hurtado et al., 2013). En los Llanos Orientales, incluyendo el Departamento de Arauca, la información sobre la superfamilia es muy escasa y se limitan a estudios de presencia/ausencia de moscas de las frutas presentados por el ICA (Plan Nacional Moscas de las Frutas, 2012). Además, en realidad no encontramos evidencias de levantamientos sistemáticos de Tephritoidea en Colombia o en la provincia de Sabana en general.
Este estudio se desarrolló con el fin de caracterizar las comunidades de Tephritoidea en dos localidades del Departamento de Arauca, en la Orinoquía colombiana, como aporte al conocimiento de la biodiversidad en la provincia biogeográfica de Sabana y, además, por primera vez, ofrecer información ecológica de las especies de importancia económica en la región.
Materiales y métodos
Área de estudio: El estudio se realizó entre agosto 2017 y febrero 2018 en los municipios de Saravena y Arauca en el Departamento de Arauca. El primero de ellos se encuentra en una región conocida como piedemonte llanero y corresponde a la zona de transición de la Sabana con la región Andina, y la segunda está localizada en la región de Sabana, ambas localidades pertenecientes a la provincia de Sabana según la clasificación biogeográfica de Morrone (2014). Estas regiones presentan un régimen de lluvias unimodal bi-estacional con dos épocas contrastantes de sequía (el último y primer trimestre del año) y de lluvias (abril a octubre) (Rippstein et al., 2001) y con climas que de acuerdo con la clasificación de Thornthwaite que van desde súper húmedos (ARA') hasta moderadamente secos (DRA') (Minorta-Cely & Rangel-Ch, 2014). En la zona rural de cada localidad se seleccionaron dos ambientes, un huerto doméstico y un fragmento de bosque. En los huertos domésticos había actividad humana con presencia de árboles frutales y los fragmentos de bosque fueron zonas de presencia de vegetación nativa poco intervenida. Todas las áreas muestreadas contaban aproximadamente con dos hectáreas de terreno. Para facilitar la comparación de la fauna, ambos tipos de ambientes estaban cercanos, con altitudes entre 127 y 272 m. Las localizaciones geográficas de los huertos fueron en Saravena el huerto doméstico (HDSar) (6°57'47.23'' N & 71°55'10.38'' W) y el fragmento de bosque (PBSar) (6°57'4.3'' N & 71°54'54.08'' W). En Arauca el huerto doméstico (HDAra) (7°0'41.45'' N & 70°44'33.24'' W) y el fragmento de bosque (PBAra) (7°0'30.9'' N & 70°44'31.8'' W).
Recolecta de insectos: En cada área de muestreo se instalaron dos trampas tipo McPhail (Multilure, Better World, Fresno, CA, USA). Cada trampa fue cebada con 200 ml de 10 % de proteína hidrolizada de maíz y se colgaron en árboles a 1.5 m de altura. Las trampas se revisaron cada siete días, se separaron las moscas y se preservaron en alcohol al 70 %. Una vez al mes se recolectaron los frutos que estaban disponibles al momento de la revisión de las trampas y que pudieran ser hospederos de moscas de las frutas, bien fuera recolectados directamente de la planta o frutos caídos. Las frutas recolectadas se separaron por especie y fueron transportadas al laboratorio donde se pesaron y contaron para calcular los índices de infestación. Posteriormente los frutos se colocaron en bandejas con vermiculita húmeda como sustrato de empupación para las larvas. Una semana después los frutos fueron disectados y las larvas o pupas de moscas fueron transferidas a cámaras de emergencia con el mismo sustrato para la obtención de moscas adultas, las cuales se conservaron en alcohol al 70 % para su identificación.
Las moscas fueron llevadas al laboratorio de entomología de la Universidad del Tolima en Ibagué, Tolima, Colombia. La identificación de las especies fue corroborada por uno de los autores, el experto N.A. Canal. Se utilizaron claves específicas y descripciones taxonómicas para géneros y especies de cada una de las familias recolectadas.
