











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroduction
Echinoderms are an ancient lineage of invertebrates that exhibit primitive characters like autotomy, regeneration, radial symmetry, and asexual reproduction (James, 1999). The asteroids (sea stars or starfishes) are present in many different marine environments, from deep abyssal depths to the intertidal zone, from the tropics to the poles (Mah & Blake, 2012; O’Hara & Byrne, 2017; Diupotex-Chong, Solís-Marín, & Laguarda-Figueras, 2017). Although asteroids are not typically abundant in marine ecosystems, they influence biodiversity, population dynamics, and can exert control on community structure as predators in intertidal and subtidal ecosystems (Lawrence, 2013; Menge & Sanford, 2013).
Starfish exhibit usually a pentameral symmetry, although numerous-armed starfish have between six to 40 rays (Hotchkiss, 2000a), e.g., Heliaster helianthus (Lamarck, 1816), Pycnopodia helianthoides (Brandt, 1835), Acanthaster planci (Linnaeus, 1758). Starfish rays exhibit a great variety of functions like locomotion (Migita, Mizukamib, & Gunji, 2005; Lawrence, 2013), to force open the shells of mollusks (Lavoie, 1956), parental care (Chia, 1966; Hamel & Mercier, 1995), food detection (Lawrence, 1987; Moore & Lepper, 1997; Dale, 1999), to right themselves (Migita et al., 2005), respiration (Lawrence, 2013), to adhere to the seabed when buffeted by waves (Thomas & Hermans, 1985; Santos, Gorb, Jamar, & Flammang, 2005), and to flee under emergency conditions (Emson & Wilkie, 1980).
Regeneration is a common phenomenon in all actual echinoderm classes (Yousra et al., 2018; Byrne, 2020). It has been particularly well-studied in asteroids, where three main aspects have been identified: 1) regeneration of body parts (arms) following self-induced or traumatic amputation; 2) regeneration of internal organs (pyloric caeca and cardiac stomach) following self-induced or traumatic mutilation, and 3) fission processes (Candia-Carnevali, 2006). Regeneration of body parts and environmental perturbations on the metamorphosis of larvae can result in an abnormality in the number of starfish arms (Hotchkiss, 1979).
Worldwide the largest record of occurrences of abnormal starfishes have been reported in India, where at least 13 species have been listed: Anthenea pentagonula (Lamarck, 1816) (Maheswaran, Narendran, Yosuva & Gunalan, 2015), Asterina lorioli Koehler, 1910 (James, 1999), Astropecten indicus Döderlein, 1888 (James, 1999; Prabhu & Bragadeeswaran, 2012; Chamundeeswari, Saranya, Shanker, Varadharajan, & Rajagopal, 2013), A. karankawai Lawrence, Cobb, Herrera, Durán-González & Solís-Marín, 2018 (ICMYL Unpublished data), Echinaster purpureus (Gray, 1840) (James, 1999), Goniodiscaster vallei (Koehler, 1910) (Maheswaran et al., 2015), Linckia laevigata (Linnaeus, 1758) (James, 1999), L. multifora (Lamarck, 1816) (James, 1999; Maheswaran et al., 2015), L. columbiae Gray, 1840 (Fisher, 1945), Nardoa galatheae (Lütken, 1864) (James, 1999), Pentaceraster regulus (Müller & Troschel, 1842) (James, 1999; Shanker & Vijayanand, 2014), Pisaster ochraceus (Brandt, 1835) (Fisher, 1945), and Protoreaster linckii (Blainville, 1830) (James, 1999; Chelladurai, Balakrishnan, Jayanthi, Ajeesh-Kumar, & Mohanraj, 2015; Chelladurai & Doss, 2016). Other species where abnormal ray numbers have been recorded include Protoreaster nodosus (Linnaeus, 1758), a starfish widely distributed in the Indo-Pacific region (Chim & Tan, 2012) and Archaster angulatus Müller & Troschel, 1842 from the Indian Ocean and western Pacific (Keesing, 2017).
