











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.62 no.3 San José jul./sep. 2014
Long-term (2002-2011) changes on Cetengraulis edentulus (Clupeiformes: Engraulidae) fisheries in Guanabara Bay, Brazil
Variaciones a gran escala (2002-2011) de las pesquerías de Cetengraulis edentulus (Clupeiformes: Engraulidae) en la bahía de Guanabara, Brasil
Variaciones a gran escala (2002-2011) de las pesquerías de Cetengraulis edentulus (Clupeiformes: Engraulidae) en la bahía de Guanabara, Brasil
Ana Clara Sampaio Franco1*, Daniel Shimada Brotto2*, David Man Wai Zee3* & Luciano Neves dos Santos1
Abstract
C. edentulus accounts for the major fishery resource in Guanabara Bay, but there are only few studies about its captures.This study analyzed the long term changes on C. edentulus catches in Guanabara Bay, focusing especially on temporal variation on capture effort, boat storage temperature, and fish price and size.We assessed the 2002-2011 database of fish landings at the pier of Rubi S.A., with records of year, month, total catch (kg), number of fish per kg, boat storage temperature (oC), and price per kg to boat owner (US$). Those variables differed among years and months (ANOVA; p<0.01) with strong yearXmonth interactions for all, probably related to C. edentulus reproductive and recruitment seasons and environmental conditions of Guanabara Bay. Generalized additive models revealed that high C. edentulus catches were associated with few boat loadings of large-sized fishes, which were conserved in higher boat storage temperatures and attained high market prices. Our long-term study stressed that variations on C. edentulus fishery were related to changes on boat storage temperature, fish size, price paid per kg, and frequency of landings, and that these descriptors are correlated with C. edentulus ecological traits and high phytoplankton availability. Nevertheless, further studies should be performed to address whether less evident factors, such as the fluctuation of concurrent Sardinella brasiliensis stock, would explain the temporal dynamics of C. edentulus stocks in Guanabara Bay.
Key words: estuary, long-term, fisheries, landings, Cetengraulis edentulus.
Resumen
Cetengraulis edentulus se distribuye por todo el Atlántico Sur y es la especie más explotada comercialmente en la bahía de Guanabara, Brasil. Entre 2002-2011 se analizaron los desembarques pesqueros de la empresa Rubi S.A., las capturas totales (kg), número de peces por kg, temperatura de almacenamiento en la embarcación (°C) y el precio pagado por kg de C. edentulus al propietario de la embarcación (US$). Todas las variables difirieron entre años y meses (ANOVA, p<0.01), con interacciones años × meses significativas para todas, probablemente a causa de los períodos reproductivo y de reclutamiento de C. edentulus y a las condiciones ambientales de la bahía. Modelos aditivos generalizados indicaron que capturas más elevadas de C. edentulus estuvieron asociadas con un menor número de desembarques, pero de peces más grandes, los cuales fueron conservados a temperaturas más elevadas y obtuvieron precios más altos de mercado. Aunque los cambios en los descriptores estén relacionados con los atributos ecológicos de C. edentulus y la productividad del fitoplancton, se necesitan más estudios para investigar si otros factores, tales como la fluctuación de los stocks de Sardinella brasiliensis, pueden afectar la dinámica temporal de los stocks de C. edentulus en la bahía.
Palabras clave: estuario, gran-escala, pesquerías, desembarques, Cetengraulis edentulus.
Engraulid fishes, commonly known as anchovies, are widely distributed in tropical and sub-tropical waters (Mcgowan & Berry, 1983). Anchovies are recognized as an important trophic link in coastal food chains, since they convert planktonic biomass into forage for higher level consumers (Hildebrand, 1963). Most engraulids spawn at coastal areas in the inner continental shelf, and recruitment occurs often in protected, shallow areas that provide food and shelter against predators (Silva, Araújo, Azevedo, & Mendonça, 2003). Adults perform seasonal migrations between estuarine and oceanic areas (MacGregor & Houde, 1996), aggregating in large shoals frequently targeted by commercial fisheries. Many species of anchovies are economically important in several regions in South America, namely Engraulis ringens in Peru, Engraulis anchoita in Argentina, and Cetengraulis edentulus in Venezuela (Whitehead, 1977) and Guanabara Bay, Brazil (Jablonski, Azevedo, & Moreira, 2006).
