










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.62 n.4 San José Oct./Dec. 2014
Do attacks by jaguars Panthera onca and pumas Puma concolor (Carnivora: Felidae) on livestock correlate with species richness and relative abundance of wild prey?
¿Se correlacionan los ataques de jaguares Panthera onca y pumas Puma concolor (Carnivora: Felidae) al ganado con la riqueza de especies y la abundancia relativa de presas silvestres?
¿Se correlacionan los ataques de jaguares Panthera onca y pumas Puma concolor (Carnivora: Felidae) al ganado con la riqueza de especies y la abundancia relativa de presas silvestres?
Abstract
Attacks by big cats on livestock are one of the major causes of human-felid conflicts and, therefore, an important factor in the conservation of these species. It has been argued that a reduction in natural prey abundance promotes attacks on domestic species, but few studies have tested this statement, and some have delivered contradictory results. We investigated whether the occurrence of attacks to livestock by jaguar and puma relates to the abundance and richness of their natural prey. In the rainy season 2009, we tracked potential prey species counting signs of presence along linear transects in 14 non-attacked cattle farms (control) and in 14 attacked cattle farms in NW Costa Rica. There was a negative relationship between the occurrence of attacks and both species richness (p=0.0014) and abundance (p=0.0012) of natural prey. Our results support the establishment of actions to promote support and recovery of natural prey, in order to diminish attacks on livestock, while maintaining jaguar and puma populations. Rev. Biol. Trop. 62 (4): 1459-1467. Epub 2014 December 01.
Key words: human-wildlife conflicts, livestock depredation, Costa Rica, jaguar, prey abundance, puma, species richness.
Resumen
Los ataques de grandes felinos al ganado son una de las principales causas de conflicto entre humanos y felinos, siendo por ello un tema prioritario para la conservación de estas especies. Se ha argumentado que la reducción en abundancia de presas naturales incrementa la ocurrencia de ataques a las especies domésticas. Sin embargo son pocos los estudios que han evaluado esta afirmación, algunos con resultados contradictorios. Nosotros investigamos cómo la ocurrencia de ataques al ganado, por parte de puma o jaguar, se relaciona con la abundancia y la riqueza de sus presas naturales. Muestreamos las presas potenciales contando los rastros de presencia a lo largo de transectos lineales en 14 fincas sin ataques (control) y en 14 fincas con ataques en el Noroeste de Costa Rica durante la temporada lluviosa de 2009. Encontramos una relación negativa entre la ocurrencia de ataques al ganado y la riqueza (p=0.0014) y abundancia (p=0.0012) de presas naturales. Nuestros resultados respaldan la aplicación de medidas que promuevan el mantenimiento y recuperación de las presas naturales como medida para reducir los ataques al ganado y conservar las poblaciones de puma y jaguar.
Palabras clave: conflicto vida silvestre, ataques al ganado, Costa Rica, jaguar, abundancia de presas, puma, riqueza de especies.
Habitat loss and fragmentation are among the major causes behind biodiversity loss (Tilman et al., 2001). Since top predators require large territories and a relatively high and diverse abundance of prey species (Linnell, Swenson, & Andersen, 2001; Macdonald & Sillero-Zubiri, 2002), they are especially sensitive to landscape changes, and most of their populations are already restricted to protected areas (Michalski, Boulhosa, Faria, & Peres, 2006). Increased interaction between people and big cats, like the jaguar (Panthera onca) and the puma (Puma concolor) tends to escalate conflicts (Saberwal, Gibbs, Chellam, & Johnsingh, 1994; Treves & Karanth, 2003; Kissling, Fernández, & Paruelo, 2009).
Large cat attacks on domestic animals are one of the main reasons for conflict with humans due to economic losses for local communities (Hoogesteijn, Hoogesteijn, & Mondolfi, 1993; Palmeira & Barrella, 2007; Inskip & Zimmermann, 2009). Palmeira (2004) in Brazil, estimated the loss attributable to jaguars as 25 865USD in six years in one big cattle farm, while 50 small farms in Northern Costa Rica recorded total cattle losses of 9 065USD, during one and a half year, when including both jaguar and puma attacks (Amit, Gordillo-Chavez, & Bone, 2013). Despite these tangible losses in that study, perception of damage overestimated real losses, implying the relevance of social and educational factors when addressing these issues.
Frequently, big cats are considered a problem by farmers, who directly retaliate on predators to minimize the incidences on livestock, thus contributing to the decline of the felid populations (Weaver, Paquet, & Ruggiero, 1996; Treves & Naughton-Treves, 2005; Inskip & Zimmermann, 2009). However, top predators like big cats, have an important role in the ecosystem regulation. Their extinction may entail severe effects in ecological processes and, consequently, in entire ecosystems or communities (Terborgh et al., 1999; Gittleman & Gomper, 2005). To understand the reasons that trigger these attacks on cattle, it has been argued that the loss of prey species richness and abundance has a direct effect on the frequency of attacks on cattle (Shaw, 1977; Mondolfi & Hoogesteijn, 1986; Weber & Rabinowitz, 1996; Woodroffe, 2001; Johnson, Vongkhamheng, Hedemark, & Sai-Thongdam, 2006; Inskip & Zimmermann, 2009; Hoogesteijn & Hoogesteijn, 2011). Theoretically, having enough natural food resources makes it unnecessary for predators to enter human territory searching for food. Nevertheless, there are few studies that address this topic, with contradictory results. For example, snow leopard (Uncia uncia) in the Himalayas (Bagchi & Mishra, 2006) and wolf (Canis lupus) in Southern Europe (Meriggi & Lobari, 1996) showed an inverse relationship between the abundance and richness of wild prey and occurrence or frequency of attacks to domestic ungulates. On the other hand, European lynx (Lynx lynx) in Norway (Odden, Herfindal, Linnell, & Andersen, 2008) and France (Stahl, Vandel, Herrenschmidt, & Migot, 2001; Stahl et al., 2002) showed that attacks on sheep match with areas with high abundance of roe deer (Capreolus capreolus). Focusing on jaguar and puma, Polisar et al. (2003) and Cavalcanti and Gese (2010) showed that these big cats selected wild prey even when cattle were available at the Llanos of Venezuela and Pantanal of Brazil, respectively.
