











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
As the world enters the third decade of the XXIth century there is growing concern for the increasing impacts of global stressors such as costal alteration, pollution (Delorenzo, 2016) eutrophication (Cloern, 2001), and climate change (Ni et al., 2019). These impacts become more noticeable at certain coastal areas such as estuarine ecosystems. Most of the information on coastal ecosystems and their environmental dynamics comes from studies conducted in the temperate regions. In spite of this intensive research effort, a great deal of questions formulated years ago to orient coastal research remains yet to be answered (Nordstrom & Roman, 1996). In this context, a better understanding of the impact of global stressors on these ecosystems might be reached if research which focuses on comparative studies in temperate and tropical sites is emphasized. Comparative research may also help to better identify the spatial and temporal variability of these stressors and the feasibility of management measures.
Golfo Dulce is a unique deep embayment on the Pacific coast of Costa Rica and a cooperative multidisciplinary research effort involving local and foreign scientists was started 25 years ago with the R/V Victor Hensen expedition. The expedition found interesting similarities between Golfo Dulce and temperate fjords and prompted it to be called a fjord-like embayment. The expedition produced pioneering biogeochemistry information and data on the distributions of its benthic and planktonic biota (Wolff & Vargas, 1994; Vargas & Wolff, 1996). This data base, complemented with follow-up studies, remains as the main reference for many faunal groups and environmental parameters. However, it needs to be updated and new research areas approached emphasizing comparative studies with similar environments worldwide. A first step towards reaching this goal is to summarize the published literature and suggest priorities for further research. Thus, the objectives of this review are to introduce the reader to published research conducted in Golfo Dulce, identify topics that need to be explored further, and propose new issues to be addressed in future surveys.
General description
Golfo Dulce (Fig. 1) is a gulf located on the Southern Pacific coast of Costa Rica (8o30’N & 83o15’W). It has an area of about 750 km2, a length of 50 km, and maximum widths between 10 to 20 km. The coastline extends for 195 km (Morales-Ramírez, 2011). Tidal amplitude is around 3 m. In marked contrast to the Gulf of Nicoya, a tectonic estuary also on the Pacific coast of Costa Rica (Vargas, 2016), the bathymetry of Golfo Dulce is characterized (Hebbeln et al.,1996) by a deep (maximum depth 210 m) basin in the upper region, a sill (60 m) at the entrance, and a slope that deepens to more than 200 m at the mouth (Fig. 1). In Golfo Dulce a gradient from oxic to hypoxic, and sometimes anoxic conditions, occurs.
This deep bathymetry and the presence of a sill at the entrance makes Golfo Dulce unique on the West coast of the American continent and also makes it resemble a temperate fjord. Also, in contrast to the Gulf of Nicoya, there are no islands. Golfo Dulce is also a tectonic gulf and the inner section is fringed by two parallel faults along its Northern and Southern shores (Malzer & Fiebig, 2008). Narrow shorelines that fall almost vertically down to basin depths (Fig. 1) characterize this inner section (Hebbeln et al.,1996). The Southern and Eastern shores of Golfo Dulce include sandy beaches and the latter has an extensive shallow subtidal platform (Fig. 1). The tip of the Osa Peninsula (Fig. 1) has a mixture of rocky shores and sandy beaches.
Most of the Peninsula includes the Corcovado National Park (424 km2) with extensive rain forests. The Northern and Western shores also include rain forests which are protected by the Golfo Dulce Forest Reserve (620 km2), the Piedras Blancas National Park (140 km2), and the Golfito Wildlife Refuge (13 km2). The two main towns on the shores are the port of Golfito on the Northern shore, and the port of Jiménez on the Northern tip of the Osa Peninsula, with populations (2011) of 12 000 and 9 000, respectively.
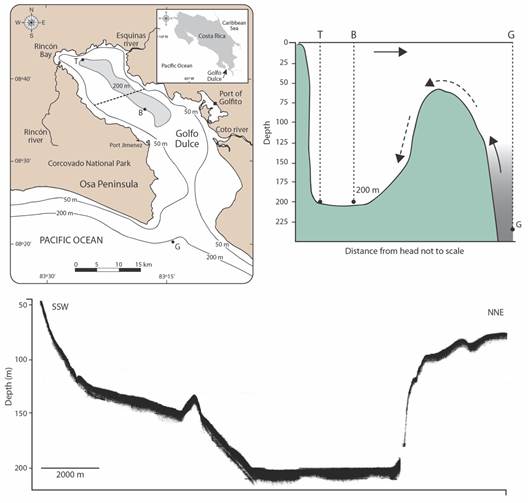
Fig. 1 Left: Map of Golfo Dulce, Pacific coast of Costa Rica. Stations B (basin edge), T (Rincón Bay), and G (Shelf edge) are indicated. Light gray in the upper gulf encloses 200 m depth area. Dashed line across indicates R/V Victor Hensen echograph transect. Right: Schematic depth profile along the central axis of Golfo Dulce. Arrows indicate water circulation. Broken arrows represent occasional influx of shelf waters over the sill. Light gray represents deep Equatorial Subsurface Waters. Bottom: R/V Victor Hensen echograph of a bottom topography transect across mid upper Golfo Dulce. Note that the ecograph starts at the 50 m depth contour. The northern fault is evidenced by the steep wall at the right of the figure. (Adapted from Hebbeln et al., 1996).
Rivers, mangrove forests and seagrass beds
The Rincón River on the West, the Coto River on the East, and the Esquinas River on the North shore are the main rivers discharging into the embayment (Fig. 1). Information on the latter two rivers has been provided by Angulo et al. (2020), Michels et al. (2006) and Tschelaut et al. (2008). In addition, several small creeks reach the shores of Golfo Dulce (Umaña, 1998). River flow varies seasonally and ranges from 20 to 180 m3/sec (Svendsen et al., 2006). Mangrove forests are found near the mouths of these rivers and are dominated by Rhizophora racemosa, R. mangle, Pelliciera rhisophoreae, Laguncularia racemosa and Avicennia germinans, among others. Species composition and dominance at these mangroves is similar to that reported for the region (Samper-Villarreal & Silva-Benavides, 2015). Recent research on the mangroves of Golfo Dulce has focused on isotopic evidence of nutrient enrichment at nutrient loaded sites from mangrove material. The macroalgae Bostrychia calliptera was found growing on R. mangle prop roots (Samper-Villareal et al., 2018). The mangroves of Playa Blanca on the Southern shore and those of two small estuaries in the inner Golfito Bay include populations of the ark clam Anadara tuberculosa, which sustain a small fishery (Silva-Benavides & Bonilla, 2001; Silva-Benavides & Bonilla, 2015). Stern-Pirlot and Wolff (2006) have made recommendations for the management of these A. tuberculosa populations. Information on physical and chemical parameters of the small estuaries at Golfito Bay is provided by Silva and Acuña-González (2006). The amino acid and protein content of the oyster Crassostrea gigas cultured at Rincón Bay has been reported by Fonseca-Rodríguez and Agüero-Pérez (2017).