The Atlantic anchoveta C. edentulus (Cuvier) is an engraulid fish, widely distrib- uted throughout the Southwest Atlantic, occur- ring from Antilles and Cuba to Southern Brazil (Whitehead, Nelson, & Wongratana, 1988). C. edentulus first matures at 118mm total length (L50 for both sexes), reaching 200mm at maximum (Silva et al., 2003; Souza-Conceição, Rodrigues-Ribeiro, & Castro-Silva, 2005). This species feeds mainly on phyto- plankton and is widely tolerant to changes in environmental conditions (Krumme, Keuthen, Barletta, Saint-Paul, & Villwock, 2008; Giar- rizzo & Krumme, 2009).It is often associated with muddy substrates of inner bay zones, where salinity is low, temperatures are high, and phytoplankton biomass is high due to the great input of organic waste (Sergipense, Caramaschi, & Sazima, 1999; Gay, Sergipense, & Rocha, 2000; Silva et al., 2003; Souza- Conceição et al., 2005; Araújo, Silva, Santos, & Vasconcellos, 2008).
C. edentulus began to be used as human food and fish meal since early 1980s, in part due to declining stocks of other traditionally exploited species, such as Sardinella brasiliensis (Whitehead et al., 1988; Jablonski et al., 2006), currently accounting for the major fishery resource in coastal bays of Rio de Janeiro, Southeastern Brazil (Araújo et al., 2008). Overall, commercial vessels operating at Guanabara Bay use purse seines (35-40m length; 12mm mesh) to catch preferentially C. edentulus but also other small pelagic fish species (i.e. S. brasiliensis) within the area (Vianna, 2009). Fish are captured near the surface, often together with a few benthic species, because of the shallow waters where those fishing operations are performed.
C. edentulus is the dominant exploited species in Guanabara Bay (12 427t in 2004), accounting for 69% of total fish production. Despite its low value per kilogram, the significant bulk of Atlantic anchoveta landings in Guanabara Bay provided more than US$1 million for local trade (Jablonski et al., 2006). While economically important, the dynamics of C. edentulus catches in Guanabara Bay is barely known, especially in relation to interannual population trends and seasonal fluctuations on its capture. In this sense, this study aims to analyze the long term changes on C. edentulus catches in Guanabara Bay, focusing especially on temporal variation on capture effort, boat storage temperature, and fish price and size between 2002 and 2011.
Materials and Methods
Guanabara Bay is an estuary around 400km2 located on the coast of Rio de Janeiro state (22°50’ S - 43°10’ W; Fig. 1). The climate is tropical-humid with a warm-rainy season (December-March) and a cold-dry sea- son (July-August) (Paranhos & Mayr, 1993; Valentin et al., 1999). Guanabara Bay is 36km long and has 3x109m3 of water volume, mean depth of 7.6m, and maximum depth of 30m at the entrance channel (Valentin et al., 1999). Tidal currents account for most water circulation in this bay, with a micro tidal regime (i.e., 0.7m of amplitude), semi-diurnal tides, and maximum currents of 1.6m/s that result in 23 days of water residence time (Kjerfve, Ribeiro, Dias, Filippo, & Quaresma, 1997). Substrate is predominantly muddy, with increasingly contribution of sand toward the outer zone, and the salinity ranges from 21.0 to 34.5 (Kjerfve et al., 1997). Guanabara Bay is surrounded by one of the largest metropolitan areas in Brazil (i.e. Rio de Janeiro city), with more than 11 million inhabitants. As consequence, the water quality is poorer at the inner zone, because of the main land proximity and the restrict circulation, while better conditions (i.e. higher transparency and dissolved oxygen) are found toward the outer zone, more influenced by adjacent oceanic waters.
The statistical data examined proceed from a period of ten years of fish landings at the pier of Rubi S.A., a fish processing industry located near Guanabara Bay at Rio de Janeiro state. Fish was caught by boats that operate with purse seines in the region from January of 2002 to June of 2011. Digital database for each fish landing with records of year, month, total catch (kg), number of fish per kg, boat storage temperature (oC), and price paid per kg to boat owner (US$) was assessed. Two-way analysis of variance (ANOVA) were applied on those variables to detect interannual and seasonal variations, followed by Tukey tests (p<0.05) whenever significant differences were recorded.
