











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
PermalinkRhinella schneideri (Werner, 1894) (Anura, Bufonidae), formerly known as Bufo paracnemis, is a terrestrial large-bodied anuran amphibian commonly found throughout South America. This species is well-adapted to different habitats, including wetlands, drylands, Cerrado and Atlantic forests, but is also found in open and urban areas (Aquino et al., 2004).
This toad species possesses two types of secretory glands. Mucous glands are distributed all over the skin, with the mucous they secrete serving to protect against transcutaneous water loss in dry/hot climate, and provides conditions for CO2 excretion and O2 uptake (Brazil & Vellard, 1926). Parotoid glands, are large post-orbital serous glands that produce and store a viscous and creamy poisonous secretion composed of proteins, serine proteases, peptides, biogenic amines, cardiotonic steroids and alkaloids (Clarke, 1997; Anjolette et al., 2015).
The parotoid poison is released as mechanism of defense against predators and microorganism infestation (Monti & Cardello, 1994). The parotoid gland of R. schneideri is a rich source of cardiotonic steroids known as bufadienolides, including “marinobufagin, bufalin, telocinobufagin, hellebrigenin, and the atypical 20S,21R-epoxymarinobufagin”, and a great number of studies on the biological activities of skin secretions of R. schneideri (Cunha-Filho et al., 2010) and other species refer to their anti-proliferative and cytotoxic activities in different cancer cells lines (Ferreira et al., 2013; Schmeda-Hirschmann, Quispe, Theoduloz, de Sousa, & Parizotto 2014, Schmeda-Hirschmann et al., 2016).
In vertebrates, the cardiovascular effects of bufadienolides and bufotoxins have been attributed to their ability to increase cardiac contractility and decrease cardiac rate by inhibiting the Na+/K+-ATPase pump in a manner similar to digoxin (Chen & Kovarikova, 1967; Sakate & Oliveira, 2000).
Studies on the effect of R. schneideri poison in nerve-muscle preparations are very scarce. Two recent reports, described such effect on avian and mammalian neuromuscular preparations: in the avian chick biventer cervicis preparation, the methanolic extract of R. schneideri poison was shown to act presynaptically by irreversibly blocking acetylcholine release; no co-participation of post-synaptic nicotinic receptors on the neuromuscular blockade was observed (Rostelato-Ferreira, Dal Belo, Cruz-Höfling, Hyslop, & Rodrigues-Simioni, 2011). Interestingly, in in vitro mice hemidiaphragm preparations R. schneideri poison extract elicited increases in the acetylcholine release, facilitating neurotransmission by a mechanism that suggests the involvement of the Na+K+ATPase pump system (Rostelato-Ferreira, Dal Belo, Leite, Hyslop, & Rodrigues-Simioni, 2014).
The purpose of this study was to investigate by transmission electron microscopy the effect of the methanolic extract of R. schneideri poison in the pre-synaptic region of the neuromuscular junction of mouse nerve-phrenic hemidiaphragm preparation, and be assured on the integrity of the postsynaptic nicotinic receptors by immunofluorescence. The study will allow correlating pharmacological, ultrastructural and immunochemical findings and by this approach to reinforce on the toxic effect exerted by R. schneideri poison at the neuromuscular synapse.
Materials and methods
Reagents and poison: Electron microscopy reagents (EPON EMBed-812 Kit, uranyl acetate, osmium tetroxide, lead citrate and glutaraldehyde) were purchased from Electron Microscopy Sciences (Hatfield, PA, USA). TRITC-conjugated α-BTX was obtained from Sigma Aldrich (St. Louis, MO, USA). Other reagents were obtained from Labsynth Produtos Laboratoriais Ltda. (Diadema, SP, Brazil) and Cristália (Itapira, SP, Brazil).
Poison was collected by manual compression of the large post-orbital parotoid toad glands. The amount of 2 g crude poison was extracted in 50 mL of methanol during three days, at room temperature, lyophilized in a SpeedVac centrifuge (Gao et al., 2010) and stored at -20 °C. The lyophilized methanolic extract was dissolved in Tyrode solution prior to use (Rostelato-Ferreira et al., 2014).