La determinación de las plantas hospederas se realizó en el Herbario Orinocense (HORI). Especímenes voucher de las moscas se depositaron en la Colección Entomológica de la Orinoquía (CEO) y en la Colección Entomológica del Museo Laboratorio de Entomología de la Universidad del Tolima (MENT-UT).
Análisis de datos: Cada área muestreada fue considerada como una comunidad diferente para efectos de análisis de la diversidad alfa y beta. La caracterización de la comunidad de Tephritoidea se realizó con los especímenes capturados en trampas. Para estimar la riqueza de especies de la comunidad y evaluar su representatividad se usaron los estimadores no paramétricos Chao 1 y Jacknife 1 calculados con el programa EstimateS 9.1.0 (Colwell, 2013) y los resultados se representaron como curvas de acumulación de especies entre áreas y municipios. Se realizaron pruebas de Shapiro-Wilk y F para verificar la normalidad y homogeneidad de los conjuntos de datos y una prueba U de Wilcoxon para comparar las abundancias y riquezas por localidad y ambiente. Para el análisis de la diversidad beta se elaboró una matriz con las especies recolectadas por sitio de estudio, se calculó el índice de diversidad Margalef y el índice de similitud entre los sitios de muestreo con el coeficiente de Jaccard (Magurran, 1989) utilizando el método de agrupación UPGMA en el Programa Past3 (Hammer et al., 2001).
Se utilizó el índice de infestación, el cual puede ser calculado como el número de larvas por kg de fruto (Instituto Colombiano Agropecuario, 2005) y permite establecer los hospederos reales de las especies de moscas presentes en los frutos, (Rodríguez et al., 1996). También permite conocer el daño directo ocasionado por las moscas y es indicador de la densidad actual y/o futuro nivel poblacional de adultos (Instituto Colombiano Agropecuario, 2005). Se calculó el índice de Mosca Trampa Día (MTD) el cual constituye un índice poblacional del número promedio de moscas capturadas en una trampa en un día de exposición de la trampa en el campo generando una medida relativa del tamaño de la población de moscas adultas en un espacio y tiempo determinados (Instituto Colombiano Agropecuario, 2011).
Resultados
Diversidad de moscas: Se recolectaron 878 ejemplares, de los cuales 742 (84.5 %) fueron capturados con las trampas McPhail y 136 (15.5 %) de frutos hospederos (Tabla 1). Se encontraron especímenes de cuatro familias: Tephritidae con 27.2 % de los individuos, Lonchaeidae con 16 %, Richardiidae con 14.7 % y Ulidiidae 40.1 %. Los individuos de la familia Ulidiidae, dada la dificultad taxonómica en el neotrópico, no fueron incluidos en el estudio. Se identificaron quince géneros, siete de Tephritidae, dos de Lonchaeidae y seis de Richardiidae y 27 especies, siendo doce especies de la familia Tephritidae (44.4%); cuatro de Lonchaeidae (14.8 %) y once de Richardiidae (40.7 %) (Tabla 1).
Tabla 1 Frecuencia y distribución de las especies de Tephritoidea en Arauca y Saravena en el departamento de Arauca, Colombia. / Table 1. Frequency and distribution of the species of Tephritoidea in Arauca and Saravena in the department of Arauca, Colombia.