Organisms recently recorded for Golfo Dulce are the seagrasses Halophila baillonii and Halodule beaudettei. On the Southern shore seagrass meadows are located at Colibri beach East of Rincón Bay and near Puerto Jiménez. On the Northern shore the beds are present at Golfito Bay and at the beach of the wildlife refuge. Meadow extension ranged from about 100 m2 at the refuge and Golfito to 900 m2 at Puerto Jiménez and 900 000 m2 at Colibri beach. Seagrasses in Golfo Dulce are found within a narrow coastal belt at water depths of 3 to 6 m in sediments containing less than 6.5 % of silt+clay. The sediments of Colibri included 26 species of polychaete worms. Sea turtles are known to feed on seagrass beds in Golfo Dulce (Samper-Villarreal et al., 2014; Samper-Villarreal & Cortés, 2020).
Coral reefs
Fringing, low diversity coral reefs are found in the Northern shores of Golfo Dulce and include the coral Porites lobata and several other species. Sedimentation has degraded these reefs considerably. Environmental conditions promoted coral reef growth in the past but are deteriorating mainly due to siltation (Cortés, 1990; Cortés, 1992; Cortés et al., 2010; Cortés, 2016). However, a Psammocora stellata reef increased its coral coverage from 46 to 83 % since 1990 (Alvarado et al., 2015). More healthy patches of coral are found in the outer gulf and are characterized by a higher richness of species. In the destruction of the reefs is also important the impact of biological eroders, such as sponges (Cliona sp), bivalves (Lithophaga spp), sipunculans (Aspidosiphon elegans), crustaceans (Upogebia sp), and others (Fonseca et al., 2006). A recent “coral gardening” experiment was conducted by Vargas-Ugalde et al. (2020) in Golfo Dulce to promote ecological restoration of damaged reefs. A new species of soft coral (Leptogorgia cortezi) was found by Breedy and Guzmán (2012) at Punta Islotes and Punta Estrella on the Northern shore reefs. A new species of cirratulid polychaete worm (Aphelochaeta zebra) has been described recently also from Punta Islotes reefs by Dean and Blake (2016).
Temperature, oxygen and salinity profiles
The Golfo Dulce region has less defined dry-rainy season periods than the Gulf of Nicoya (Cortés, 2016). Rainfall is common during the whole year and may range from 100 mm to more than 600 mm/month. The drier months are December through March, when rainfall may be around 100 mm per month while peak rainfall occurs in October (Svendsen et al., 2006). Due to the enclosed nature of the upper region which is protected from northerly winds by forest covered mountains the waters of the inner gulf are relatively calm most of the time. An early survey of water parameters was conducted in March 1969 during a cruise of the R/V Thomas Thompson (Richards et al., 1971). The results indicated that the embayment was characterized by surface salinities of 31 to 33 psu and temperatures of up to 32.2 oC at the surface and as low as 14.3 oC at 200 m depth. The results of the cruise also provided evidence for the hypothesis of the renewal of basin water with offshore water flowing over the sill (Fig. 1).
The deep bathymetry of Golfo Dulce exerts an important control on the distribution of oxygen and on the biota of the ecosystem. The surface waters are well oxygenated most of the time and oxygen concentrations decrease rapidly with depth to hypoxic or even anoxic conditions with occasional formation of hydrogen sulphide near the bottom. The hydrogen sulphide is apparently removed by oceanic water occasionally flowing across the sill and is difficult to detect during sporadic visits (Córdoba & Vargas, 1996). The survey conducted in March 1969 by the R/V Thomas Thompson detected oxygen concentrations of less than 10 % below 30 to 40 m depths and hydrogen sulphide was found a few meters above the sediments of the basin. Bottom salinities were near 35 psu (Richards et al., 1971). A study of physical and chemical parameters was conducted by Brenes-Rodríguez and León-Coto (1988). They considered Golfo Dulce as a shallow sill estuary that could be divided into two main layers separated by the 19 oC isotherm. A strong thermohalocline makes mixing difficult between layers. The study also identified denitrification and nitrate reduction processes as well as organic matter decomposition.
Another survey was conducted in December 1993 aboard the R/V Victor Hensen (Wolff & Vargas, 1994; Vargas & Wolff, 1996). At station G (200 m, Fig. 1) located at the entrance to Golfo Dulce, oxygen concentrations (electronic sensor, % saturation) were near 50 % at the surface, increased to 60 % at around 10 m, and dropped to 30 % at 50 m. Concentrations then dropped sharply with depth and were near 2 % at 200 m. Water temperature was 13 oC at 200 m. These conditions are similar to those found in offshore waters along the Pacific coast of Costa Rica. At station B (200 m, Fig. 1) located at the basin edge, the concentration of oxygen at the surface was near 40 %, increased to 60 % at 10 m below the surface, and then decreased to near 2 % at depths of more than 150 m. Temperature was 13 oC at 200 m. Another profile was obtained by Córdoba and Vargas (1996) in January 1994 at station T (200 m, Fig. 1) located in the North-West margin of the upper Golfo Dulce. At this station temperature ranged from 31.5 oC at the surface to 17 oC at 200 m. A thermocline was evident between 20 and 50 m. Salinity at the surface was 29.5 psu and close to 35.0 psu at the bottom. Dissolved oxygen concentrations (Winkler method, micromoles/litre) varied between 425 µmol/l at 10 m to non-detectable at 200 m. Nitrate was also non-detectable at 200 m, but it was found several meters above the bottom. Another survey by Acuña-González et al. (2006b) at Station T gathered data during occasional visits between June 1993 and September 1995 (Fig. 2).