Animals: Male Swiss mice (25-30 g) were obtained from the Multidisciplinary Center for Biological Investigation (CEMIB/UNICAMP). Animals were housed at 23 ± 3 oC under a 12 h light/dark cycle with free access to food and water. This work was approved by an institutional Committee for Ethics in Animal Use (CEUA/UNICAMP, protocol no. 1552-1 and was done in accordance with the ethical guidelines established by the Brazilian Society of Laboratory Animal Science (SBCAL, formerly the Brazilian College of Animal Experimentation - COBEA). The mice were euthanized by halothane inhalation and the diaphragm, with its phrenic nerve branch, was dissected; animal death was confirmed by exsanguination.
Mouse phrenic nerve-hemidiaphragm preparations (PND) (Bülbring, 1946) were incubated for 60 min with 50 µg/mL of the methanolic extract in Tyrode solution (composition in mM: NaCl 137, KCl 2.7, CaCl2 1.8, MgCl2 0.49, NaH2PO4 0.42, NaHCO3 11.9 and glucose 11.1; pH 7.5 at 37 ºC) and constantly aerated by carbogen (95 % O2 - 5 % CO2), while control samples were exposed to Tyrode alone (n = 3-5 per group) at same conditions. The 50 µg/mL extract concentration was selected because it was shown to induce neuromuscular facilitation in PND preparation (Rostelato-Ferreira et al., 2014).
Ultrastructural analysis by Transmission Electron Microscopy (TEM): After neuromuscular facilitation achievement at 60 min of incubation with the methanolic extract of R. schneideri toad (50 µg/mL) or at basal condition when preparations were incubated with Tyrode solution (control) or methanolic extract for 5 min or 60 min (n = 3/group), were washed with the vehicle and then immersed in Karnovsky fixative (2 % paraformaldehyde, 2.5 %glutaraldehyde in 0.1 M sodium cacodylate buffer, pH 7.2).
Under light microscopy, the hemidiaphragm was gently stretched and pinned on wax and then immersed in the fixative solution for 30 min at room temperature. Samples of muscle tissue, 1 mm in width, were collected from the vicinity of phrenic nerve distal branching where axons ending and end-plate regions are more profusely concentrated. The muscle samples were then transferred to flasks containing the same fixative solution and maintained at 4 oC overnight. Muscle samples were washed in 0.1 M sodium cacodylate buffer, post-fixed in 1 % OsO4 for 2 h at room temperature, washed again in the same buffer followed by distilled water, contrasted with 5 % aqueous uranyl acetate solution for 1 h and finally washed in distilled water (Rocha, Leonardo, De Souza, Palma, Da Cruz-Höfling, 2008; Rocha, Souza, Palma, da Cruz-Höfling, & Harris, 2009). After dehydration in an acetone series (30 %, 50 %, 70 %, 90 % and 3 x 100 %, 5 minutes each bath), the samples were immersed in Epon:acetone mixture (2:1, 1:1, 1:2, respectively, 3 hours each bath) and pure Epon resin overnight, followed by fresh Epon for 72 h at 60 oC, for resin polimerization.
Resin blocks were trimmed perpendicular to the long axis of the muscle fibers and ultrathin sections (60 nm thick) were collected on Formvar-coated copper slot grids and contrasted with uranyl acetate (5 % aqueous) followed by lead citrate, 1 minute each. A quantitative analysis of the neuromuscular junction ultrastructure was done using a JEOL JEM-2100 transmission electron microscopy operated at 120 kV. Only those nerve boutons sitting in a well-defined, deeply folded synaptic trough and covered with processes from a terminal Schwann cell, were formally analyzed.
Immunofluorescence: Transversal and longitudinal cryosections of PND (n = 3-5 animals per group), previously incubated in Tyrode solution alone (control) or in methanolic extract for 5 or 60 minutes (n = 3-5 preparation/group) were collected onto subbed glass slides, permeabilized in ethanol and methanol (-20 oC, 10 min each), and in 0.1 % Triton X-100 in phosphate buffered saline - PBS (15 min, room temperature), rinsed with PBS and incubated overnight in a moist chamber at 4 oC with TRITC-conjugated α-BTX and then mounted in glycerin-jelly. The positive immunolabellings was assessed using a fluorescence BX51TF microscope (Olympus Optical C. Ltd., Tokyo, Japan).
Statistical analysis: The results were expressed as the mean ± SEM and statistical comparisons were done using Student’s t-test with P < 0.05 indicating significance. All data analyses were done using Microcal Origin 8 SR4 v.8.0951 (Microcal Software Inc., Northampton, MA, USA) software.