| Municipio de Saravena | Municipio de Arauca | Total | ||||||||||||||
| HD | PB | HD | PB | |||||||||||||
| Familia | Total de adultos | Especie | Trampa | Fruto | Trampa | Fruto | Trampa | Fruto | Trampa | Fruto | ||||||
| Tephritidae | 36 | 84 | Anastrepha bezzii + (Lima 1934) | 2 | 2 | |||||||||||
| Anastrepha obliqua + (Macquart 1835) | 96 | 1 | 4 | 101 | ||||||||||||
| Anastrepha parishi + (Stone 1942) | 1 | 10 | 11 | |||||||||||||
| Anastrepha striata + (Schiner 1868) | 2 | 3 | 5 | |||||||||||||
| Anastrepha sp. | 1 | 1 | ||||||||||||||
| 9 | Baryplegma sp. | 9 | 9 | |||||||||||||
| 2 | Blepharoneura sp1 | 1 | 1 | |||||||||||||
| Blepharoneura sp2 | 1 | 1 | ||||||||||||||
| 19 | 18 | Euarestopsis paupera Hering 1937 | 18 | 17 | 1 | 1 | 37 | |||||||||
| 1 | Hexachaeta sp. | 1 | 1 | |||||||||||||
| 34 | 26 | Neotaracia imox (Bates 1934) | 1 | 58 | 1 | 60 | ||||||||||
| 1 | 9 | Xanthaciura chrysura (Thomson 1869) | 1 | 1 | 4 | 4 | 10 | |||||||||
| Lonchaeidae | 55 | 69 | Neosilba batesi + (Curran 1932) | 1 | 5 | 115 | 121 | |||||||||
| Neosilba zadolicha + McAlpine & Steyskal 1982 | 3 | 3 | ||||||||||||||
| 8 | 8 | Lonchaea longicornis + Williston (1896) | 9 | 9 | ||||||||||||
| Lonchaea sp. | 1 | 6 | 7 | |||||||||||||
| Richardiidae | 4 | 19 | Coilometopia sp. | 17 | 6 | 23 | ||||||||||
| 3 | Euolena sp. | 1 | 2 | 3 | ||||||||||||
| 1 | Hemixantha lutea Hennig 1937 | 1 | 1 | |||||||||||||
| 20 | 6 | Melanoloma affinis Loew 1873 | 4 | 4 | ||||||||||||
| Melanoloma cyanogaster Wiedeman 1830 | 19 | 19 | ||||||||||||||
| Melanoloma varians (Schinner 1868) | 2 | 2 | ||||||||||||||
| Melanoloma sp. | 1 | 1 | ||||||||||||||
| 9 | 17 | Richardia podagrica (Fabricius 1805) | 19 | 2 | 1 | 2 | 24 | |||||||||
| Richardia sp1. | 1 | 1 | ||||||||||||||
| Richardia sp2. | 1 | 1 | ||||||||||||||
| 19 | 31 | Sepsisoma. Anale (Schiner 1868) | 34 | 1 | 14 | 1 | 50 | |||||||||
| Total por sexo | 220 | 288 | Total por área | 191 | 75 | 231 | 11 | |||||||||
| Total | 508 | Total por municipio | 266 | 242 | 508 | |||||||||||
HD: Huerto doméstico. PB: Parche de bosque.
Las especies más abundantes en cada una de las familias fueron: Anastrepha obliqua (19.8 %), Neosilba batesi (23.8 %) y Sepsisoma anale (9.8 %) respectivamente (Tabla 1).
Diversidad Alfa: En Saravena se recolectaron 23 especies (PBSar = 19; HDSar = 11 y exclusivas = 13) y en Arauca 14 especies (PBAra = 6; HDAra = 14, exclusivas = 4). Las huertas domésticas presentaron mayor abundancia de moscas con 83 % de los especímenes recolectados (Tabla 1).
Las curvas de acumulación de especies se muestran con una tendencia asintótica, tanto en el muestreo general, como por localidades y ambientes, lo cual indica que el esfuerzo de muestreo representó adecuadamente la diversidad de la zona. La diversidad recolectada representa el 75 % de la diversidad teórica (S = 23, Chao1 = 36.2) y en cuanto a municipios y áreas representó entre 66 (S = 23, Chao1 = 35.1) y 81 % (S = 14, Chao1 = 17.4). La curva de acumulación de especies durante el tiempo de muestreo fue similar para los dos municipios presentando valores mayores entre los meses de septiembre y noviembre.