Generalized additive models (GAMs) were applied to analyse the relationship of C. edentulus catches with others variables. GAMs are an extension of generalized linear models that do not assume a particular functional relationship between the response variable and the predictor. The model complexity of GAMs was chosen by the stepwise selection procedure using the Akaike information criterion (AIC). AIC considers not only the goodness of fit but also parsimony, penalizing more complex models (Burnham & Anderson, 1998).
Results
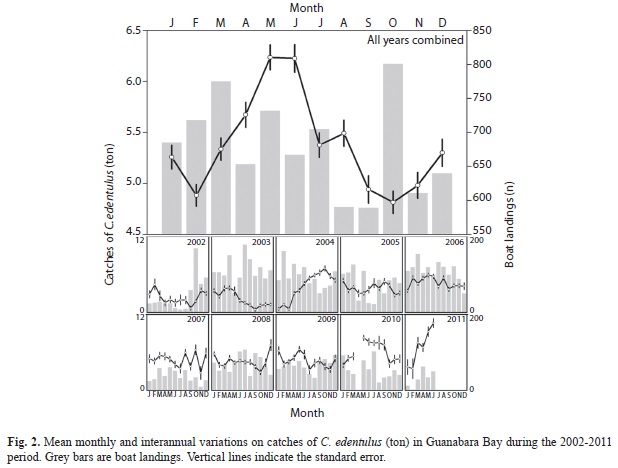
The catches of C. edentulus varied significantly among years (F7, 7 866=80.9, p<0.01) and months (F10, 7 866=4.2, p<0.01), with a strong year × month interaction (F87, 7 866=11.2, p<0.01). Overall, the catches increased progressively through years, with low yields being recorded in 2002-2003, intermediate values within the 2004-2009 period, and greater catches at 2010-2011 (Tukey test, p<0.05 for all, Fig. 2). High C. edentulus yields were found in May-June whereas low values were recorded during February and September-November (Tukey test, p<0.05 for all), a trend that was not clearly observed for any year separately.
The amount of C. edentulus per kg differed significantly with years (F7, 7 866=2 947.3, p<0.01) and months (F10, 7 866=187.6, p<0.01), with a significant year × month interaction (F87, 7 866=56.7, p<0.01). Higher numbers of C. edentulus per kg were recorded during 2003-2004, indicating a greater contribution of relatively small-sized individuals in the catches (Tukey test, p<0.05 for all, Fig. 3). Since 2004, the number of C. edentulus per kg decreased progressively with years, suggesting increased contributions of larger fishes after that. Small C. edentulus were particularly abundant in the catches of February-April, with a trend of increased contributions of larger fishes toward September, when the lowest numbers of C. edentulus per kg were recorded (Tukey test, p<0.05 for all, Fig. 3). This monthly trend was only clearly observed for 2004, with high among-years variance.
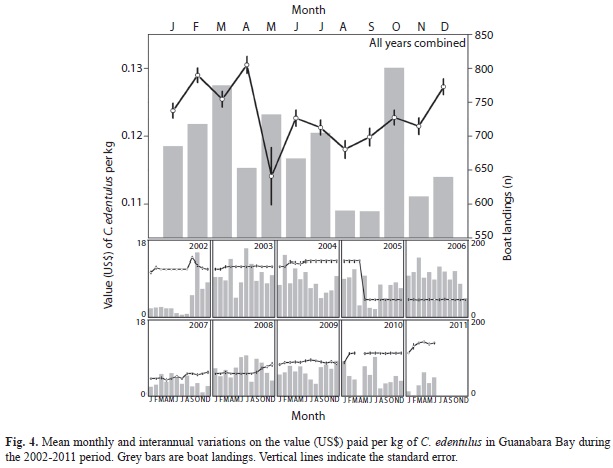
The value paid per kg of C. edentulus differed significantly with years (F7, 7 866=13 356.4, p<0.01) and months (F10, 7 866=67.8, p<0.01), and a significant year × month interaction was also detected (F87, 7 866=270.8, p<0.01). High values were recorded during 2002-2004, with a sharp decrease after April, 2005 (Tukey test, p<0.05 for all, Fig. 4). C. edentulus remained at low values per kg until the middle of 2007, when the values began to increase progressively, reaching at 2011 the similar high values before 2005. The highest values were recorded during December-April, peaking at February and April (Tukey test, p<0.05 for all).This temporal trend was not clearly detected for any separated year, probably because of the high monthly variation among years, as observed especially in May.