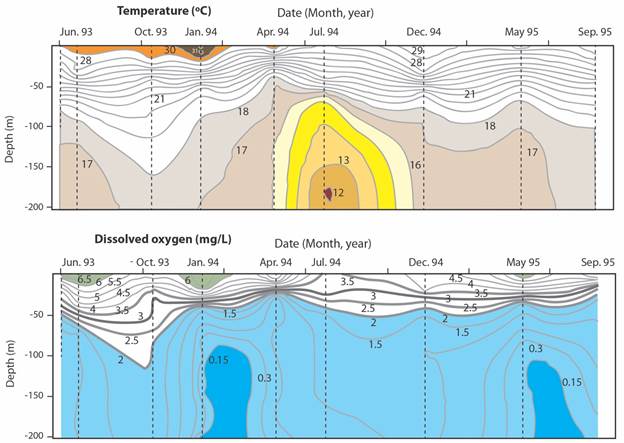
Fig. 2 Occasional sampling (June 1993 to September 1995) at depth intervals from surface to 200 m depth at station T (Upper Golfo Dulce, near Rincón Bay, Fig. 1). Top. Temperature (oC) oscillations. Bottom. Oxygen (mg/l) oscillations. Oxygen concentrations of less than 2 mg/l are shaded in blue. Adapted from Acuña-González et al. (2006b).
The survey at station T (Fig. 2) illustrated temporal oscillations of parameters at a given depth: The highest surface temperature observed (31 oC) was recorded on January 1994. The minimum temperature (12 oC) was observed on July 1994 at a depth of more than 180 m. Maximum oxygen concentrations (milligrams/litre) near the surface were 6.5 mg/l and observed in June 1993, January 1994, and May 1995. The lowest oxygen concentrations (0.15 mg/l) were seen at depths of more than 100 m during visits on February 1994 and June 1995 and lasted for short periods. Anoxic conditions were not found. Hypoxia is defined as oxygen concentrations lower than 2.8 mg O2/l (Wu, 2002). Thus, hypoxic conditions characterize Golfo Dulce at depths or more than 30 m (Fig. 2).
A comprehensive study of water parameters along Golfo Dulce was conducted between 2005 and 2007 by Morales-Ramírez et al. (2015) and included an evaluation of rainfall, currents, and nutrient concentrations. Rainfall ranged from 100 to near 700 mm/month. The study found the presence of low speed currents (range 5.1 cm/s in the inner gulf to 17.8 cm/s at the mouth). At a station on the Western end of Golfo Dulce near Rincón Bay (Fig. 1) several surface water parameters were recorded. A maximum Secchi disk depth of 10 m was measured. At a depth interval of 70 to 140 m the minimum temperature was 16.6 oC and oxygen concentration was 0.5 mg/l. The maximum temperature found at this depth range was 17.1 oC and oxygen concentration was 0.95 mg/l. Concentrations of nitrate (16.6 µmol/l) and phosphate (2.7 µmol/l) at this depth range are similar to those reported for equatorial subsurface waters (23 and 2.0 µmol/l, respectively) at the entrance to the Gulf of Nicoya by Chaves and Birkicht (1996). These studies, as well as previous ones, illustrate the temporal variability of parameters in Golfo Dulce and provide a unique framework for future comparative studies focusing on describing the seasonal and depth fluctuations of environmental parameters.
Another survey by Rincón-Alejos and Ballesteros-Sakson (2015) collected data during the dry and rainy seasons of 2009-2010 using a CTD (Conductivity, Temperature, Depth) recorder at 33 stations in the upper gulf. The study also measured Secchi disk depth. In marked contrast to the upper Gulf of Nicoya that has maximum Secchi disk depth of 2 m (Vargas-Zamora et al., 2018) the disk was visible in Golfo Dulce as deep as 21.5 m. A wide surface salinity range was recorded during the rainy season (18.0 to 31.4 psu). Temperature at the surface ranged from 27.9 to 32.0 oC.
Oxygen stratification, H2S formation and the anammox reaction
During the R/V Victor Hensen expedition (Wolff & Vargas, 1994; Vargas & Wolff, 1996) part of the time was devoted to biogeochemistry studies. A transect from shelf waters to over the sill and into the basin revealed that Golfo Dulce had decreasing concentrations of dissolved oxygen and nitrate at similar depths when compared to those found offshore (Fig. 3).
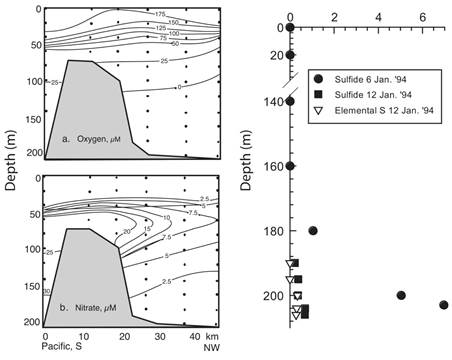
Fig. 3 Left: Golfo Dulce. R/V Victor Hensen, January 1994. Vertical profiles of dissolved oxygen and nitrate along a 40 km transect (Shelf edge - sill - basin). Note that no oxygen was found below 100 m in the basin and was near 25 µmol/l in shelf waters at the same depth. Nitrate followed a similar pattern with values of 2.5 µmol/l at 150 m in the basin and 30 µmol/l in the shelf at the same depth. Right: Dissolved sulphide and suspended elemental sulfur near station T, upper Golfo Dulce. Note change of sulphide at 200 m from 7 µmol/l to less than 1 µmol/l after 6 days. From Ferdelman et al. (2006).
As a result, the water column of the inner region of Golfo Dulce was described as divided into four depth layers: the oxic layer between the surface and the thermocline; the interval between 40 to 100 m characterized by a steep oxygen gradient; a third layer from about 100 to 200 m, which is sub-oxic but nitrate is present and a fourth interval several meters above the bottom sediments, with occasional nitrate depletion and sulphide accumulation (Thamdrup et al., 1996). Dissolved sulphide concentration measured near station T (Fig. 1) on January 6th, 1994 (Fig. 3) increased from 1 µmol/l at 180 m to 7 µmol/l at 203 m, and decreased to around 1 µmol/l after six days (Ferdelman et al., 2006). H2S disappeared coinciding with the presence of nitrate found during another simultaneous study (Córdoba & Vargas, 1996). Rates of oxygen consumption were higher in the surface waters (up to 60 µmol/l/day), decreased to below 10 µmol/ l /day in the pycnocline and was non-detectable deeper than 60 m (Thamdrup et al., 1996; Ferdelman et al., 2006). Sulphide oxidizing bacteria of the genera Thiospira and Thiovulum, as well as the filamentous bacterium Beggiatoa were identified in the sediments (Kuever et al., 1996; Thamdrup et al., 1996). Years later, Gallardo and Espinoza (2007) identified large multicellular filamentous bacteria (Beggiatoa spp, Thioploca spp), as well as spherical Thiomargarita-like forms in slightly sulphidic core samples taken at 195 m depth in Golfo Dulce. These morphologically diverse bacterial communities were also found in organically rich sediments below the oxygen minimum zone in coastal areas from central Chile to Costa Rica.