Por otro lado, las curvas de abundancia fueron diferentes entre los dos municipios; se encontró mayor riqueza de especies para Saravena entre enero y marzo (sequía) y para Arauca entre septiembre y noviembre (lluvias) (Fig. 1). Tanto la riqueza (W = 702; N =127; P < 0.05) como la abundancia de especies (W = 690; N = 127; P < 0.05) fue mayor en el municipio de Saravena que en Arauca. Respecto a los ambientes, la riqueza (W = 1861.5; N = 127; P < 0.05) fue mayor en los parches de bosque que en las huertas domésticas, mientras que la abundancia (W = 2032.5; N = 127; P < 0.05) fue mayor en las huertas domésticas que en los parches de bosque.
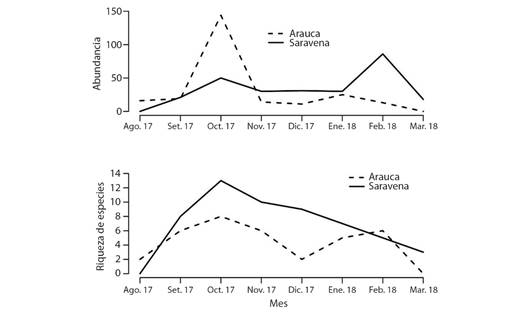
Fig. 1 Riqueza y Abundancia de especies de Tephritoidea en dos localidades del departamento de Arauca, Colombia. / Fig. 1. Richness and abundance of species of Tephritoidea in two localities of the department of Arauca, Colombia.
Diversidad Beta: Cuatro especies son comunes en todas las áreas muestreadas (Euarestopsis paupera, Xanthaciura chrysura, Richardia podagrica y Sepsisoma anale). Según el índice de similitud de Jaccard, la huerta doméstica y el parche de bosque del municipio de Arauca presentaron la mayor similitud de riqueza de especies (60 %), mientras que los ecosistemas del municipio de Saravena presentaron la menor similitud (27 %). El parche de bosque de Saravena fue el más disímil en su composición de especies (Fig. 2).
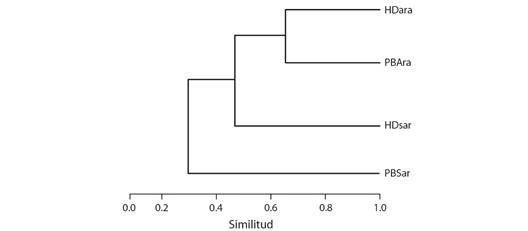
Fig. 2 Dendrograma de similitud de Jaccard (UPGMA) de acuerdo con las especies de Tephritoidea recolectadas en dos áreas diferentes de dos localidades del departamento de Arauca, Colombia. HDSar = Huerta doméstica Saravena, PBSar = Parche de bosque Saravena, HDAra = Huerta doméstica Arauca, PBAra = Parche de bosque Arauca. / Fig. 2. Jaccard similarity dendrogram (UPGMA) according to the species of the Tephritoidea collected in two different areas of two localities of the department of Arauca, Colombia. HDSar = Saravena domestic garden, PBSar = Saravena forest patch, HDAra = Arauca domestic garden, PBAra = Arauca forest patch.
Los valores de riqueza específica proporcionados por el índice de Margalef indican baja diversidad en dos de las cuatro áreas muestreadas, la huerta doméstica de Saravena (1.71) y la huerta doméstica de Arauca (1.95) y diversidad media en el parche de bosque de Arauca (2.08) y el parche de bosque de Saravena (4.00).
Especies de importancia económica: Se recolectaron 580 frutos (30.4 kg) de siete especies botánicas que se encontraron durante el periodo de muestreo y, a partir de seis de esos hospederos, emergieron 136 moscas adultas de 524 larvas, las cuales pertenecían a tres especies de Lonchaeidae y una de Tephritidae. Citrus sinensis (naranja) fue el hospedero de mayor abundancia durante el muestreo en ambos municipios, aunque también el de menor emergencia de moscas adultas. Annona muricata (guanábana) en el municipio de Arauca presentó el mayor número de emergencia de moscas adultas, mientras que en Citrus sp. (mandarina) en Saravena no se encontraron larvas. Psidium guajava (guayaba) fue el único hospedero afectado por un Tephritidae (A. striata). Los índices de infestación presentaron valores bajos, por debajo de 0.1, con excepción de Annona muricata (142.5 larvas/ kg fruto) y Miconia sp. (33.11 larvas/ kg fruto) (Tabla 2).