The Panamic Cushion Star Pentaceraster cumingi (Gray, 1840) (Asteroidea: Oreasteridae) is one of the most common starfish in the Tropical Eastern Pacific (TEP) (Solís-Marín et al., 2014; Reyes-Bonilla, Vázquez-Arce, González-Cuéllar, Herrero-Pérezrul, & Weaver, 2016). It has a highly variable coloration, from red-orange or red-green to a gray background color with an overlying bright red network (Kerstitch & Bertsch, 2007). P. cumingi, like most starfish, has pentameric symmetry, is relatively large (up to 17.4 cm of diameter), with an inflated body wall, and immobile spines stud in the upper surface. It is distributed from the Gulf of California, Mexico to Northern Perú, inhabits rocky reefs, patch reefs, and sandy areas, from low intertidal zone up to 183 m deep (Hickman, 1998; Solís-Marín et al., 2014).
The Mexica buried echinoderms in their ritual deposits for at least a half-century. Six species of starfish have been found so far in the offerings, and the presence of P. cumingi was common (10 000 elements have been found in 13 offerings) (Martín-Cao-Romero et al., 2017). While it is true that many animals (or artifacts made from them) were buried as gifts to the supernatural, in most cases they were manipulated as symbols of specific divinities, of particular regions of the universe, or of important cosmic processes (López-Luján, Chávez-Valderas, Zúñiga-Arellano, Aguirre-Molina, & Valentín-Maldonado, 2012).
It is well known that this starfish forms large aggregations, which may correspond to feeding (Reyes-Bonilla, González-Azcárraga, & Rojas-Sierra, 2005) or a reproductive strategy (Reyes-Bonilla et al., 2016). The Panamic Cushion Starfish plays an important ecological role, maintaining the benthic communities in the TEP by feeding on microorganisms and other echinoderms, such as sea urchins, controlling populations of those species (Dee, Witman, & Brandt, 2012; Reyes-Bonilla et al., 2016). The present study focuses on the population density, size structure, and occurrence of abnormalities in the radial pattern of an established population of P. cumingi from the central Mexican Pacific.
Materials and methods
The study was carried out in front of El Novillo islet in Bahía Chamela, Jalisco, in the Central Mexican Pacific (19º33’15” N & 105º07’25” W). The Bahía Chamela was declared a Natural Protected Area in 2002, with the Sanctuary category. Includes seven main islands (Pajarera, Cocinas, La Colorada, San Agustín, San Pedro, San Andrés, and La Negra) and some islets. El Novillo islet (also called Islote Novillos) is rocky with a maximum elevation of 13 m, its surface of 0.73 ha, and its located 1 400 m from mainland. Around the islet, the habitat consists of a sandy plain and mixed rubble substrate, with maximum depths of ~7-10 m (Ríos-Jara et al., 2013). Organisms were sampled from June 2016 to October 2019 (cold and warm seasons). One sampling event was conducted at each season for every year. To determine the density of starfishes in the area, three belt transects of 20x1 m were made, per sampling campaign, by SCUBA diving. Transects were placed parallel to each other and separated two meters between them. A total of 849 starfishes were collected randomly. The measurements that were took were arm length (A, B, C, D, and E) from the mouth center to the tip of arm, minor radius length (r) from mouth center to the end of interradius (C-D), arm breadth (br) at the base of the A-arm (in the case no A-arm, it was replaced with the B-arm), the number of arms, and weight (using a portable electronic balance, 1 g accuracy). All specimens were drained on a plastic tray before recording the weight, to eliminated variability on weighing. Arms position was determined following the Carpenter system, which recognized the C- and D-arm where the madreporite exists in-between, and the opposite arm to madreporite as A-arm (Hotchkiss, 2000a). Starfishes with an abnormal number of arms were photographed in oral and aboral views, to determine the presence and direction of the ambulacral grooves. After being measured and photographed, all individuals were returned to the sea. Water temperature and maximum depth were recorded with a diving computer (Dive Rite Nitek duo). To calculate radius length-weight relationships, pentameric specimens were selected. Abnormal starfishes were examined independently. All data was analyzed using the Minitab 17 software.