An important line of research in Golfo Dulce focused on the interactions at the water-sediment interface of the basin. It was proposed five decades ago that basin waters are renewed on a more or less regular basis by coastal waters flowing over the sill and removing sulphide (Richards et al., 1971). Sulphide concentrations of less than 3 µmol/l were found above 15 cm of the sediment and decreased over a few days. Organic matter debris degraded through bacterial sulphate reduction may serve as patchy sources of sulphide (Stief et al., 2017). Other types of micro-organisms studied in Golfo Dulce are anaerobic ammonium oxidizing (Anammox) bacteria that play a key role in the marine nitrogen cycle through the anammox reaction. Anammox activity has been reported in the sediment and water column of Golfo Dulce where it accounted for up to 35 % of the total N2 formation in the water column. This process makes Golfo Dulce similar to other low oxygen zones worldwide. The anammox activity quantified in sediments ranged from 0.08 f mol/cell/day N2 in Golfo Dulce, to 0.98 f mol/cell/day N2 in the Gullmar fjord, Sweden (Schmid et al., 2007). Other studies have focused on the bacteria and related topics (Canfield, 1998; Dalsgaard et al., 2003; Dalsgaard et al., 2005; Rush et al., 2014; Stief et al., 2017).
The finding of the anammox reaction in the deep waters of Golfo Dulce was an important discovery facilitated by joint international cooperation (Dalsgaard et al., 2003). This reaction makes Golfo Dulce a nitrogen sink (Acuña-González et al., 2006b). The anammox reaction occurs in other ecosystems such as the Black Sea (Dalsgaard et al., 2006) and comparisons of the reaction rates have been done with those found in Golfo Dulce (Jensen et al., 2008). More recently, studies on the diversity, metabolic potential and activity of Marinimicrobia bacteria, microbial enzymes, and other microbiological aspects have been conducted in Golfo Dulce (García-Robledo et al., 2016; Padilla et al., 2016; Bertagnolli et al., 2017; Bertagnolli et al., 2020). The study by Padilla et al. (2017) provided first field evidence for methanotrophic partial denitrification under anoxic conditions. However, more research would certainly help to achieve a better understanding of these processes.
Oxygen producers and primary production
Unlike the Gulf of Nicoya estuary (Vargas, 2016) there is no information on oxygen producing phytoplankton species in Golfo Dulce or primary productivity estimates based on the release of oxygen by microalgae. The study by Wolff et al. (1996) mentions that the phytoplankton of Golfo Dulce appears dominated by diatoms and also by dinoflagellates that include red tide species.
However, primary productivity data is available from a station at the channel entrance to a small secondary shallow bay on the Northern shores of Golfo Dulce where the port city of Golfito is located (Fig. 1). At this site, net primary productivity ranged from 695 to 1 371 gC/m2/year. Concentrations of chlorophyll-a determined at this site ranged from 0.8 to 2.1 mg/m3, and dissolved oxygen was above saturation (Vargas-Zamora et al., 2018). An unpublished report by Lei (2002), based on remote sensing techniques, estimated phytoplankton primary production (PPC) of Golfo Dulce in the range of 138 to 1 007 gC/m2/year. PPC was found higher during the rainy season. As a comparison, net primary productivity estimates for the upper Gulf of Nicoya estuary ranged from 191 to 1 601 gC/m2/year (Vargas-Zamora et al., 2018).
The study by Rincón-Alejos and Ballesteros-Sakson (2015) provided data on surface chlorophyll concentrations at 22 stations along Golfo Dulce over a period of one year (Fig. 4). A maximum chlorophyll-a concentration of 91.67 µmol/l was reported during the dry season (January, 2009) at the entrance to Golfito Bay while a minimum concentration of 1.34 µmol/l was found near Rincon Bay. In marked contrast to the results found in January 2009, the maximum chlorophyll-a concentrations found in August 2009 and January 2010 were 5.2 and 1.2 µmol/l, respectively (Fig. 4).
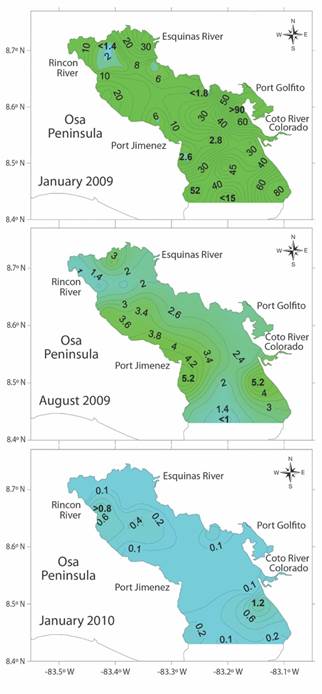
Fig. 4 Chlorophyll-a concentrations (µmol/l) at the surface of 22 stations along Golfo Dulce. January and August 2009 and January 2010. Adapted from Rincón-Alejos and Ballesteros-Sakson (2015).
The R/V Victor Hensen faunal survey and other studies
Morales-Ramírez (2011) estimated 1 022 reported species of organisms (from bacteria to mammals) for Golfo Dulce. This richness represented close to 22 % of the total number of species reported for the Pacific coast of Costa Rica. This percentage needs to be updated based on the information made available after 2010.
A significant advance on the knowledge of the biodiversity of Golfo Dulce was obtained during the R/V Victor Hensen survey (Wolff & Vargas, 1994). Plankton and trawl nets were used to study the zooplankton and demersal crustaceans. A total of 33 species of pelagic copepods were found, including Paracalanus aculeatus, Temoropia mayumbaensis, Lucicutia ovalis, Candacia catula and Corycaeus speciosus (Morales-Ramírez, 1996). The abundance, biomass, and vertical distribution of other micro-crustaceans, such as ostracods were studied by Morales-Ramírez and Jakob (2008). Four species of ostracods were identified belonging to the genera: Cypridina, Euconchoecia and Porroecia. Their abundance appears high when compared to offshore waters. Another survey (Morales-Ramírez et al., 2015) reported dominance (44 to 84 %) of copepods such as Oncaea mediterranea, Oithona plumifera, Acrocalanus gibber, Paracalanus parvus, P. aculeatus and Centropages furcatus. The gelatinous zooplankton was also studied and 41 species were identified of which 33 were cnidarians, six were thalaceans and two were ctenophores (Morales-Ramírez & Nowaczyk, 2006). The biomass spectra and species composition of the zooplankton communities was studied by von Wangelin and Wolff (1996). They found that at the inshore and offshore stations high amounts of gelatinous zooplankton (Siphonophores and salps) dominated the largest size groups, and that ostracods and brachyuran zoea appear as main crustacean components inshore during the dry season.