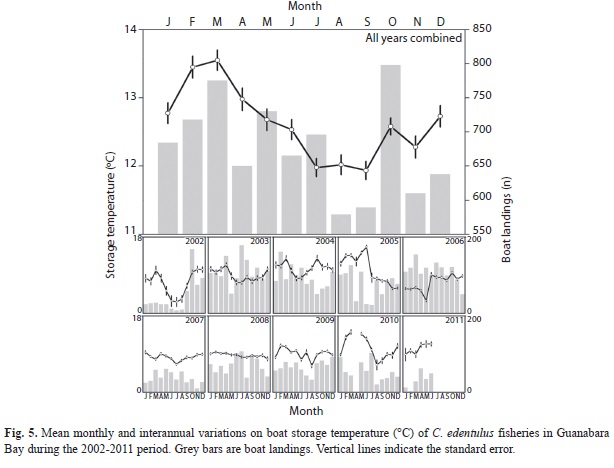
Boat storage temperature changed significantly among years (F7, 7 839=77.5, p<0.01) and months (F10, 7 839=17.8, p<0.01), with a strong year × month interaction (F87, 7 839=15.8,
p<0.01). Higher temperatures were recorded during 2004-2005 and in 2010 (Tukey test, p<0.05 for all), with a broad variation among years (Fig. 5). Low storage temperatures were observed during July-September whereas high temperatures were recorded in February and March.
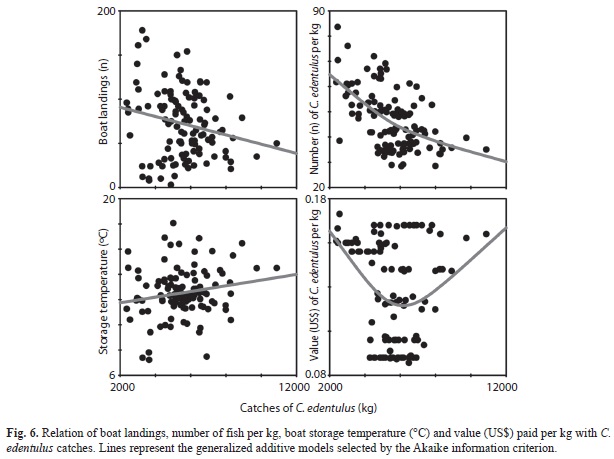
The response curves (GAMs) of boat landings, number of C. edentulus per kg, stor- age temperature and value of C. edentulus per kg also varied with the catches (Fig. 6). The AIC revealed a negative linear trend and April (Tukey test, p<0.05 for all).This temporal trend was not clearly detected for any of boat landings (F1, 111=5.2, p=0.02) with C. edentulus catches, while a positive linear trend was selected for the storage temperature (F1,111=3.8, p=0.05).A non-linear response was found for the number of C. edentulus per kg (AIC, F1, 110=5.6, p=0.02), with an exponential decrease with the catches. A non-linear relationship was also found for the value paid per kg of C. edentulus (AIC, F1, 110=28.7, p<0.01), with a trend of great values at the extremes of the catches and a broad oscillation of values at intermediate captures.
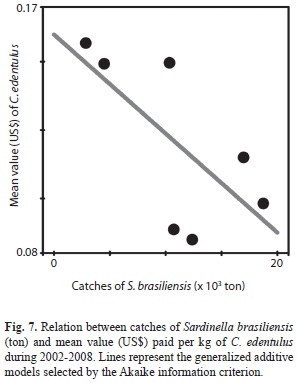
No relationship (AIC, GAM) was found between the catches for C. edentulus (our data) and S. brasiliensis (data retrieved from CEPSUL Rio de Janeiro state from 2001 to 2007: http://www.icmbio.gov.br/cepsul/biblioteca/acervo-digital/37-download/estatistica/111-estatistica.html “Accessed 20 Sep 2013”). However, AIC selected a negative linear response (AIC, F1, 6=4.4; p=0.09) between the mean values of C. edentulus per kg and S. brasiliensis catches (time lag of a year; Fig. 7). This trend indicates that C. edentulus attained lower market prices in response to increased catches of S. brasiliensis in the preceding year.