In Central America, both puma and jaguar attack livestock and are illegally hunted in retaliation (Amit, Rojas, Alfaro, & Carrillo, 2009). Since the Isthmus is essentially an ecological corridor for many species, it is critical to reduce the conflict between humans and cats in the area for long-term conservation strategies (Rabinowitz & Zeller, 2010). Our study aims to clarify whether there is a relationship between the occurrence of attacks on cattle by these two species of big cats -jaguar and puma- and the richness and abundance of their natural potential prey species. Our hypothesis was that farms without attacks present higher richness and abundance of wild prey than farms with attacks.
Materials and methods
Study area: The study was carried out in the volcanic range called Cordillera de Guanacaste, in Northwestern Costa Rica (11º00’00” - 10º33’50” N and 84º58’00” - 85º39’00” W). The area includes different categories of protected areas, from national parks to private reserves, which usually coincide with the top of the mountains. There is a rainy season from May to November, and a dry season from December to April. Habitat types include Tropical Premountain Wet Forest, Tropical Wet Forest, Tropical Premountain Rain Forest, Tropical Mountain Rain Forest, and Tropical Dry Forest (Holdridge, 1967). Primary forest is mostly found inside protected areas; outside them, the landscape is a mosaic of secondary forest with open areas for agricultural and livestock use with human population scattered in the slopes of mountains, and tourism complementing those traditional economic activities. In these areas, potential natural prey for jaguars and pumas include, among others, species like opossums (Didelphis marsupialis and D. virginiana), armadillo (Dasypus novemcinctus), agouti (Dasyprocta punctata), paca (Cuniculus paca), monkeys (Alouatta palliata, Cebus capucinus and Ateles geoffroyi), coati (Nasua narica), peccaries (Pecari tajacu and Tayassu pecari), white-tailed deer (Odocoileus virginianus) and Central American red brocket (Mazama temama). Poaching is common in the area both in and outside protected areas, and elimination of jaguars and pumas by cattle farmers occurs (Gordillo, 2010; Amit & Fernández, 2012).
Sample selection: A pre-classification was made making a short-list of attacked farms using a pre-existing database (Amit et al., 2013). From May to July of 2010, the study area was visited in order to locate more farms, both attacked and non-attacked. Pre-listed and new farms were inspected in situ and farmers were inquired about the occurrence of attacks by big cats on livestock in their farms or in the surroundings by means of a semi-structured interview on: 1) date of attack, 2) description of traces, 3) aspect of the attacked animal or its remains, and 4) the circumstances of the attack. Cattle farms with attacks were confirmed based on the existence of recent (less than six months) attacks on livestock (cattle and horses) attributed to puma, jaguar or both. We assessed the qualitative evidence reported for the farmers to minimize possible sources of error from other mortality causes comparing with reported descriptions by Hoogesteijn and Hoogesteijn (2011) and Aranda (2000); we discarded doubtful attributions and we did not distinguish between the two cats for the analysis. For farms to be classified as non-attacked, we set the condition of having no attacks for the past two years. In non-attacked farms, the current potential presence of felids was assessed through information provided by officials of the National System of Conservation Areas (SINAC, acronym in Spanish), direct observation of traces and the proximity (less than 10km) to areas with known presence of big cats (Escobedo, 2011). Only farms with extensive production systems were included to standardize for vulnerability to attacks, which can be lower in more intensive husbandry systems (Amit et al., 2013).
In this sense, the extensive management farms in the study area focus on meat production, while the intensive management farms are dedicated to dairy production.
The final selection included 14 attacked farms (AF) and 14 non-attacked farms (NAF). In each farm, we established two linear transects of 500m in length, which were separated from each other a minimum of 500m. A total of 56 transects were delimited in terms of accessibility (pre-existing trails), appropriate conditions of substrate (over 60% free of vegetation and stones), and little human and livestock traffic (less than two records); all factors to maximize detection of wildlife tracks. In the five cases where we could not conduct a continuous 500m transect because the size of the habitat patch, it was divided in two to three smaller transects closer than 300m apart. To characterize the habitat we established four categories: 1) primary forest, 2) secondary forest, 3) open forest and 4) pastures; and to characterize the substrate, we used two categories: 1) optimal, where transect had over 81% of terrain free of vegetation and stones; and 2) suboptimal, where suitability for fingerprinting were between 60% and 80% (Aranda, 2000).
Data collection: Each transect was run once from late July to late August of 2010, when the rainfall allowed a good quality of the substrate throughout the study area, but was not the peak of the rainy season.