Other group of organisms found during the expedition were the arrow worms or chaetognaths (Hossfeld, 1996) with 10 species recorded for Golfo Dulce of which nine were of the genus Sagitta and one of the genus Krohnitta. The ichthyoplankton was studied by Molina-Ureña (1996) who reported larvae of the families Bregmacerotidae, Myctophidae, Engraulidae and Ophichthidae as the more abundant among the 15 families identified for Golfo Dulce.
The decapod and stomatopod crustacean fauna were also evaluated in Golfo Dulce with trawl nets during the R/V Victor Hensen survey and the results summarized by Castro and Vargas (1996), Jesse (1996), and Vargas et al. (1996). No crustaceans were found in the deeper low-oxygen waters. Golfo Dulce was found to be a relatively species poor area, with smaller biomass and diversity, when compared to other sites on the Pacific coast of Costa Rica such as the Gulf of Nicoya. The recently updated list of decapod and stomatopod crustaceans collected during the R/V Victor Hensen and those hand-collected intertidally and by diving during other surveys includes 106 species, of which four were stomatopods and 102 were decapods belonging to 32 families (Vargas-Castillo & Vargas-Zamora, 2020). More recently Salas-Moya et al. (2021) listed the crustaceans associated with the soft corals Eugorgia mutabilis and Leptogorgia cortezi, as well as to the sea star Pentaceraster cunningi and onuphid polychaete worms.
Fish diversity and fisheries
A trawl survey of the fish present in Golfo Dulce was conducted as part of the R/V Victor Hensen expedition and a total of 75 species were collected there (Bussing & López, 1996). The fish fauna was dominated by small species of the genera Cynoscion and Porichthys, which occurred in low densities, perhaps due to a low carrying capacity of the embayment (Wolff, 1996). An updated list of pleuronectiform flatfish species collected in Golfo Dulce by the R/V Victor Hensen was assembled by Vargas et al. (2020) and included 19 species of which Symphurus leei was the most frequent. Other studies emphasized the critical role of Golfo Dulce as a habitat and nursery area for the shark Sphyrna lewini (Zanella & López-Garro, 2015; Zanella et al., 2019). Feeding aggregations of the whale shark Rhincodon typus were reported for the inner Golfo Dulce by Pacheco-Polanco (2015). Hanna (2016) reported 226 sightings of R. typus for the period 2001-2007 which included several juveniles (2-3.5 m long).
The fish fauna on the Eastern shore of Golfo Dulce has been recently studied at the mangrove swamp of Zancudo at the mouth of the Coto River (Feutry et al., 2010). A total of 1 591 individuals belonging to 82 species in 30 families were found at Zancudo. Sciaenidae was the most diverse family (8 spp) during the rainy season and Carangidae (10 spp) was so during the dry season. Species collected more frequently were: Anchoa mundeola, Centropomus armatus, Diapterus peruvianus and Mugil curema. The fish fauna of an intertidal rocky islet in Golfo Dulce was studied by Palacios-Martínez (2015). He found a total of 28 species of which the more abundant were Thalassoma lucasanum (Labridae), and the pomacentrids Chromis atrilobata and Abudefduf trocshelii.
Studies on the fisheries of Golfo Dulce were initiated by Segura and Campos (1990) who estimated in 5.5 % the post-harvest losses at landing sites. A description of the artisanal fishery was conducted by Guzmán-Mora and Molina-Ureña (2007) who found that small boats (4-10 m long) comprised the main fleet of fishermen who employed hook and line, long line and trammel nets. Four species (the snappers, Lutjanus guttatus and L. peru, the mackerel, Scomberomorus sierra, and the grunt Pomadasys panamensis) accounted for 66.5 % of the captures. The study by Guzmán-Mora and Molina-Ureña (2007) concluded that the fisheries in Golfo Dulce are mainly artisanal, based at the port of Golfito, and are less developed than those in the Gulf of Nicoya, and only four species account for up to 67 % of the captures. López-Garro and Zanella (2015a) evaluated onboard operations of the artisanal fishing fleet in Golfo Dulce and found that from 872 fish observed, 39.6 % were sharks, 26.1 % were bait fish (Ariidae), 13.1 % were rays and 12.7 % were commercial fish of the families Lutjanidae, Sciaenidae and Serranidae. They also analysed 609 sharks representing 10 species of which Sphyrna lewini accounted for 51.8 % followed by Mustelus lunulatus (23.5 %). Most of S. lewini were juveniles. Captures of the ray Dasyatis longa were studied by López-Garro and Zanella (2015b) who found that among 112 rays captured 75.7 % were of this species. The establishment of marine areas of responsible fishing and co-management of fisheries has been evaluated for Golfo Dulce by Fargier et al. (2014).
Reptiles and cetaceans
There is an important base of information on air breathing vertebrates that live on this ecosystem. There are reports on the population of the sea snake Pelamis platurus (Bessesen, 2012) and its colour variability (Solórzano, 2011), and on a recently described subspecies (Bessesen & Galbreath, 2017). The population and habitat use of the green turtle Chelonia mydas has been described by Chacón-Chaverri et al. (2015a), and Bessesen and Saborío (2012). The trophic ecology of the hawksbill sea turtle Eretmochelys imbricata was studied by Chacón-Chaverri et al. (2015b) and by Méndez-Salgado et al. (2020).
Research on marine mammals has been emphasized with studies on humpback whales (Megaptera novaeangliae) by Bessesen (2015) and Herra-Miranda et al. (2016), the false killer whale Pseudorca crassidens (Sánchez-Robledo et al., 2020), dolphins (mainly the bottlenose dolphin Tursiops truncatus and the spotted dolphin Stenella attenuata) and other cetaceans (Acevedo, 1995; Acevedo-Gutiérrez, 1996; Acevedo & Burkhart,1998; Cubero-Pardo, 1998; Cubero-Pardo, 2007a; Cubero-Pardo, 2007b; Oviedo, 2007; Herra-Miranda et al., 2016; Moreno & Acevedo-Gutiérrez, 2016; Oviedo et al., 2018). Based on 943 sightings over a period of nine years the study by Oviedo et al. (2015) identified 12 species of cetaceans in Golfo Dulce. A gradient of cetacean diversity was observed from the inner Golfo to waters near the shelf region. Bottlenose and spotted dolphins dominated the inner area while bottlenose dolphins and migratory humpback whales dominated the sill area.