Tabla 2 Frutos hospederos y especies de moscas de las frutas en dos localidades del departamento de Arauca e índices de infestación. / Table 2. Host fruits and species of fruit flies in two localities of the department of Arauca and infestation indexes.
| Área | Familia | Hospederos | Nombre común | Peso total (kg) | No. de frutos | No. total de larvas | Especie de mosca de la fruta | Total de adultos | No. de muestras | Índices de infestación Larvas/kg fruto |
| Arauca | Annonaceae | Annona muricata | Guanábana | 0.80 | 1 | 114 | N. batesi N. zadolicha L. longicornis | 97 3 9 | 1 | 142.5 |
| Myrtaceae | Psidium guajava | Guayaba | 1.60 | 39 | 20 | N. batesi A. striata | 13 3 | 2 | 0.01246 | |
| Rutaceae | Swinglea glutinosa | Limoncillo | 4.23 | 24 | 9 | N. batesi | 3 | 2 | 0.00212 | |
| Rutaceae | Citrus sinensis | Naranja | 6.68 | 46 | 264 | N. batesi | 2 | 4 | 0.03952 | |
| Saravena | Laurales | Persea americana | Aguacate | 2.36 | 12 | 4 | N. batesi | 2 | 2 | 0.00169 |
| Rutaceae | Citrus sinensis | Naranja | 13.28 | 86 | 108 | N. batesi | 1 | 6 | 0.00813 | |
| Rutaceae | Citrus sp. | Mandarina | 1.30 | 17 | 0 | - | 0 | 2 | 0 | |
| Melastomataceae | Miconia sp. | Baya | 0.15 | 355 | 5 | N. batesi | 3 | 3 | 33.11 | |
| Total | 30.40 | 580 | 524 | 4 | 136 | 6 | 175.67 |
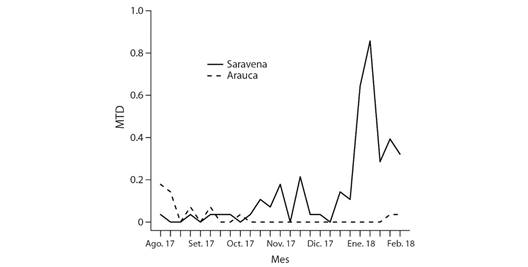
En las trampas McPhail se recolectaron 118 individuos del género Anastrepha. En Arauca diez fueron hembras y seis machos mientras que en Saravena fueron 72 hembras y 30 machos. Los índices poblacionales en las localidades estuvieron entre 0.0-0.85 (Fig. 3). En Saravena los valores del MTD más bajos se presentaron en los meses de septiembre a noviembre (época de lluvias) y los valores mayores en los meses de enero a febrero (época de sequía). En Arauca los valores máximos se presentaron en los meses de septiembre a noviembre (periodo de más lluvias) y los más bajos en los meses de enero a febrero (periodo de menos lluvias) (Fig. 3).
Discusión
El conocimiento de los patrones de distribución de las especies y sus causas son la base de la moderna biología de la conservación y, la biogeografía es una herramienta informativa potente (Morrone, 2001). La provincia biogeográfica de sabana es una amplia zona que se extiende en el oriente colombiano y occidente de Venezuela, representa uno de los mayores ecosistemas de sabana del mundo, en peligro por las actividades antrópicas, pero además poco estudiado (Morrone, 2001; Morrone, 2014), en Colombia, por ejemplo, la Orinoquía es un área donde las políticas oficiales promueven la colonización agropecuaria (Fondo para el Financiamiento del Sector Agropecuario, 2014). Las moscas tefritóideas pueden ser utilizadas como indicadores biológicos y en este trabajo se presenta por primera vez información taxonómica precisa y sistemática de las especies presentes en dos áreas de la provincia de Sabana.