A list of potential mammal and bird prey species was built from a bibliographic search. The species included in this list were only considered when diet composition came from empiric studies, including direct observations or scats and gut content analyses. According to this, from all species bigger than 1kg detected, we only excluded the Baird’s tapir (Tapirus bairdii) as prey (unpublished data were not a criteria), but small carnivores were included in the database (Mondolfi & Hoogesteijn, 1986; Rabinowitz & Nottingham, 1986; Chinchilla, 1994; Taber, Novaro, Neris, & Colman, 1997; Núñez, Miller, & Lindzey, 2000; Tavares de Almeida, 2003; Rosas-Rosas, 2006). Species were classified as medium-size (between 1kg and 12kg) and large-size (over 12kg), according to general bibliography. Amphibians, reptiles and smaller birds were not included in the database, since they are generally too small to leave detectable traces and generally constitute a small fraction of biomass consumed by pumas and jaguars.
The presence and relative abundance of potential prey was assessed by recording their traces (i. e. set of signals of an animal as footprints and excrements, belonging to one individual; Sutherland, 1996; Aranda, 2000). Any direct observation on the trail was counted as part of one trace; observations from the trail to the visible surroundings (including tree-dwelling species) were counted also as a trace of presence. In the case of gregarious mammals (primates and peccaries), data were taken as a trace of a troop (group of animals that go together).
Transects were walked by the same two researchers at a speed of about 0.5km/h and only recent one-day-old traces were scored as the afternoon rain ensured fresh traces. Traces were identified in situ with the help of literature (Reid, 1997; Aranda, 2000). In cases of doubt, we took photographs and plaster casts for further analysis by experts. No ambiguous records were used in the analysis. Traces of the same species in different parts of the transect were assessed on whether it was the same specimens to avoid double counting, considering species habits, proximity, similarity and direction (Aranda, 2000).
We calculated species richness for individual transects as the total number of species detected per transect. Four different relative abundance indexes were calculated per transect: 1) Total Prey, 2) Medium-size Prey, and 3) Large-size Prey. These indexes were calculated as the number of traces per transect.
We used generalized linear mixed models (GLMM; Lindstrom & Bates, 1990) using the program R 2.10.1 (R Development Core Team, 2009) and the package nlme (Bates & Maechler, 2009). A model was built including as random factors Transect nested within Farm, and the existence of attacks as binomial response variable. The explanatory variables were the indexes of richness and relative abundance. Correlation among these variables was determined by “Variance Inflation Factor”, using the package car, and the process drop1.
Results
Up to 57 semi-structured interviews to farmers were made, of which 22 had evidences of recent attacks and 34 none. In one case it was not clear the cause of a dead calf.
Habitat and substrate differences between selected AF and NAF were not significant (habitat p=0.511; substrate p=0.603), which allowed us to focus on differences in prey abundance and richness as a possible explanation for attacks on livestock. Total sampling effort was 14km for each AF and NAF.
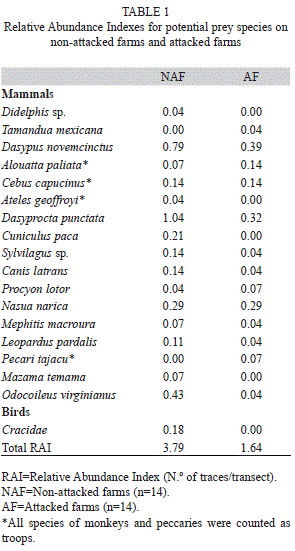
A total of 156 animal traces were recorded from 18 potential prey species in addition to jaguar and Baird’s tapir. Although it was detected nearby, we found no records of puma on transects. The white-lipped peccary (Tayassu pecari) was not detected, while the collared peccary (Pecari tajacu) was detected but in low number. We recorded 16 and 13 species in NAF and AF, respectively (Table 1).
Average species richness was 2.7 in NAF and 1.3 in AF. The two most recorded middle-sized species were the agouti, accounting for 25% of all records, and the armadillo, with 21.7% of traces. The most frequently recorded large-size prey was white-tailed deer (Odocoileus virginianus), accounting 8.6% of all traces (Table 1).
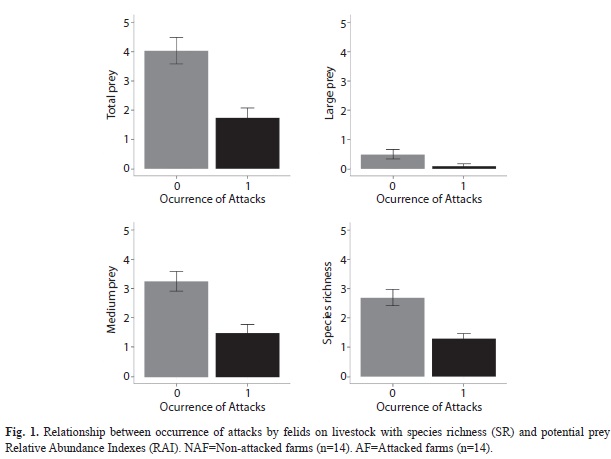
Finally, attacks occur along with low species richness (p=0.001), as well as with low relative abundance of medium-size prey (p=0.001), large-size prey (p=0.046), and total prey (p=0.001) (Table 2, Fig. 1).