Oviedo-Correa et al. (2009) evaluated the potential impact of the proposed construction of tuna culture farms on the populations of cetaceans of Golfo Dulce. The presence of lacaziosis-like disease photographed in T. truncatus has been reported by Bessesen et al. (2014).
Sedimentary environments
The sedimentary environments of Golfo Dulce are divided into two groups: those exposed to oxic conditions to a depth of around 30 m, and those exposed to hypoxic or anoxic conditions at deeper waters. Sediments at the inner basin consist mostly of turbidites, while other areas include a wide range of soft sediments, mainly silts and clays (Hebbeln & Cortés, 2001). Black and very soft sediments were reported in the inner basin (200 m), while brown and soft ones were found near Rincón Bay (50 m). Green and hard sediments were found at 75 m near the sill (León-Morales & Vargas, 1998). The first evaluation of the benthic communities was conducted in 1969 aboard the R/V Thomas Thompson using a van Veen grab at depths from 64 to 200 m in the inner basin. The abundance and biomass of invertebrates were found to be less than those expected for a tropical environment. Most of the fauna was found at depths around 70 m and only a few to zero individuals at depths greater than 100 m. The fauna included 25 species of polychaete worms of which, Paraonis lyra (Paraonidae) was the most abundant (Nichols-Driscoll, 1976).
Another evaluation took place in 1993 during the expedition of the R/V Victor Hensen. A box corer was used to sample the bottom sediments at depths from 43 to 200 m (Wolff & Vargas, 1994) and the results were summarized by León-Morales and Vargas (1998). Higher numbers of individuals were found at depths from 43 to 100 m in the upper region. The station located near the sill (75 m) was the most diverse (37 species), followed by those at the mouth (34 and 29 species). A 50 m deep station near Rincón Bay yielded 25 species. Sediments deeper than 100 m were azoic. The fauna was dominated by polychaetes (47 species), but faunal composition was drastically different from that reported by Nichols-Driscoll (1976). The polychaete P. lyra was not found. The most abundant of the 27 species of polychaetes were members of the genera Prionospio and Aricidea, which accounted for 19.5 and 13.3 % of the total number of 1 608 individuals of all groups collected (Dean, 1996; León-Morales & Vargas, 1998). Other groups of benthic invertebrates collected during the R/V Victor Hensen were the mollusks (Cruz, 1996). The most abundant gastropods in Golfo Dulce were Polystira oxitropis and Strombina fusinoidea, while Tellina ecuadoriana was a frequent bivalve. A later inventory of intertidal and shallow-water mollusks listed 252 species of gastropods, mainly prosobranchs and pyramidelids (Hoisaeter, 1998). Information on other intertidal soft-bottom fauna of Golfo Dulce is scarce and work focused on sandy beaches at the entrance to Golfo Dulce (Corrales-Ugalde & Sibaja-Cordero, 2015). The upper intertidal was dominated by the isopod Cirolana salvadoriensis and the low intertidal by polychaete worms of the families Nereididae and Pisionidae. The intertidal sedimentary environments of the Bay of Golfito yielded three new species of cirratulid polychaete worms (Caulleriella dulcencis, C. minuta and C. murilloi) described by Dean and Blake (2007).
Models
Information on Golfo Dulce available up to 1994 was used as input for the construction of a steady-state trophic ecological model. To facilitate comparisons the Golfo Dulce model (Wolff et al., 1996) used the same analytical approach applied later in the Gulf of Nicoya model (Wolff et al.,1998). The model estimated a Total System Throughput (sum of all flows) of 1 404 g/m2/year, which is low when compared to other coastal ecosystems, such as the Gulf of Nicoya estuary (3 049 g/m2/year) or the Caeté estuary (10 559 g/m2/year) in Brazil (Wolff et al.,1998; Wolff, 2006). The results indicate that the ecosystem functions in a different manner when compared to the Gulf of Nicoya model and other tropical systems for several reasons. The most important is that Golfo Dulce is dominated by biomass and energy flow produced in the pelagic zone, thus resembling more an open ocean system than an estuarine one. Second, benthic pelagic coupling is minor as a result of its deep bottom topography. The model also predicted that, due to its reduced benthic biomass and inferred low primary productivity, an increase in the pelagic fishing pressure would negatively affect resident populations of predators, such as fish and mammals. However, these projections have not been tested yet. The trophic model needs to be improved with quantitative information on the biomass of the phytoplankton and zooplankton fractions, the amount of detritus entering the system, biomass of pelagic and demersal fish, crustacean biomass, and estimates of primary productivity and of other imports and exports. The relative importance of data on fish populations is crucial since the model used 20 compartments of which 12 correspond to fish groups.
A second modelling approach focused on the circulation of water masses in Golfo Dulce. A bidimensional non-linear numerical model was developed by Quirós (2003). He concluded that the bottom topography of Golfo Dulce allows it to be considered a tropical fjord. However, the embayment is wider than other fjords and allows the existence of temporal current swirls in the interior region. The tidal wave is the main force and the role of freshwater input (mainly of the Coto River) is relatively minor.
Another physical oceanography modelling effort in Golfo Dulce dealt with further understanding its water circulation patterns (Svendsen et al., 2006). The study conducted between 1999 and 2001 combined field measurements (drifting buoys and current meter deployments) with computer modelling. The results indicate that along the longitudinal axis of Golfo Dulce the gradients in salinity and temperature are maintained by the freshwater input from the three main rivers (Fig. 1). Local wind and topographic steering results in velocity gradients in the upper layer. A three-layer current structure is present at the sill area while in the inner part a two-layer estuarine circulation occurs over slowly moving deep water. The results also described the upper region of Golfo Dulce as having surface waters of low salinities, high temperatures, and low current speeds. On November 1999, surface salinity was near 20 psu with a strong halocline in the upper 5 m. Temperature was 28 oC at the surface and decreased to 17 oC at 60 m. On February 2001 the surface salinity was 33 psu and temperature was 31 oC. Temperature dropped to 15 oC at 60 m. Current speed measurements (drifting buoys) indicated lower velocities (range 3 to 39 cm/s) when compared to those (near 100 cm/s) found in the Gulf of Nicoya estuary. The computer simulations indicated that Golfo Dulce has a less defined estuarine circulation pattern than the Gulf of Nicoya (Voorhis et al., 1983; Vargas, 2016), with an out-flow of low salinity waters in the surface layer and a compensating inflow of coastal waters at intermediate depths (5 to 30 m) in the sill area This pattern, which needs to be explored further with the deployment of anchor stations to measure currents speeds, appears to vary seasonally and relatively strong currents are predicted by the model at the sill most of the year (Fig. 5).