Results
After being exposed, for 5 minutes, to 50 µg/mL of methanolic extract, PND preparations showed a profound and significant 32 % reduction in the number of synaptic vesicles, per nerve terminal, when compared to control (n = 4-6): 74 ± 4 synaptic vesicles for methanolic extract versus 103 ± 11 for Tyrode solution (P < 0.05). After 60 minutes, the number of synaptic vesicles was recovered and reached 121 ± 3, what means 64 % increase relative to the counted after 5 min incubation and by 17 % relative to control (Figure 1).
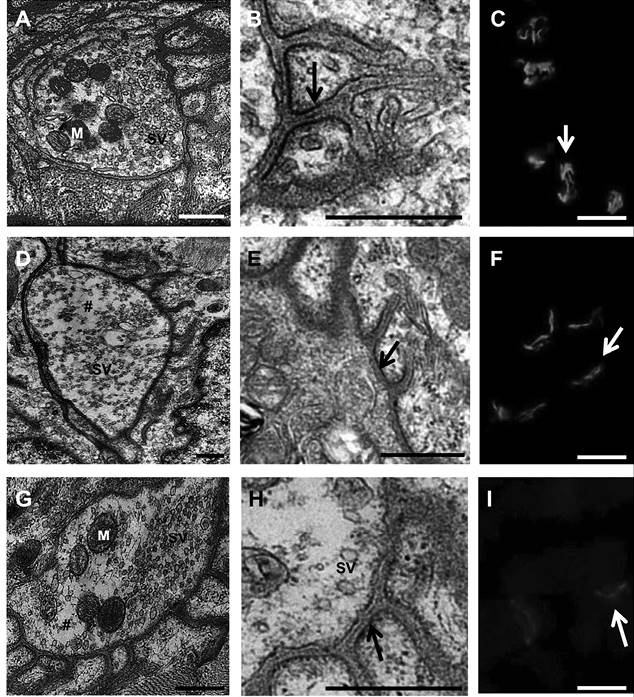
Figure 1 Phrenic nerve-hemidiaphragm preparations (PND). A, B, C - Control preparations incubated with Tyrode solution (vehicle); A, shows the typical terminal bouton with synaptic vesicles (SV) polarized in front of the synaptic cleft and with mitochondria (M) positioned at the opposite site; B, shows detail of the post-synaptic junctional folds exhibiting the classical electron density of the sarcolemma where acetylcholine receptors (AChRs) are abutted (arrows); C, confirms the integrity of the AChRs labeled with TRITC-conjugated α-BTX. D to I - preparations incubated with 50 µg/mL of Rhinella schneideri’s granular gland secretion methanolic extract for 5 min (D, E, F) and 60 min (G, H, I). Notice in D that the synaptic vesicles lost their typical distribution close to the presynaptic axolemma indicating that terminal bouton was remodeled after 5 minutes in contact with the poison extract; in E and F it is shown that the AChRs are maintained in place indicating that the postsynaptic nicotinic receptors were unaffected. In G, it is shown that the population of synaptic vesicles was reestablished after 60 min of incubation with the methanolic extract; in H and I it was shown that a AChR are in place, although with labeling somewhat decreased. Bars: 500 nm.
The methanolic extract of R. schneideri poison did not affect muscle morphology suggesting absence of myotoxic effect; however, the extract exhibits a neurotoxic action, since remodels the nerve terminals and the intramuscular motor innervation in PND preparation. Figure 2 illustrates and describes such effects.
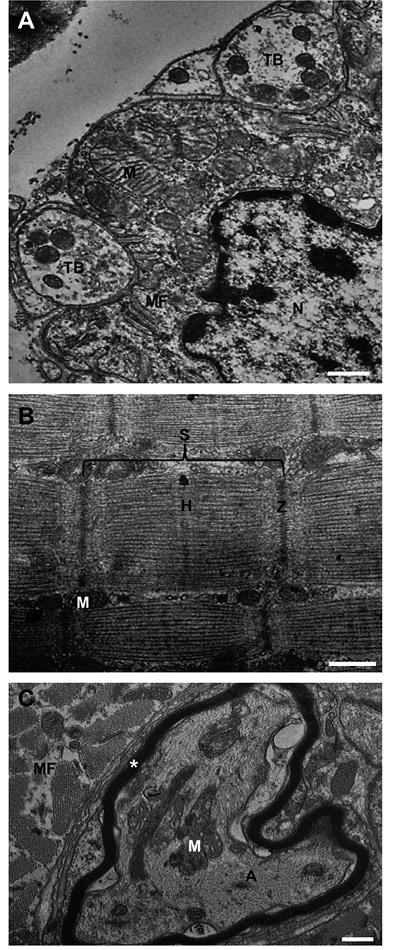
Figure 2 PND preparations exposed to 50 µg/mL of methanolic extract after 60 min incubation. Although the terminal boutons (TB) were reloaded with synaptic vesicles (panel A) the mitochondria (M) in the vicinity of the sole plate nucleus (N) are swollen; despite, mitochondria and sarcomere (S) ultrastructure are well organized indicating preserved muscle fibers morphology (panel B). However, intramuscular motor axons (MA) present intramyelinic vacuoles (*) derived from detachment of inner myelin lamellae. MF = muscle fiber; Z and H = actin myofilaments junction and myosin filament band, respectively. Bars: 500 nm.