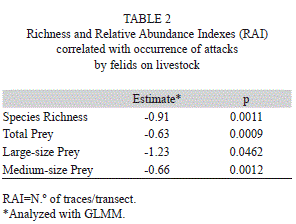
Discussion
Our results showed that lack of natural food resources for predators was correlated with attacks on domestic animals, confirming the trends in the literature (Polisar et al., 2003). This science-based argument support management strategies on cattle farms improving richness and abundance of potential prey species for jaguars and pumas to reduce conflict with humans. Jaguars are known to select wild prey even when cattle is available and increases in wild prey abundance decreases attacks on cattle (Cavalcanti & Gese, 2010). Exceptions to these patterns can be related to other interacting factors like landscape variables, livestock management or even individual cat behavior (Escobedo, 2011; Hoogesteijn & Hoogesteijn, 2011; Moa et al., 2006). Several further variables were collected but the quality and homogeneity of them were not good enough to make more detailed analyses.
Even though this study was limited to a single walk per transect, we were able to detect a significant pattern, and as our aim was not to evaluate the presence and abundance of rare mammals, sampling effort was robust enough.
In Costa Rica, poaching is still widespread, both in private and public protected areas (Altrichter & Almeida, 2002; Amit & Fernández, 2012); that although hunting was declared illegal by the Wildlife Conservation Law (Programa Estado de la Nación, 2013). During our fieldwork, on four occasions we found traces of poachers and farmers regularly reported presence of illegal hunters. In this sense, it is important to emphasize that three of the most coveted species by hunters (paca, armadillo and white-tailed deer; Redford & Robinson, 1987), showed a tendency for higher relative abundance indexes in NAF than in AF.
These results suggest that a variety of stakeholders, wildlife authorities, farmers and communities, have potential benefits in the maintenance and recovery of populations of wild prey species in those places where big cat attacks on livestock are an issue. On one hand, farmers should enforce poaching control in their farms; setting suitable areas within their land, especially water sources, to serve as refuge for prey. On the other hand, wildlife managers and policy-makers should promote an active participation of local communities in projects to monitor wildlife populations –both predators and prey– and to track the incidence of attacks on livestock.
In locations where wild prey populations had been reduced, authorities and NGOs may also promote improving habitat quality and considering reinforcing populations through restocking. In addition, it is essential that managers improve control and sanctioning of poaching. Institutions in general should raise awareness in local communities about the negative effects of poaching in farms, as well as in protected areas.
From a practical point of view, species richness and the relative abundance of prey could be used as indicators of the vulnerability of livestock farms. In the same way, high rates of livestock predation in an area might be a good indicator of problems with wild prey populations.
Since stakeholders face the challenge of balancing the potential negative effects of top predators on the local economy and livelihood, it can help if solutions are framed positively, such as when one source of conflict, prey availability, is acknowledged as a feasible and high priority goal that can be managed.
Acknowledgments
The fieldwork was conducted with the permission of SINAC-MINAET. We thank Fundació Autònoma Solidària and Panthera Foundation for their partial funding, and A. Varela and D. Stein who helped in the field. This study would not have been possible without the interest and cooperation of the farmers, whom allowed entrance to their properties and helped in many ways.
References
Altrichter, M. & Almeida, R. (2002). Exploitation of white-lipped peccaries Tayassu pecari (Artiodactyla: Tayassuidae) on the Osa Peninsula, Costa Rica. Oryx, 36(2), 126-132. [ Links ]
Amit, R., Rojas, K., Alfaro, L. D., & Carrillo. E. (2009). Conservación de Felinos y sus Presas Dentro de Fincas Ganaderas. (Technical Report). Costa Rica: Programa Jaguar-Universidad Nacional. [ Links ]
Amit, R. & Fernández. G. (2012). Interacciones entre jaguar, puma y humanos en Guanacaste, Costa Rica. (Technical Report). Costa Rica: Asociación Confraternidad Guanacasteca. [ Links ]
Amit, R., Gordillo-Chavez, E. J., & Bone, R. (2013) Jaguar and puma attacks on livestock in Costa Rica. Human-Wildlife Interactions, 7(1),77-84. [ Links ]
Aranda, M. (2000). Huellas y otros rastros de los mamíferos grandes y medianos de México. México: Instituto de Ecología, A.C. [ Links ]
Bagchi, S. & Mishra, C. (2006). Living with large carnivores: predation on livestock by the snow leopard (Uncia uncia). Journal of Zoology, 268, 217-224. [ Links ]