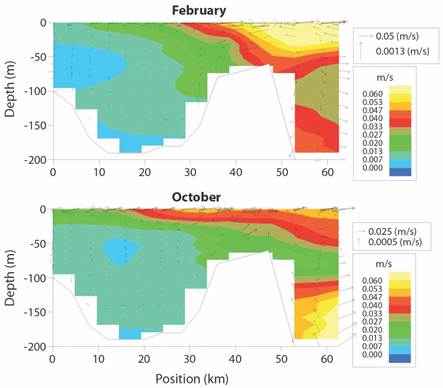
Fig. 5 Simulated monthly (February, dry season - October, rainy season) current speeds (m/sec) in a vertical section along the central Golfo Dulce. Small right upper boxes: note large scale differences between vertical and horizontal currents. The sill area is located between positions 30 to 50 km. Adapted from Svendsen et al. (2006).
Pollution
The Golfo Dulce region is still relatively underdeveloped as most of the Northern shoreline is fringed by rain forests and the major city (Port of Golfito, Golfito Bay, Fig. 1), has a population of about 12 000. However, the concentrations of coliform bacteria in the waters of Golfito Bay were found to be the highest when compared to three sites along the Pacific coast of Costa Rica (García et al., 2006). Concentrations of petroleum hydrocarbons were evaluated at Golfito Bay by Acuña-González et al. (2004), and were found below the reference value of 10 µg/l considered as a borderline for polluted conditions. The presence of trace metals in sediments of Golfito Bay were studied with atomic absorption by García-Céspedes et al. (2004). Maximum concentrations of Cu (128 µg/g), Pb (8.2 µg/g), Zn (109 µg/g) and Fe (7.2 %) may indicate point sources of metal pollution. Concentrations of 13 elements, including metals were analysed by X ray fluorescence in sediment samples from several locations around Golfo Dulce by Salazar et al. (2004). The results indicate no evidence of anthropogenic pollution.
Agricultural activities focus around banana and palm oil plantations. These activities probably have introduced some pollutants into the system, however data is scarce. A study conducted in 1995-1996 analysed residues of organochlorinated pesticides in 47 silty to fine sand sediment samples from several locations in Golfo Dulce (Spongberg & Davies, 1998). The results indicated that pesticide levels were below those reported for contaminated areas elsewhere and many compounds were near the detection limits. The highest concentration found was of pp-DDT, with a concentration of 76 µg/kg dw of sediment. Concentrations of Polychlorinated Biphenyls (PCBs) were also evaluated in Golfo Dulce by Spongberg (2004). Results based on 29 sediment samples indicated that PCB contamination was considered moderate despite the relatively pristine conditions of the area. Maximum PCB concentrations were found at the port of Golfito and reached 16 µg/g dw. PCBs were also studied by Spongberg (2006) in specimens of the sipunculan worm Phascolosoma perlucens from rocky intertidal habitats near the Rincón River (Fig. 1). A concentrations of 67.7 ng/g dw was the highest when compared to those found in this species in other localities along the Pacific coast of Costa Rica. However, PCBs concentrations in sediments and organisms from Golfo Dulce were considered low when compared to other sites worldwide including those in temperate regions (Spongberg & Witter, 2008). A study of residues of pharmaceutical and personal care products (PPCP) in rivers draining into Golfo Dulce was conducted by Spongberg et al. (2011). They reported the presence of 28 compounds including caffeine, salicylic acid and antibiotics such as ciprofloxacin and doxycycline. A recent survey of contaminants in rivers draining into Golfo Dulce detected the presence of the herbicide clomazone, the insecticide triazophos and the fungicides isoprothiolane and propiconazole (Fournier et al., 2019). Other sources of pollutants may have originated far from Golfo Dulce. An early report (Carpenter & Beasley, 1981) studied transuranic radionuclides (Plutonium and Americium) in 200 m deep surface sediments from Golfo Dulce. They found measurable activities of both. A new survey focusing on the evaluation of these and other contaminants in Golfo Dulce is needed.
El Niño (ENSO) and La Niña
Among other external forces acting upon the embayment are the El Niño (ENSO) and La Niña. However, we found only three published reports where this topic has been addressed for Golfo Dulce: A study evaluated the impact of El Niño 1997-1998 on water circulation (Quesada-Alpízar & Morales-Ramírez, 2004) and found this warming event enhanced vertical stratification in Golfo Dulce. Lower temperatures and higher concentrations of dissolved oxygen were observed in deep waters perhaps indicating an intrusion of offshore water over the sill. The second report evaluated the impact of El Niño 1997-1998 on the non-gelatinous zooplankton (Quesada-Alpízar & Morales-Ramírez, 2006). Most of the groups were more abundant above the thermocline. The copepods were the most abundant (71 %), followed by the appendicularians (8 %), ostracods (8 %), polychaete larvae (1.8 %) and chaetognaths (1 %). During El Niño the thermocline was deeper and the abundance of the groups decreased. The third study reported on the impact of El Niño 1997-1998 on the corals of Golfo Dulce. The most visible impact was coral bleaching, which was observed in five species. Psammocora spp were affected most (75 %), followed by Pocillopora damicornis (61 %), and P. elegans (61 %). Mortality was observed in the latter two species (12 and 5%, respectively). Branching colonies were more affected by bleaching than massive species (Jiménez et al., 2001).
Discussion
The published information available for Golfo Dulce emphasizes the unique characteristics of this deep tropical coastal ecosystem. The middle basin allows the study of parameters along a 200 m vertical depth gradient, from oxic to hypoxic and occasionally anoxic conditions, leading to episodes of hydrogen sulphide formation near the bottom. Golfo Dulce is one of the few deep tropical sites where anoxic conditions have been reported and differs from similar environments in the occasional renewal of its basin waters. However, the magnitude and frequency of this renewal needs to be explored further. The sill depth, the extent of the anoxic zone, the concentrations of oxygen and hydrogen sulphide at basin depth and anammox rates have been compared with other localities around the world such as the Baltic and Black seas, the Cariaco Trench in Venezuela, and several European fjords (Anderson & Devol; 1987; Jensen et al., 2008). Both the Gulf of Nicoya and Golfo Dulce are limited at the mouth by waters characteristic of extensive zones on the Eastern Tropical Pacific, particularly deep waters with low oxygen and high nutrient concentrations (Fiedler & Talley, 2006). Previous cooperative research has focused on this feature particularly on the Gulf of Nicoya (Voorhis et al., 1983; Chaves & Birkicht, 1996), but several questions remain to be answered for Golfo Dulce such as the magnitude and frequency of these waters entering the basin.