Results
Population size structure: For analysis of population size structure, only five armed starfishes were considered. The density of P. cumingi in El Novillo islet exhibited little variation throughout the sampling period (Kruskal-Wallis test H = 9.14, P = 0.243). The highest mean density was found in October 2019 (2.03±0.05 ind/m-2) and the lowest in March 2017 (0.66±0.13 ind/m-2). Mean density for the other months ranged from 1.34 to 1.91 ind/m-2 (Fig. 1). In general, the highest densities were found during the warm season, although no relationship was observed between abundance and temperature. Cold season temperature values ranged from 20.6 to 27.9 °C and warm season ranged from 29.0 to 30.5 °C. Average major radius (R), minor radius (r), and arm breadth (br) were similar between the sampling months, except for br in October 2016 which was the lowest of all (Table 1).
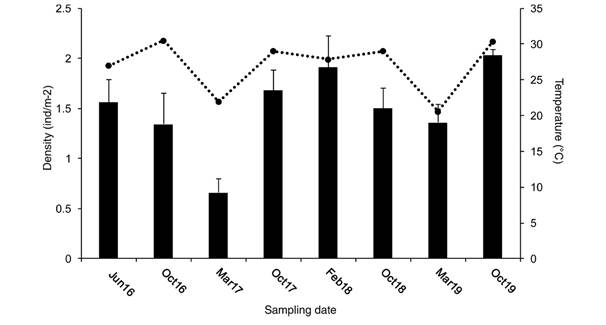
Table 1 Abundance and growth rates of Pentaceraster cumingi by sampling date
| Sampling date | Number of starfish in sample | Number of abnormal starfish* | Mean R (mm) | Mean r (mm) | Mean br (mm) | Depth (m) | Temp (°C) |
| Jun. 2016 | 94 | 0 | 112.1±11.2 | 56.0±5.6 | - | 9.3 | 27.0 |
| Oct. 2016 | 148 | 10 | 112.8±13.7 | 52.4±6.3 | 38.6±14.7 | 7.2 | 30.5 |
| Mar. 2017 | 81 | 2 | 114.8±8.7 | 50.2±5.6 | 52.4±8.6 | 7.2 | 21.9 |
| Oct. 2017 | 119 | 2 | 113.6±12.4 | 46.9±6.1 | 59.2±9.7 | 7.6 | 29.0 |
| Feb. 2018 | 115 | 7 | 114.0±14.1 | 46.8±6.1 | 55.2±10.9 | 6.5 | 27.9 |
| Oct. 2018 | 90 | 0 | 114.9±10.8 | 45.9±5.2 | 53.9±7.2 | 7.7 | 29.0 |
| Mar. 2019 | 80 | 1 | 113.5±11.1 | 47.4±5.6 | 58.8±13.2 | 7.2 | 20.6 |
| Oct. 2019 | 122 | 6 | 123.0±13.0 | 50.0±7.0 | 58.0±9.0 | 7.3 | 30.4 |
| Total | 849 | 28 |
Arm breadth for June 2016 were not measured. *excluded from the mean values.
The maximum size (R) of collected starfish was 160 mm. The average length for normal starfish was 123.8±15.2 mm and the average weight was 326.0±62.4 g (Table 2). The most frequent length classes ranged from 110 to 120 mm (Fig. 2). Larger individuals (> 150 mm) were found in October 2019 and small individuals (< 50 mm) were found in October 2016, 2017 and February 2018. The length classes were widely separated in October 2016 than the other months. Differences in the arm length (t = -3.95; P < 0.05) and body weight (t = 2.82; P = 0.005) were found between cold and warm seasons (Fig. 3).