Bates, D. & Maechler, M. (2009). lme4: Linear mixed-effects models using S4 classes. R package version 0.999375-32. Retrieved from http://CRAN.R-project.org/package=lme4 [ Links ]
Cavalcanti, S. & Gese, E. (2010). Kill rates and predation patterns of jaguars (Panthera onca) in the Southern Pantanal, Brazil. Journal of Mammalogy, 91(3), 722-736. [ Links ]
Chinchilla, F. A. (1994). La dieta del jaguar (Panthera onca), el puma (Puma concolor), el manigordo (Felis pardalis) y dos métodos de evaluación de la abundancia relativa en el Parque Nacional Corcovado, Costa Rica. (Tesis de Maestría). Universidad Nacional, Costa Rica. [ Links ]
Escobedo, A. (2011). Influencia del paisaje y del tipo de manejo de fincas ganaderas sobre los ataques de grandes felinos (Panthera onca y Puma concolor) a animales domésticos en Costa Rica (Master´s thesis). Centro Agronómico Tropical de Investigación y Enseñanza, Costa Rica. [ Links ]
Gittleman, J. L. & Gomper, M. E. (2005). Plight of predators. The importance of carnivores for understanding patterns of biodiversity and extinction risk. In P. Barbosa & I. Castellanos (Eds.), Ecology of predator-prey interactions (pp. 370-380). Oxford, UK: Oxford University Press. [ Links ]
Gordillo, E. J. (2010). Depredación de ganado por jaguares y pumas en el noroeste de Costa Rica y la percepción de los finqueros hacia el problema. (Tesis de Maestría). Universidad Nacional, Costa Rica. [ Links ]
Holdridge, L. R. (1967). Life zone ecology. San José, Costa Rica: Tropical Science Center. [ Links ]
Hoogesteijn, R., Hoogesteijn, A., & Mondolfi, E. (1993). Jaguar predation and conservation: cattle mortality caused by felines on three ranches in the Venezuelan Llanos. Symposium of the Zoological Society of London, 65, 391-407. [ Links ]
Hoogesteijn, R. & Hoogesteijn, A. (2011). Estrategias para Mitigar la Depredación por Grandes Felinos en Fincas Ganaderas en Latinoamérica: Una Guía. Campo Grande, Brasil: Panthera. [ Links ]
Inskip, C. & Zimmermann, A. (2009). Human-felid conflict: a review of patterns and priorities worldwide. Oryx, 43(1), 18-34. [ Links ]
Johnson, A., Vongkhamheng, C., Hedemark, M., & Sai-Thongdam, T. (2006). Effects of human-carnivore conflict on tiger (Panthera tigris) and prey populations in Lao PDR. Animal Conservation, 9, 421-430. [ Links ]
Kissling, W. D., Fernández, N., & Paruelo, J. M. (2009). Spatial risk assessment of livestock exposure to pumas in Patagonia, Argentina. Ecography, 32, 807-017. [ Links ]
Lindstrom, M. J. & Bates, D. M. (1990). Nonlinear mixed effects models for repeated measures data. Biometrics, 46, 673-687. [ Links ]
Linnell, J. D. C., Swenson, J. E., & Andersen, R. (2001). Predators and people: conservation of large carnivores is possible at high human densities if management policy is favorable. Animal Conservation, 4, 345-349. [ Links ]
Macdonald, D. W. & Sillero-Zubiri, C. (2002). Large carnivores and conflict: lion conservation in context. In A. J. Loveridge, T. Lynam & D. W. Macdonald (Eds.), Lion Conservation Research. Workshop 2: Modelling Conflict (pp. 1-8). Oxford, UK: Wildlife Conservation Research Unit, Oxford University. [ Links ]
Meriggi, A. & Lovari, S. (1996). A review of wolf predation in Southern Europe: does the wolf prefer wild prey to livestock? Journal of Applied Ecology, 33, 1561-1571. [ Links ]
Michalski, F., Boulhosa, R. L., Faria, A., & Peres, C. A. (2006). Human-wildlife conflicts in a fragmented Amazonian forest landscape: determinants of large felid depredation on livestock. Animal Conservation, 9, 179-188. [ Links ]
Moa, P. F., Herfindal, I., Linnell, J. D. C., Overskaug, K., Kvam, T., & Andersen, R. (2006). Does the spatiotemporal distribution of livestock influence forage patch selection in Eurasian lynx? Wildlife Biology, 12, 63-70. [ Links ]
Mondolfi, E. & Hoogesteijn, R. (1986). Notes on the biology and status of the jaguar in Venezuela. In S. D. Miller & D. D. Everett (Eds.), Cats of the world: biology, conservation, and management (pp. 125-146). Washington, D.C.: National Wildlife Federation. [ Links ]
Núñez, R., Miller, B., & Lindzey, F. (2000). Food habits of jaguars and pumas in Jalisco Mexico. Journal of Zoology, 252, 373-379. [ Links ]
Odden, J., Herfindal, I., Linnell, J. D. C., &. Andersen, R. (2008). Vulnerability of domestic sheep to lynx depredation in relation to roe deer density. The Journal of Wildlife Management, 72(1), 276-282 [ Links ]
Palmeira, F. B. L. (2004). Predaçao de bovinos por oncas no norte do estado de Goias (Master´s thesis). Universidade de Sao Paulo, Piracicaba, Brazil. [ Links ]
Palmeira, F. B. L. & Barrella, W. (2007). Conflictos causados pela predaçao de rebanhos domésticos em comunidades quilombolas na Mata Atlantica. Biota Neotropical, 7(1), 119-128. [ Links ]