Discussion
Our findings revealed significant interannual and seasonal changes on C. edentulus catches, which can be explained by non-mutually exclusive reasons. Although not directly assessed in this study, the increasing catches of C. edentulus from 2002 to 2011 could be related to the environmental changes experienced by Guanabara Bay (Paranhos & Mayr, 1993; Caddy, 2000). Changes were reported in the trophic chain of eutrophicated semi-enclosed marine systems, toward the dominance of planktivorous fish, in response to the increased nutrient enrichment, bottom hypoxia episodes, and top predator removal (Caddy, 2000). Guanabara Bay has undergone a continuous eutrophication process, mostly because of its location in the Rio de Janeiro city, one of the most urbanized areas of Brazil. Only 15% of the total urban sewage released into Guanabara Bay, which includes organic matter, nutrients, hydrocarbons, heavy metals, and organic pollutants, is being adequately treated (Kjerfve et al., 1997). High chlorophyll-a concentrations (i.e. hypertrophic) were founded and bottom hypoxic conditions related to high urban sewage loads, mostly at the inner zone of Guanabara Bay, where large densities of C. edentulus were found (Ribeiro & Kjerfve, 2002). Therefore, the prevalent eutrophic conditions in the Bay (Lavrado, Carvalho, Mayr, & Paranhos, 1991; Sergipense et al., 1999) seem to be highly profitable to C. edentulus stocks, since its planktivorous-pelagic habit allow a successful use of food resources with few risks for this species to face bottom anoxic conditions.
Seasonal changes on C. edentulus catches could be explained by sociocultural and bio- logical effects. Captures peaked on May and June, which is probably related to Holy Week, the most important Christian festival characterized by remarkable increase on fish product consumption. However, the lowest catches of C. edentulus on October and February could be influenced by spawning season and recruit- ment, respectively. The reproductive season of this species occurs from September to Decem- ber (spring) at Guanabara Bay (ACS Franco, personal observations) when adults seem to migrate for spawning, decreasing C. edentulus availability. The spawning site of C. edentulus in Guanabara Bay is rather controversial on whether adult fish migrate to inner estuarine areas or outer oceanic zones to breed. However, the rare occurrence of C. edentulus at outside bay habitats, the preference of this species for mud bottoms, and the presence of eggs and juveniles in Guanabara Bay, sug- gests that the entire reproductive cycle occurs inside this area (Sergipense & Sazima, 1995; Silva et al., 2003; Castro, Bonecker, & Valentin, 2005; Tubino, Monteiro-Neto, Moraes, & Paes, 2007). The increased contribution of juveniles from the previous spawning event to the stock probably explained the low catches on February, since these small sized fish are less vulnerable to the mesh commonly used by purse seine fishery.
The amount of C. edentulus per kg was considered here as an estimation of fish size. According to this descriptor, it was clear that catches consisted mainly of relatively small- sized individuals within 2003 and 2004, sug- gesting stock overexploitation or the use of weak criteria for minimum fish size commercialized at the industry until 2004. Since then, the number of C. edentulus per kg decreased progressively through years, which can be related to an increased contribution of large-sized fishes. Seasonal changes on fish size agree with the temporal trend found for C. edentulus catches, with a dominance of large individuals in September (i.e. reproduction) and small-sized individuals in February (i.e. recruitment).
Storage temperature fluctuated considerably through years in our study, with a trend of stabilization in 2006 to 2008, when industry supported fishermen with ice. Seasonal changes were also found for temperature, which are mostly related to climate, with high temperatures in warmer months (December to February; summer) and low temperatures in colder months (June to August; winter). The storage temperature and time are linked to the quality of fish meal for human consumption (Köse & Erdem, 2004). The exposition of Engraulis encrasicolus to high temperatures after catching may lead to histamine production, which can cause human intoxication if consumed (Rossano, Mastrangelo, Ungaro, & Riccio, 2006). The ideal storage condition for E. encrasicolus is a 3:2 ice/fish proportion, a temperature of 4°C or less and a maximum storage time of 2 days (Köse & Erdem, 2004; Visciano, Campana, Annunziata, Vergara, & Ianieri, 2007). Therefore, the action of provid- ing ice for fishermen could be an interesting measure for the industry to maintain fish within the quality standards.