Se identificaron ejemplares pertenecientes a quince géneros y 27 especies de las familias Lonchaeidae, Richardiidae y Tephritidae. Los registros de E. paupera, H. lutea, M. affinis, M. cyanogaster, M. varians y S. anale son nuevos para Colombia y para la Orinoquía colombiana se registran por primera vez A. bezzi, N. imox, X. chrysura, R. podagrica y las cuatro especies de la familia Lonchaeidae. En Colombia de estas tres familias se habían reportado 40 géneros y 192 especies (Morgado & Souto, 2016; Rodríguez et al., 2018; Savaris et al., 2016; Wendt & Ale-Rocha, 2016), por lo cual nuestros resultados, incluyendo los nuevos registros, representan el 13.6 % de la diversidad del grupo para el país. En los Llanos Orientales se habían registrado cinco géneros y 26 especies de estas familias (Díaz & Vásquez, 1993; Lobos-Aguirre, 1997; Martínez-Alava, 2007; Núñez, 1981; Plan Nacional Moscas de las Frutas, 2012; Rodríguez & Arévalo, 2015; Rodríguez et al., 2018) y en el departamento de Arauca solo existían los registros de moscas de las frutas de importancia económica con nueve especies del género Anastrepha(Martínez-Alava, 2007; Plan Nacional Moscas de las Frutas, 2012; Rodríguez et al., 2018). Las especies Anastrepha obliqua y Neosilba batesi fueron las más abundantes durante el estudio; estas son polífagas y de amplia distribución (Aluja et al., 2003; Saavedra-Díaz et al., 2017) y esto explicaría porque son las especies más abundantes en este trabajo.
Los estudios de biodiversidad y biología de la conservación son esenciales en los ecosistemas, pero todos los inventarios son incompletos y el total de especies es imposible registrarlo (Jiménez-Valverde & Hortal, 2003); según los mismos autores las curvas de acumulación de especies y rarefacción son herramientas matemáticas que permiten modelar la diversidad teórica, muy utilizadas actualmente a pesar de que la recolecta de biodiversidad está afectada por diversos factores. Las curvas de acumulación de especies construidas en este trabajo permiten indicar que los esfuerzos de muestreo son confiables porque cumplen con el requisito de mostrar una tendencia a hacerse asintótica y porque los valores de representación de la diversidad teóricos pueden considerarse adecuados (Jiménez-Valverde & Hortal, 2003), por lo cual, la diversidad presentada en este trabajo, para el método de muestreo utilizado, es representativa de la realidad. Larsen et al. (2018) muestran como los estudios cortos fallan en estudiar adecuadamente la biodiversidad, pero trabajos como el nuestro aportan datos que en conjunto con otra información ayudan a entender los ecosistemas.
Arauca vs Saravena: Los valores de riqueza y abundancia de especies fueron estadísticamente diferentes en los dos municipios. Estos parámetros presentaron valores más altos en Arauca entre septiembre y noviembre (periodo de lluvias). En Saravena la riqueza de especies presentó valores más altos de septiembre a noviembre (periodo de lluvias), mientras que la abundancia presentó valores más altos entre enero a marzo (sequía). Aspectos climáticos contrastantes en los dos sitios de muestreo podrían ser la causa de estos resultados. Arauca es sabana con una temperatura media de 30 °C y precipitaciones anuales de 1 500 mm (Instituto de Hidrología, Meteorología y Estudios Ambientales, 2019) y una estacionalidad bimodal marcada que reduce la presencia de hospederos y la sobrevivencia de las moscas. En contraste, Saravena se encuentra en el piedemonte llanero (zona de transición entre sabana y la cordillera de los Andes) y tiene una temperatura media de 25 °C y precipitación de 3 290 mm anuales y alta humedad (Instituto de Hidrología, Meteorología y Estudios Ambientales, 2019), condiciones más adecuadas para que la sobrevivencia de los tefrítidos sean los períodos de menos lluvias, permitiendo una mayor disponibilidad de hospederos. Esto se ratifica en el análisis de similitud de Jaccard que indicó que los sitios de muestreo se agrupan de acuerdo con el municipio y una mayor diversidad de especies en Saravena, quizás por representar una zona de transición entre la Sabana y los Andes. Diversos trabajos han mostrado la influencia de la precipitación en los picos poblacionales de moscas frugívoras (Aluja, 1994; Canesin & Uchôa-Fernandes, 2007; Jirón & Hedström, 1988; Oliveira et al., 2009), siendo que en regiones con climas extremos las poblaciones son mayores con condiciones más benignas; Jirón & Hedström (1988) mostraron la influencia del clima en la fenología de los hospederos y por tanto en las moscas frugívoras. Muy seguramente los tefrítidos no frugívoros recolectados en este trabajo también están influenciados por la presencia de hospederos y estos a la vez por las condiciones climáticas.