Polisar, J., Maxit, I., Scognamillo, D., Farrell, L., Sunquist, M. E., & Eisenberg, J .F. (2003). Jaguars, pumas, their prey base, and cattle ranching: ecological interpretations of a management problem. Biological Conservation, 109, 297-310. [ Links ]
Programa Estado de la Nación. (2013). Capítulo IV: Armonía con la Naturaleza. Decimonoveno Informe Estado de la Nación en Desarrollo Humano Sostenible. San José, Costa Rica: Programa Estado de la Nación. [ Links ]
R Core Team (2009). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Retrieved from http://www.R-project.org/. [ Links ]
Rabinowitz, A. R. & Nottingham, B. G. (1986). Ecology and behavior of jaguar (Panthera onca) in Belize, Central America. Journal of Zoology, 210, 149-159. [ Links ]
Rabinowitz, A. & Zeller, K. A. (2010). A Range-wide model of landscape connectivity and conservation for the jaguar Panthera onca. Biological Conservation, 143, 939-945. [ Links ]
Redford, K. H. & Robinson, J. G. (1987). The game of choice: patterns of Indian and colonist hunting in the Neotropics. American Anthropology, 89, 650-667. [ Links ]
Reid, F. A. (1997). A field guide to the mammals of Central America and Southeast Mexico. New York: Oxford University Press. [ Links ]
Rosas-Rosas, O. C. (2006). Ecological status and conservation of jaguars in Northeastern Sonora, Mexico. (Doctoral dissertation). Las Cruces, USA: New Mexico State University. [ Links ]
Saberwal, V. A., Gibbs, J. P., Chellam, R., & Johnsingh, A. J. T. (1994). Lion-human conflict in the Gir Forest, India. Conservation Biology, 8, 501-507. [ Links ]
Shaw, H. G. (1977). Impact of mountain lion on mule deer and cattle in Northwestern Arizona. In R. L. Phillips & C. Jonkel (Eds.), Proceedings of the 1975 predator symposium, June 16-19, 1975, (pp. 17-32). Missoula, USA: University of Montana. [ Links ]
Stahl, P., Vandel, J. M., Herrenschmidt, V., & Migot, P. (2001). Predation on livestock by an expanding reintroduced lynx population: long term trend and spatial variability. Journal of Applied Ecology, 38, 674-687. [ Links ]
Stahl, P., Vandel, J. M., Ruette, S., Coat, L., Coat Y., & Balestra, L. (2002). Factors affecting lynx predation on sheep in the French Jura. Journal of Applied Ecology, 39, 204-216. [ Links ]
Sutherland, W. J. (1996). Ecological Census Techniques. Cambridge, UK: Cambridge University Press. [ Links ]
Taber, A. B., Novaro, A. J., Neris, N., & Colman, F. H. (1997). The food habits of sym-patric jaguar and puma in the Paraguayan Chaco. Biotropica, 29, 204-213. [ Links ]
Tavares de Almeida, R. (2003). Ecología y conservación de felinos silvestres en el área de influencia del Parque Nacional Corcovado, Costa Rica. (Master´s thesis). Universidad Nacional, Costa Rica. [ Links ]
Terborgh, J., Estes, P., Paquet, K., Ralls, D., Boyd-Heger, B., Miller B. J., & Noss, R. (1999). The role of top carnivores in regulating terrestrial ecosistems. In M. E. Soulé & J. Terborgh (Eds.), Continental conservation: Scientific foundations of regional reserve networks (pp. 39-64). Covelo, USA: Island Press. [ Links ]
Tilman, D., Fargione, J., Wolff, B., D’Antonio, C., Dobson, A., Howarth, R., & Swackhamer, D. (2001). Forecasting agriculturally driven global environmental change. Science, 292, 281-284. [ Links ]
Treves, A. & Karanth, K. U. (2003). Human-carnivore conflict and perspectives on carnivores management worldwide. Journal of Conservation Biology, 17(6), 1491-1499. [ Links ]
Treves, A. & Naughton-Treves, L. (2005). Evaluating lethal control in the management of human- wildlife conflict. In R. Woodroffe, S. Thirgood, & A. Rabinowitz (Eds.), People and Wildlife: Conflict or coexistence? (pp. 86-106). Cambridge, UK: Cambridge University Press. [ Links ]
Weaver, J. L., Paquet, P. C., & Ruggiero, L. F. (1996). Resilience and conservation of large carnivores in the Rocky Mountains. Conservation Biology, 10, 964-976. [ Links ]
Weber, W. & Rabinowitz, A. (1996). A global perspective on large carnivore conservation. Conservation Society, 10(4), 1046-1054. [ Links ]
Woodroffe, R. (2001). Strategies for carnivore conservation: Lessons from contemporary extinctions. In J. L. Gittleman, S. M. Funk, D. Macdonald, & R. K. Wayne (Eds.), Carnivore Conservation (pp. 61-92). Cambridge, UK: Cambridge University Press. [ Links ]
Amit, R., Rojas, K., Alfaro, L. D., & Carrillo. E. (2009). Conservación de Felinos y sus Presas Dentro de Fincas Ganaderas. (Technical Report). Costa Rica: Programa Jaguar-Universidad Nacional. [ Links ]
Amit, R. & Fernández. G. (2012). Interacciones entre jaguar, puma y humanos en Guanacaste, Costa Rica. (Technical Report). Costa Rica: Asociación Confraternidad Guanacasteca. [ Links ]
Amit, R., Gordillo-Chavez, E. J., & Bone, R. (2013) Jaguar and puma attacks on livestock in Costa Rica. Human-Wildlife Interactions, 7(1),77-84. [ Links ]
Aranda, M. (2000). Huellas y otros rastros de los mamíferos grandes y medianos de México. México: Instituto de Ecología, A.C. [ Links ]