Table 2 Mean and standard deviation of normal and abnormal starfish Pentaceraster cumingi
| Number of arms | N | Weight (g) | R (mm) | r (mm) | br (mm) | |||||
| A | B | C | D | E | X | |||||
| Normal 5 arms | 821 | 326.0±62.4 | 123.8±15.2 | 122.8±16.2 | 120.9±14.2 | 120.3±13.5 | 119.1±12.7 | - | 50.5±7.3 | 57.5±16.4 |
| Abnormal 5 arms (two ends) | 12 | 308.2±70.2 | 104.9±23.0 | 112.1±20.8 | 111.0±18.5 | 113.7±20.9 | 106.3±18.1 | - | 47.1±6.0 | 51.8±12.2 |
| Abnormal 4 arms (*A) | 3 | 263.0±45.9 | - | 130.0±64.0 | 125.0±18.5 | 125.0±8.1 | 125.0±5.0 | - | 50.0 | 65.0±7.0+ |
| Abnormal 4 arms (*B) | 2 | 284.5±54.4 | 131.0±18.3 | - | 125.5±27.5 | 133.5±19.0 | 133.0±18.3 | - | 43.5±9.1 | 58.0±1.4 |
| Abnormal 4 arms (*C) | 1 | 248 | 120 | 120 | - | 110 | 120 | - | 40 | 80 |
| Abnormal 4 arms (*D) | 1 | 258 | 115 | 114 | 115 | - | 116 | - | 50 | 50 |
| Abnormal 6 arms | 9 | 368.2±41.9 | 124.2±18.3 | 108.0±25.6 | 124.8±20.0 | 117.0±18.5 | 109.8±14.3 | 125.2±19.3 | 53.8±4.2 | 38.6±11.5 |
*A = No arm A; *B = No arm B; *C = No arm C; *D = No arm D.
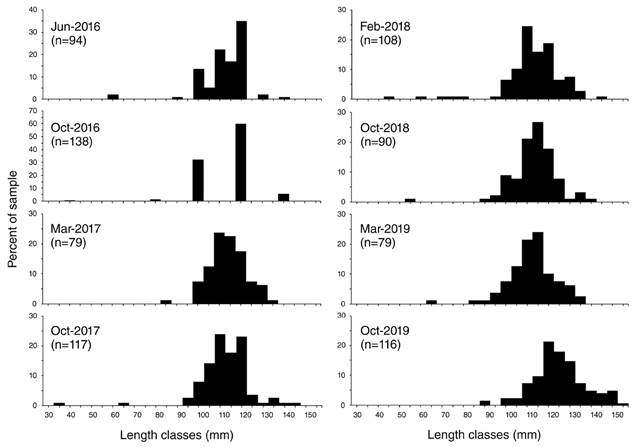
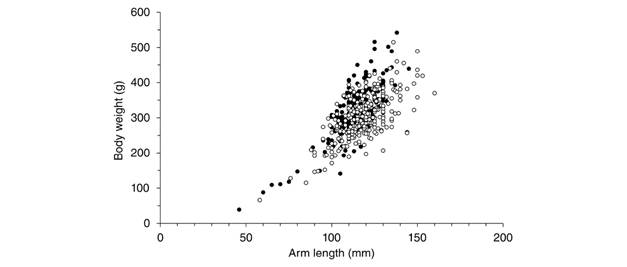
Fig. 3 Length-weight relationships of Pentaceraster cumingi in cold (black circles) and warm (white circles) seasons.
Occurrence of abnormal starfish. A total of 849 individuals of P. cumingi were collected and measured, 28 of them (3.29 %) had an abnormal arm number (i.e. individuals with four arms, six arms, and with a bifurcated arm, Table 1). From all samples, 0.82 % had four arms, 1.06 % had six arms, and 1.41 % had one bifurcated arm. The largest number of abnormal starfish was found in October 2016 when the individual size was the lowest, followed by February 2017 and October 2019.
Abnormalities in P. cumingi radial pattern are caused at least by three processes: 1) by mutilation of one arm, which gives rise to a four-armed starfish (Fig. 4b). In this case, an external mechanism, such as predation, causes the abnormality, and because a remnant of the lost arm is present there is a possibility for regeneration; 2) malformations during development, can give rise to four or six armed starfish (Fig. 4c, Fig. 4e), one individual presented a fusion of two ambulacral grooves (Fig. 4d); and 3) by regeneration, can give rise to a bifurcated arm (Fig. 4f). Bifurcated arms were found only in five-armed starfish and no more than one bifurcated arm was found in the same individual.