Huerto doméstico vs parche de bosque: La riqueza de especies fue mayor en parches de bosque respecto a las huertas domésticas, sin embargo, la abundancia fue mayor en los huertos domésticos. Aunque todas las especies registradas en este estudio son nativas de la región y en general de amplia distribución, cabe destacar que la mayor abundancia en huertos domésticos corresponde a especies frugívoras, mientras que en los parches de bosque las especies predominantes son las relacionadas con flora nativa como asteráceas (Norrbom, 2010). En Saravena, en la huerta doméstica la especie dominante fue A. obliqua y en el parche de bosque M. cyanogaster mientras que en Arauca en la huerta doméstica fue N. batesi y en el parche de bosque X. chrysura. E. paupera, X. chrysura, R. podagrica y S. anale, que son especies no frugívoras, se encontraron en todas las áreas muestreadas. Estudios previos han mostrado como en áreas poco intervenidas las especies oligófagas tienden a ser predominantes (Aluja et al., 2003). Nuestros resultados indican que las poblaciones de moscas de las frutas (sensu frugívoras) en las áreas muestreadas son naturalmente bajas (como se presenta en los parches de bosque) y que aumentan con la actividad antrópica, que incrementa la disponibilidad de más recursos ecológicos como frutos hospederos para su desarrollo. Es posible predecir que la influencia antrópica podría conllevar un aumento de las especies de moscas y propiciar adaptaciones a los hospederos plantados provocando la reducción de las poblaciones de otras especies. Esto es validado por Aluja (1994) y Soto y Jirón (1989) que plantearon que la frecuencia de las especies de moscas de las frutas está influenciada principalmente por factores ecológicos como la abundancia y riqueza de las especies de plantas hospederas y los huertos frutícolas proveen estas condiciones óptimas para su proliferación. Actualmente la región estudiada está poco intervenida, sin embargo, como lo sugieren estos resultados, el efecto sobre la biodiversidad de moscas puede ser significativa. Intervenciones antrópicas a gran escala, como está planteado por las políticas gubernamentales, podría acentuar los efectos sobre ciertos taxones de insectos como los registrados en este estudio. Esas transformaciones generan la necesidad de establecer mecanismos de estudio y monitoreo con miras a procurar el mantenimiento de la biodiversidad local y los servicios ecosistémicos que proveen. También sugerimos, con base en nuestros resultados, que el aumento de la actividad antrópica dirigida al sector frutícola llevará al aumento de las poblaciones de insectos plaga.
Moscas de la fruta: El resultado del índice poblacional para las especies de moscas de la fruta no presentó valores que sobrepasaran los umbrales económicos (MTD < 1), concordando con información publicada previamente en los boletines epidemiológicos del ICA (Plan Nacional Moscas de las Frutas, 2012). Los valores bajos del MTD deben estar relacionados con la poca presencia de cultivos de plantas hospederas, pues la presencia de frutos hospederos es uno de los factores ecológicos que más influye positivamente en las poblaciones de moscas frugívoras (Aluja, 1994; Soto & Jirón, 1989). Planteamos que las condiciones climáticas en los dos sitios de muestreo debieron influir en los valores del MTD; en el municipio de Arauca, de clima seco, los MTD fueron más altos en época de lluvia (agosto - noviembre), ya en Saravena, de clima muy húmedo, fueron mayores en época de bajas lluvias (diciembre - febrero). Sin embargo, el hecho que las mayores poblaciones se encuentren en áreas de intervención antrópica sugiere un efecto favorable para estos insectos, si a futuro se amplían las áreas de producción frutícola en estos ecosistemas.