Bagchi, S. & Mishra, C. (2006). Living with large carnivores: predation on livestock by the snow leopard (Uncia uncia). Journal of Zoology, 268, 217-224. [ Links ]
Bates, D. & Maechler, M. (2009). lme4: Linear mixed-effects models using S4 classes. R package version 0.999375-32. Retrieved from http://CRAN.R-project.org/package=lme4 [ Links ]
Cavalcanti, S. & Gese, E. (2010). Kill rates and predation patterns of jaguars (Panthera onca) in the Southern Pantanal, Brazil. Journal of Mammalogy, 91(3), 722-736. [ Links ]
Chinchilla, F. A. (1994). La dieta del jaguar (Panthera onca), el puma (Puma concolor), el manigordo (Felis pardalis) y dos métodos de evaluación de la abundancia relativa en el Parque Nacional Corcovado, Costa Rica. (Tesis de Maestría). Universidad Nacional, Costa Rica. [ Links ]
Escobedo, A. (2011). Influencia del paisaje y del tipo de manejo de fincas ganaderas sobre los ataques de grandes felinos (Panthera onca y Puma concolor) a animales domésticos en Costa Rica (Master´s thesis). Centro Agronómico Tropical de Investigación y Enseñanza, Costa Rica. [ Links ]
Gittleman, J. L. & Gomper, M. E. (2005). Plight of predators. The importance of carnivores for understanding patterns of biodiversity and extinction risk. In P. Barbosa & I. Castellanos (Eds.), Ecology of predator-prey interactions (pp. 370-380). Oxford, UK: Oxford University Press. [ Links ]
Gordillo, E. J. (2010). Depredación de ganado por jaguares y pumas en el noroeste de Costa Rica y la percepción de los finqueros hacia el problema. (Tesis de Maestría). Universidad Nacional, Costa Rica. [ Links ]
Holdridge, L. R. (1967). Life zone ecology. San José, Costa Rica: Tropical Science Center. [ Links ]
Hoogesteijn, R., Hoogesteijn, A., & Mondolfi, E. (1993). Jaguar predation and conservation: cattle mortality caused by felines on three ranches in the Venezuelan Llanos. Symposium of the Zoological Society of London, 65, 391-407. [ Links ]
Hoogesteijn, R. & Hoogesteijn, A. (2011). Estrategias para Mitigar la Depredación por Grandes Felinos en Fincas Ganaderas en Latinoamérica: Una Guía. Campo Grande, Brasil: Panthera. [ Links ]
Inskip, C. & Zimmermann, A. (2009). Human-felid conflict: a review of patterns and priorities worldwide. Oryx, 43(1), 18-34. [ Links ]
Johnson, A., Vongkhamheng, C., Hedemark, M., & Sai-Thongdam, T. (2006). Effects of human-carnivore conflict on tiger (Panthera tigris) and prey populations in Lao PDR. Animal Conservation, 9, 421-430. [ Links ]
Kissling, W. D., Fernández, N., & Paruelo, J. M. (2009). Spatial risk assessment of livestock exposure to pumas in Patagonia, Argentina. Ecography, 32, 807-017. [ Links ]
Lindstrom, M. J. & Bates, D. M. (1990). Nonlinear mixed effects models for repeated measures data. Biometrics, 46, 673-687. [ Links ]
Linnell, J. D. C., Swenson, J. E., & Andersen, R. (2001). Predators and people: conservation of large carnivores is possible at high human densities if management policy is favorable. Animal Conservation, 4, 345-349. [ Links ]
Macdonald, D. W. & Sillero-Zubiri, C. (2002). Large carnivores and conflict: lion conservation in context. In A. J. Loveridge, T. Lynam & D. W. Macdonald (Eds.), Lion Conservation Research. Workshop 2: Modelling Conflict (pp. 1-8). Oxford, UK: Wildlife Conservation Research Unit, Oxford University. [ Links ]
Meriggi, A. & Lovari, S. (1996). A review of wolf predation in Southern Europe: does the wolf prefer wild prey to livestock? Journal of Applied Ecology, 33, 1561-1571. [ Links ]
Michalski, F., Boulhosa, R. L., Faria, A., & Peres, C. A. (2006). Human-wildlife conflicts in a fragmented Amazonian forest landscape: determinants of large felid depredation on livestock. Animal Conservation, 9, 179-188. [ Links ]
Moa, P. F., Herfindal, I., Linnell, J. D. C., Overskaug, K., Kvam, T., & Andersen, R. (2006). Does the spatiotemporal distribution of livestock influence forage patch selection in Eurasian lynx? Wildlife Biology, 12, 63-70. [ Links ]
Mondolfi, E. & Hoogesteijn, R. (1986). Notes on the biology and status of the jaguar in Venezuela. In S. D. Miller & D. D. Everett (Eds.), Cats of the world: biology, conservation, and management (pp. 125-146). Washington, D.C.: National Wildlife Federation. [ Links ]
Núñez, R., Miller, B., & Lindzey, F. (2000). Food habits of jaguars and pumas in Jalisco Mexico. Journal of Zoology, 252, 373-379. [ Links ]
Odden, J., Herfindal, I., Linnell, J. D. C., &. Andersen, R. (2008). Vulnerability of domestic sheep to lynx depredation in relation to roe deer density. The Journal of Wildlife Management, 72(1), 276-282 [ Links ]
Palmeira, F. B. L. (2004). Predaçao de bovinos por oncas no norte do estado de Goias (Master´s thesis). Universidade de Sao Paulo, Piracicaba, Brazil. [ Links ]