Discussion
The species richness of Asteroidea in Bahía Chamela is low. Only three species have been recorded in the bay: Phataria unifascialis (Gray, 1840), Pharia pyramidata (Gray, 1840), and P. cumingi (Ríos-Jara et al., 2013). The low species richness maybe related to a poor sampling effort on starfishes itself. Taking into consideration that some starfish have specific habits, like being nocturnal, roost under rocks, inside cavities, or are buried in soft bottoms, therefore their record can be hard or less detectable with common methods. However, these three species are dominant because they can colonize easy different types of habitats (Luna-Salguero & Reyes-Bonilla, 2010). The occurrence of P. cumingi at Bahía Chamela is continuous throughout the year, with densities that are constant in both seasons (cold and warm). The species reaches population densities from 0.8 ind/50 m2 in the Gulf of California, Mexico (Reyes-Bonilla et al., 2005) to 0.02 ind/m2 in the Gulf of Chiriqui, Panama (Alvarado, Guzmán, & Breedy, 2012). Starfish densities in Bahía Chamela ranged from 0.66 to 2.03 ind/m2, above of those reported for other sites. Population density is similar to those found in aggregation events (probably for a reproduction event) in the Southern Gulf of California, where densities of up to 3 ind/m2 were reported (Reyes-Bonilla et al., 2016). However, these aggregations have been observed only for a few days, unlike the population at Bahía Chamela, which is constant all year. Physical and environmental conditions in the bay such as shallow water, weak currents that provide protection of islands and islets, warmer temperatures, and extensive sandy bottoms can provide sufficient resources (space and food) to maintain a persistent aggregation of P. cumingi (Sloan & Aldridge, 1981; Guillou, 1996; Reyes-Bonilla et al., 2005).
Due to the lack of studies of P. cumingi populations in other localities, it is not possible to compare the mean population sizes present in Bahía Chamela. The P. cumingi population exhibited a seasonal growth pattern, with differences between cold and warm seasons. A possible explanation for this difference is that the length-weight relationship between seasons varied according to factors such a food availability, feeding rate, gonad development, and spawning period (Sebens, 1987). The growth rate for P. cumingi seems to be related to seawater temperature as higher growth rates were recorded in the months when seawater temperature was also higher. Seasonal variation in growth had been described for other starfish species, which usually showed a decrease in growth rate associated with gonad maturation (Freeman, Richardson, & Seed, 2001).
Recruitment events are thought to have occurred between June and August (summer), however, recruits of P. cumingi occur in cryptic habits and their observation is hard. One recruit (< 10 mm) was found attached under a rock in a rocky reef habitat, away from the area where adults were found (Galván-Villa pers. obser.). Possibly, juveniles (> 30 mm) may migrate from rocky reefs to rubble areas, where adults feed to continue its growth. Spatial patterns in the recruitment of starfishes have been associated with different habitat characteristics such as depth, substrate type, temperature, and hydrodynamics (Metaxas, 2013). This selection of habitat has been associated with increased survival, which in turn is affected by the availability of food and predation pressure (Johnson, Sutton, Olson, & Giddins, 1991; Manzur, Barahona, & Navarrete, 2010).
Pentaceraster cumingi belongs to the family Oreasteridae, which is one of the twenty living families of starfishes that are exclusively five-rayed (Hotchkiss, 2000a). Arm number abnormalities are relatively common in the family Oreasteridae. Other studies have reported similar conditions for the congener P. regulus, a common starfish from the Western Central Pacific (James, 1999; Shanker & Vijayanand, 2014), P. linckii, widely distributed in the Indian Ocean (James, 1999; Chelladurai et al., 2015; Chelladurai & Doss, 2016), and P. nodosus, from the Indo-Pacific region (Chim & Tan, 2012). Abnormal arm number have been attributed to several causes like injury, regeneration errors, malnutrition, congenital issues, or damages in the metamorphosis process (Moore, 1974; Hotchkiss, 1979; Watts, Scheibling, Marsh, & McClintock, 1983). The most common cause seems to be by regeneration process (Hotchkiss, 1979). Incidence of mutilation in some starfish has been suggested by fishing gears (Ramsay et al., 2001; Byrne, 2020). It is not considered in this study because the population studied was located inside a protected area where fishing activities are not allowed.