En la recolecta de frutos las especies de la familia Lonchaeidae fueron las que presentaron asociación con los hospederos. En este estudio encontramos que N. batesi fue la especie más polífaga y se asoció con cinco hospederos: guanábana (Annona muricata), aguacate (Persea americana), guayaba (Psidium guajava), naranjas (Citrus sinensis) y limoncillo (Swinglea glutinosa). Mientras que A. striata fue la única especie de la familia Tephritidae que se obtuvo a partir de frutos y se asoció a Psidium guajava.
También se observó a N. batesi, N. zadolicha y L. longicornis compartiendo el hospedero Annona muricata (guanábana) como previamente reportado por Saavedra-Díaz et al. (2017), y en Rollinia mucosa (Anón Amazónico) se encontraron en un mismo fruto tres especies de Neosilba cohabitando. En frutos de Psidium guajava se encontraron especímenes de A. striata y N. batesi de la misma recolecta, sin embargo, no pudimos observar si se encontraban compartiendo el mismo fruto. Se ha documentado que especies de las familias Tephritidae y Lonchaeidae compartan el mismo hospedero sin afectar su desarrollo, pero las especies de Anastrepha en el bosque seco tropical no comparten hospederos (Bomfin et al., 2014; Nicácio & Uchôa, 2011; Ruiz-Hurtado et al., 2013; Saavedra-Díaz et al., 2017). Castañeda et al. (2010) planteó que las especies polífagas, las de mayor importancia económica, usan diferentes hospederos, algunos incluso compartidos con otras especies polífagas, como una adaptación y estrategia ecológica para mantener sus poblaciones en épocas de poca presencia de frutos.
La información acerca de índices de infestación por especies de la familia Lonchaeidae en Colombia es escasa. Santamaría et al. (2014) mostraron la incidencia de larvas de Dasiops spp. en distintos órganos de pasifloras con un máximo de dos larvas/órgano. Saavedra-Díaz et al. (2017) mostraron índices de infestación bajos en muestras de frutos recolectadas del árbol con un máximo de 2.06 larvas/fruto y 123.38 larvas/kg fruto, mientras que en muestras recolectadas de suelo fue de 8.03 larvas/fruto y 68.63 larvas/kg fruto. En el presente estudio se obtuvieron índices de infestación aún más bajos (< 0.1 larvas/kg fruto) en ambos municipios, a excepción de octubre en Arauca en frutos de guanábana (142.5 larvas/kg de fruto). Este último dato, sin embargo, corresponde a un único fruto que estaba disponible, del que se obtuvieron 109 moscas adultas. De igual manera, las bayas de Miconias son pequeñas y a pesar del número recolectado (355) apenas pesaron 151 g, resultando en un índice de infestación más alto que en los otros hospederos. Estos resultados indican que, aunque los individuos de la familia Lonchaeidae se encuentran presentes en los frutos, su presencia no es tan dañina como se da en otros países (Aguiar-Menezes et al., 2004; Araújo & Zucchi, 2002; Strikis et al., 2011; Uchôa-Fernandes et al., 2003). Sin embargo, es necesario ampliar los estudios de esta familia no sólo en la región de los Llanos Orientales sino también en el país.
En conclusión, este trabajo aporta información novedosa sobre la biodiversidad de Tephritoidea en la provincia biogeográfica de Sabana y sobre el comportamiento de las moscas de la fruta (Tephritidae) en la Orinoquía colombiana, la cual es ampliamente desconocida. Se documenta la diversidad local con nuevos registros para el país y aspectos de su historia natural para una región con mucho potencial frutícola según las agendas del Estado colombiano.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