Palmeira, F. B. L. & Barrella, W. (2007). Conflictos causados pela predaçao de rebanhos domésticos em comunidades quilombolas na Mata Atlantica. Biota Neotropical, 7(1), 119-128. [ Links ]
Polisar, J., Maxit, I., Scognamillo, D., Farrell, L., Sunquist, M. E., & Eisenberg, J .F. (2003). Jaguars, pumas, their prey base, and cattle ranching: ecological interpretations of a management problem. Biological Conservation, 109, 297-310. [ Links ]
Programa Estado de la Nación. (2013). Capítulo IV: Armonía con la Naturaleza. Decimonoveno Informe Estado de la Nación en Desarrollo Humano Sostenible. San José, Costa Rica: Programa Estado de la Nación. [ Links ]
R Core Team (2009). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Retrieved from http://www.R-project.org/. [ Links ]
Rabinowitz, A. R. & Nottingham, B. G. (1986). Ecology and behavior of jaguar (Panthera onca) in Belize, Central America. Journal of Zoology, 210, 149-159. [ Links ]
Rabinowitz, A. & Zeller, K. A. (2010). A Range-wide model of landscape connectivity and conservation for the jaguar Panthera onca. Biological Conservation, 143, 939-945. [ Links ]
Redford, K. H. & Robinson, J. G. (1987). The game of choice: patterns of Indian and colonist hunting in the Neotropics. American Anthropology, 89, 650-667. [ Links ]
Reid, F. A. (1997). A field guide to the mammals of Central America and Southeast Mexico. New York: Oxford University Press. [ Links ]
Rosas-Rosas, O. C. (2006). Ecological status and conservation of jaguars in Northeastern Sonora, Mexico. (Doctoral dissertation). Las Cruces, USA: New Mexico State University. [ Links ]
Saberwal, V. A., Gibbs, J. P., Chellam, R., & Johnsingh, A. J. T. (1994). Lion-human conflict in the Gir Forest, India. Conservation Biology, 8, 501-507. [ Links ]
Shaw, H. G. (1977). Impact of mountain lion on mule deer and cattle in Northwestern Arizona. In R. L. Phillips & C. Jonkel (Eds.), Proceedings of the 1975 predator symposium, June 16-19, 1975, (pp. 17-32). Missoula, USA: University of Montana. [ Links ]
Stahl, P., Vandel, J. M., Herrenschmidt, V., & Migot, P. (2001). Predation on livestock by an expanding reintroduced lynx population: long term trend and spatial variability. Journal of Applied Ecology, 38, 674-687. [ Links ]
Stahl, P., Vandel, J. M., Ruette, S., Coat, L., Coat Y., & Balestra, L. (2002). Factors affecting lynx predation on sheep in the French Jura. Journal of Applied Ecology, 39, 204-216. [ Links ]
Sutherland, W. J. (1996). Ecological Census Techniques. Cambridge, UK: Cambridge University Press. [ Links ]
Taber, A. B., Novaro, A. J., Neris, N., & Colman, F. H. (1997). The food habits of sym-patric jaguar and puma in the Paraguayan Chaco. Biotropica, 29, 204-213. [ Links ]
Tavares de Almeida, R. (2003). Ecología y conservación de felinos silvestres en el área de influencia del Parque Nacional Corcovado, Costa Rica. (Master´s thesis). Universidad Nacional, Costa Rica. [ Links ]
Terborgh, J., Estes, P., Paquet, K., Ralls, D., Boyd-Heger, B., Miller B. J., & Noss, R. (1999). The role of top carnivores in regulating terrestrial ecosistems. In M. E. Soulé & J. Terborgh (Eds.), Continental conservation: Scientific foundations of regional reserve networks (pp. 39-64). Covelo, USA: Island Press. [ Links ]
Tilman, D., Fargione, J., Wolff, B., D’Antonio, C., Dobson, A., Howarth, R., & Swackhamer, D. (2001). Forecasting agriculturally driven global environmental change. Science, 292, 281-284. [ Links ]
Treves, A. & Karanth, K. U. (2003). Human-carnivore conflict and perspectives on carnivores management worldwide. Journal of Conservation Biology, 17(6), 1491-1499. [ Links ]
Treves, A. & Naughton-Treves, L. (2005). Evaluating lethal control in the management of human- wildlife conflict. In R. Woodroffe, S. Thirgood, & A. Rabinowitz (Eds.), People and Wildlife: Conflict or coexistence? (pp. 86-106). Cambridge, UK: Cambridge University Press. [ Links ]
Weaver, J. L., Paquet, P. C., & Ruggiero, L. F. (1996). Resilience and conservation of large carnivores in the Rocky Mountains. Conservation Biology, 10, 964-976. [ Links ]
Weber, W. & Rabinowitz, A. (1996). A global perspective on large carnivore conservation. Conservation Society, 10(4), 1046-1054. [ Links ]
Woodroffe, R. (2001). Strategies for carnivore conservation: Lessons from contemporary extinctions. In J. L. Gittleman, S. M. Funk, D. Macdonald, & R. K. Wayne (Eds.), Carnivore Conservation (pp. 61-92). Cambridge, UK: Cambridge University Press. [ Links ]
1. Unit of Ecology, CREAF, Autonomous University of Barcelona, Edifici C, Cerdanyola del Vallès, 08193, Catalonia, Spain; aburgas@gmail.com, bernat.claramunt@uab.cat
2. Department of Wildlife Ecology and Conservation, University of Florida, 110 Newins-Ziegler Hall, PO Box 110430, Gainesville, 32611-0430, FL, USA; jaguar.rar@gmail.com
Received 28-I-2014. Corrected 28-VI-2014. Accepted 29-VII-2014.


{kind=link}
{kind=link}