The coral reefs of Golfo Dulce provide opportunities for ecological studies which include the impact of bioerosion by organisms. The rate of destruction of these reefs has been found to be higher than their growth rate (Quesada-Alpízar & Cortés, 2006). Biodiversity studies need to be intensified in Golfo Dulce and coral reefs and shallow sedimentary environments probably host many yet undescribed species.
Most of the available information on Golfo Dulce comes from short surveys conducted years ago. Also, a great deal of information focuses on specific topics or groups of organisms and a comprehensive multidisciplinary exploration is urgently needed such as that conducted during the R/V Victor Hensen expedition. Thus, there is a need to update and complement the existing information and carry out new evaluations of environmental parameters using state of the art technology. Recent advances in remote sensing, autonomous data loggers, multiparameter electronic sensors, and global positioning (GPS) facilitate data gathering. These new information updates must emphasize obtaining temporal data sets covering longer time and spatial frames. Ample surveys in space and time may facilitate the identification of the impact of local, regional and global stressors acting at different scales. The updates and new evaluations of environmental parameters must be approached under the perspective of increasing coastal development and other threats such as eutrophication (Cloern, 2001; Kennish, 2002; Cloern et al., 2014). El Niño (ENSO) and La Niña temperature oscillations (Wang & Fiedler, 2006), and the impact of climate change on estuarine nutrients (Statham, 2012). The crucial role of ENSO on the decline of Eastern Pacific coral reefs over the past 2 500 years has been emphasized by Toth et al. (2012). Latitudinal comparisons may provide a unique opportunity for the understanding of the functioning of this ecosystem and help to develop predictive models. Selection of target sites, with similar bathymetric characteristics to Golfo Dulce are important in these comparisons and international collaborative efforts are important in this context.
Among the factors affecting coastal embayments worldwide is the likely increase of water temperature due to global warming, that may lead to stronger water stratification, increased microbial respiration, and consequently increased water deoxygenation (Ni et al., 2019). Climate change may intensify the impacts of other stressors such as the dynamics of pollutants (Delorenzo, 2016) or increases in nutrient loads and eutrophication (Cloern, 2001). Comparative studies addressing these and related topics in temperate and tropical systems such as the Gulf of Nicoya and Golfo Dulce may hasten our understanding of these processes and may also contribute to propose on-time remedial measures.
Golfo Dulce has mangrove forests that grow under the influence of semidiurnal tides close to 3 m mean amplitude. Golfo Dulce also has mud flats, sand flats and sandy beaches that provide opportunities for the latitudinal comparative testing of several hypotheses proposed years ago for tropical soft-bottom benthos (Alongi, 1989). These include the diversity of tropical vs temperate epifaunas and infaunas, the study of vertical zonation patterns, the role of opportunistic species, the intensity of predation, competition, colonization, the role of microbes, the estimates of secondary production, and the response to pollution. Of particular relevance is the impact of hypoxic conditions (Wu, 2002). The relatively easy access to intertidal soft-sediment environments in Golfo Dulce facilitates studies on the seasonal population patterns of macrofaunal species, such as that conducted in similar environments in the Gulf of Nicoya by Vargas-Zamora et al. (2015). The polychaete fauna is still relatively unexplored in Golfo Dulce when compared to the Gulf of Nicoya estuary. Moreover, the study of meiofaunal and microfaunal groups is yet to be initiated in Golfo Dulce. Subtidal sediments provide the opportunity to study the influence of grain size and low oxygen concentrations on several groups of the demersal fauna like macro-crustaceans, mollusks and demersal fish such as flatfish, a group represented in Golfo Dulce by 19 species (Vargas et al., 2020).
The extensive shallow region of the upper Gulf of Nicoya and the smaller shallow areas fringing Golfo Dulce lend themselves to the study of the comparative importance of benthic-pelagic coupling and the role of sediment-water oxygen and nutrient exchanges. A recent survey of the literature by Boynton et al. (2018) found 182 reports on this topic worldwide. Most of the reports were from studies conducted in Europe and North America, with only one report from Mexico. The first report from Central America was published by Gómez-Ramírez et al. (2019) and focused on the Gulf of Nicoya estuary.
This literature survey indicates that information on the Golfo Dulce ecosystem has increased significantly since the construction of the Golfo Dulce trophic model by Wolff et al., (1996) which allowed scientific predictions to be made of the functioning of the ecosystem. The following research topics are suggested as priorities for future studies in Golfo Dulce aimed at providing data in support of an updated trophic model or the development of a new one:
Estimate seasonal primary productivity of the water column.
Estimate detrital contribution to the system.
Identify the species and quantify the biomass of the main planktonic autotrophic groups (diatoms, micro-flagellates, dinoflagellates, others).
Identify the species and quantify the biomass of the main water column heterotrophic groups (protozoans, cnidarians, micro-crustaceans, fish, cetaceans).
Identity, quantify, and map the distributions of the main benthic groups (filter and deposit feeding polychaete worms, crustaceans, mollusks, demersal fish).
Identify the frequency and magnitude of the influx of offshore water entering the basin over the sill.
Estimate the frequency and concentrations of H2S formation at the basin.
Describe the temporal and spatial oscillations of the oxic-hypoxic-anoxic interfaces.
Quantify the amounts of contaminants entering the system and their residence time.
Perform paleo-oceanographic investigations based on sediment cores.
Update the trophic model.
Explore new modelling approaches to integrate the available information.
In summary, Golfo Dulce is a unique and accessible tropical ecosystem that provides opportunities for cooperative and comparative research. In view of the increasing impact of local, regional and global stressors there is a need of studies which allow the estimation of the value of the services provided by this ecosystem as a whole and by certain components such as fringing mangrove forests, rocky shores and reefs, tidal flats, and fishery resources as outlined by Barbier et al. (2011).
Ethical statement. The authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interests of any kind, and that we followed all pertinent ethical and legal procedures and requirements. A signed document has been files in the Revista archives.