Discussion
Previous studies showed that the methanolic extract of R. schneideri poison compounds are capable of affecting neurotransmission in avian and mammalian preparations. The recovery of the number of synaptic vesicles after 60 min of incubation with the methanolic extract of R. schneideri granular (parotoid) gland secretion indicates a transient increase in the neurotransmitter release, which is in accordance to the facilitation observed by Rostelato-Ferreira et al. (2014) in PND preparations.
In their study, Rostelato-Ferreira et al. (2014) reported that diaphragm preparations that were incubated with R. schneideri methanolic extract (50 µg/mL of granular gland secretion) responded with an increase in quantal content (release of acetylcholine) that was inhibited with a non-toxic concentration of ouabain (1 mg/mL), an inhibitor of the Na+/K+-ATPase, after 5 min prior to incubation, indicating that these effects were prevented through the inhibition of Na+-K+-ATPase activity of muscle motor innervation.
The muscle facilitation apparently resulted from enhanced presynaptic neurotransmitter release since electrophysiological measurements indicated an increase in the miniature end plate potentials (MEPPs) frequency (Rostelato-Ferreira et al., 2014). Such response is generally associated with a facilitating effect on neuromuscular transmission (Lundh, Schiller, & Elmqvist, 1977; Rodrigues-Simioni et al., 1997).
Toad poisons are highly cardiotoxic and exert their cardiotoxicity by a digoxin-like effect (Zelnik, 1965; Brownlee, Johnson & Mills, 1990). Ouabain, a cardiotonic steroid derivative, which structurally resembles digoxin, inhibits Na+-K+-ATPase pump activity, and at low concentration enhances ACh release, its quantum content and the frequency of MEPPs (Haimann, Torri-Tarelli, Fesce, & Cascarelli, 1985; Balezina & Lapteva, 2007). We suggest that the decrease in the number of synaptic vesicles in the nerve terminal of preparations incubated with the R. schneideri poison extract might be resulted in enhanced neurotransmitter release elicited by a mechanism involving inhibitory effect on Na+-K+-ATPase pump (Rostelato-Ferreira et al., 2014).
The methanolic extract of R. schneideri poison also caused facilitation of twitch tension followed by neuromuscular blockade in chick biventer cervicis avian preparation. However, the extract was neurotoxic since it remodeled the nerve terminals and intramuscular neurons. The lack of myotoxicity agreed with the findings of Rostelato-Ferreira et al. (2011) who reported that in chick biventer cervicis preparations this methanolic extract of R. schneideri poison did not affect postsynaptic AChRs or muscle contractility, based on the unaltered responses to exogenous ACh or KCl, respectively.
Cruz-Höfling & Rodrigues-Simioni (1998) observed a reduction in the number of synaptic vesicles following the morphological alterations in the myofibrils and mitochondria using guanidine. Rocha et al. (2009) showed that mastoparan Polybia-MPII, isolated from Polybia paulista wasp venom, resulted statistically significant reduction in the volume fraction of the terminal boutons occupied by synaptic vesicles and also showed that there was an associated reduction in cross-sectional area. Both, guanidine and mastoparan Polybia-MPII, promoted a decrease in the number of synaptic vesicle followed by a myotoxic effect. However, the myotoxic effect was not observed after incubation with the methanolic extract from R. schneideri poison, indicating that this poison seems to act mainly at presynaptic site, including in motor innervation.
In conclusion, our results suggest that the methanolic extract of R. schneideri granular gland (parotoid) secretion promotes a transitory decrease in the number of synaptic vesicles after a few minutes of incubation, interfering with synaptic neurotransmission. The immunolabeling of ACh receptors, as evidenced by TRITC-α-bungarotoxin immunoreactivity, suggests that the secretion extract has minor effect on the nicotinic receptors being the primordial site of action the nerve terminal.