Some lab experiments have shown that changes in environmental factors like salinity can influence ray formation during early development in asteroids (Watts, Scheibling, Marsh, & McClintock, 1982; Marsh, Watts, Chen, & McClintock, 1986; Clark, 1988). Watts et al. (1983) reported a rise of metamorphosed individuals with ray number abnormalities by increasing salinity to 39 %. The incidence of arm abnormalities in P. cumingi from Bahía Chamela was higher in comparison with another species studied. For the starfish Archaster angulatus Müller & Troschel, 1842, Lawrence, Keesing and Irvine (2010), found an incidence of 0.4 % of both four and six arm abnormalities, and Keesing (2017) found an incidence of 1.5 % of four arms and 1.3 % of six arms in the same species. It is not possible to determine the causes of this high level of arm abnormality in P. cumingi, due to the lack of more environmental data. Other factors, such as sample method, size distribution area, and habitat type should be considered for estimation of the incidence of abnormalities.
Four arm condition is not common in starfishes. Some specimens with four evident arms, but with five ambulacral grooves, were found in Bahía Chamela. In these cases, injury or arm mutilation for depredation is the cause of the apparent abnormality, but the arm can be regenerated, as it was observed in other specimens. On the other hand, specimens with four arms and four ambulacral grooves support the synchronic hypothesis, in which abnormalities arise during the metamorphosis, when the pathway to form the rudiments of the five primary rays operates for only a short time, switches off and does not re-occur (Hotchkiss, 2000a). As mentioned above, changes in environmental parameters may be the cause of this abnormality.
Only one specimen with a double ambulacral groove was found. This abnormality is very rare, and it has been reported in a few other sea stars as Asterias forbesi (Desor, 1848), A. problema Lutken, 1872, A. rubens Linnaeus, 1758, Eremicaster vicinus Ludwing, 1907, Pisaster ochraceous segnis Fisher, 1926, and Stephanasterias albula (Stimpson, 1853) (Hotchkiss, 2000b) and it is considered a result of injury or a regeneration process. Hotchkiss (2000b) deduced that this abnormality is a rare result of regeneration, in which the two terminal plates remain coalesced along their inner margin, the ambulacral grooves are kept parallel, and there is just one tip to the double ray. However, in the specimen found in Bahía Chamela, the two ambulacral grooves were fused at the base, and most of this abnormality likely arose during metamorphosis, and not by a regeneration process. A detailed examination of terminal plates, adambulacral, inferomarginal and superomarginal plates is necessary to establish the origin of the abnormality. Other conditions there will develop two tips to the ray but no individuals with this characteristic were found.
Due to the basic skeletal structure of asteroids with flexible arms, they have both species and genera with pentameral and other symmetries (Stephenson, 1967). The supernumerary rays of multiradiate species as Acanthaster, Pycnopodia, Solaster, Crossaster, etc., was explained by the ‘Five-Plus’ hypothesis, that proposes that supernumerary rays develop by independent pathways that operate after the five primary rays have initially formed (Hotchkiss, 2000a). For pentamerous species, the apparition of an extra ray is an unusual event that has not been well studied. The six-armed abnormality was the most common (1.06 %) in P. cumingi. Examination of the ventral view of specimens allowed to confirm the presence of six ambulacral grooves and to separate from specimens with a bifurcation caused by an injury. The presence of well-defined six ambulacral grooves could demonstrate that the abnormality is the result of the metamorphosis process, altered maybe for environmental changes (e.g. Marsh et al., 1986).
The presence of morphological abnormalities is less favorable to survival of individuals, and the incidence of abnormalities can be an indicator of environmental degradation (Jangoux, 1987). Wu, Ji, Wang, and Lv (2012) used mathematical and physical methods to evaluate the superiority of starfish with five arms in comparison with those with a different number of arms, concerning detection, turning over, autotomy, and adherence. They conclude that the optimal number of arms varies under different environmental conditions. In the case of multiradiate taxa some advantages have been identified. Supernumerary rays increase the number of tube feet, then multiradiate asteroids could be more mobile, more resistant to being detached from the seabed, and more successful feeders (Herringshaw, Smith, & Thomas, 2007). Although four-armed starfish exhibited such disadvantages that it is hard for them to survive, the relatively smaller disadvantages of six-armed starfish might allow a few species to exist but the advantages of five arms will permit it to evolve into the dominant pattern of species. As other authors suggest, further studies are needed to understand the causes for the abnormality in pentameral starfishes and the mechanisms that cause it.
Ethical statement: authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