Our results also revealed the importance of catch success to the four major descriptors of C. edentulus fisheries in Guanabara Bay. When assessed in conjunction, GAMs revealed that great C. edentulus catches were associated with few boat loadings, compounded mostly of large-sized fishes, which were conserved in higher boat storage temperatures and attained high market prices. The unexpected relationship between few boat landings and great catches could be related to periods of C. edentulus over abundance, in which the boat storage could be fulfilled with a single purse seine set, resulting in a single landing. On the other hand, several boat landings and low catches might be associated with periods of high stock scattering, requiring more purse seine sets to compensate the failures or low individual and temperature. Although GAMs suggested that highest prices were paid for low and high C. edentulus catches, those trends seems to be more related to industry strategies and market demand than to species ecological traits.
Other variables than those directly assessed in the present study could also have influence on C. edentulus fisheries. The growth of C. edentulus fisheries in Guanabara Bay was apparently triggered by the collapse, since the later 1990s, of Sardinella brasiliensis stocks (a higher prized target) in the Brazilian coast (Cergole, Saccardo, & Rossi-Wongtschowski, 2002). C. edentulus also attained lower market prices in response to increased catches of S. brasiliensis in Rio de Janeiro State at the preceding year, suggesting important interferences of concurrent fish stocks. Further studies are thus needed to evaluate the influence of other and perhaps less evident, factors on C. edentulus fishery, in a system of multiple exploited stocks.
Our long-term study revealed that seasonal and interannual variations on C. edentulus fishery were related to changes on boat storage temperature, fish size, price paid per kg, and frequency of landings. Our findings also stressed that these descriptors are influenced by C. edentulus ecological traits (i.e. reproductive and recruitment seasons) and changes on ecosystem conditions (i.e. eutrophication). Nevertheless, further studies should be performed to address whether less evident (i.e. the fluctuation of concurrent stocks) or even commonly dismissed (i.e. fleet characteristics, improvement of boat and fishing gears) factors would explain the long and short scale temporal dynamics of C. edentulus stocks in Guanabara Bay.
Acknowledgments
We especially thank Conservas Rubi Ltda and Marítima Pescados SA for providing financial support and allow for the use of C. edentulus database. We are also grateful to Graduate Course in Neotropical Biodiversity (PPGBIO-UNIRIO), Laboratory of Theoretical and Applied Ichthyology (LICTA-UNIRIO), and Veiga de Almeida University (UVA) for providing the logistic support. This work was also funded by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro, Brazil (research grant to LN Santos, E-26/110.166/2010; E-26/111.548/2010), National Council for Scientific and Technological Development (CNPq), Brazil (Programa PELD – Baía de Guanabara), Brazilian Federal Agency for Support and Evaluation of Graduate Education (CAPES), Brazil, (graduate grant to ACS Franco).
References
Araújo, F. G., Silva, M. A., Santos, J. N. S., & Vasconcellos, R. M. (2008). Habitat selection by anchovies (Clupeiformes: Engraulidae) in a tropical bay at Southeastern Brazil. Neotropical Ichthyology, 6, 583-590. [ Links ]
Burnham, K. P. & Anderson, D. R. (1998). Model selection and multimodel inference: a practical information- theoretic approach. New York: Springer. [ Links ]
Caddy, J. F. (2000). Marine catchment basin effects versus impacts of fisheries on semi-enclosed seas. Journal of Marine Sciences, 57, 628-640. [ Links ]
Castro, M. S., Bonecker, A. C. T., & Valentin, J. L. (2005). Seasonal variation in fish larvae at the entrance of Guanabara Bay, Brazil. Brazilian Archieves of Biology and Technology, 48(1): 121-128. [ Links ]
Cergole, M. C., Saccardo, S. A., & Rossi-Wongtschowski, C. L. D. B. (2002). Fluctuations in the spawning stock biomass and recruitment of the Brazilian sardine (Sardinella brasiliensis): 1977-1997. Revista Brasileira de Oceanografia, 50, 13-25. [ Links ]
Gay, D., Sergipense, S., & Rocha, C. F. D. (2000). Ocorrên- cia e distribuição de Cetengraulis edentulus (Cuvier) (Teleostei, Engraulidae) na Laguna de Itaipu, Niterói, Rio de Janeiro Brasil. Revista Brasileira de Zoologia, 17, 463-472. [ Links ]
Giarrizzo, T. & Krumme, U. (2009). Temporal patterns in the occurrence of selected tropical fishes in mangrove creeks: implications for the fisheries management in the North Brazil. Brazilian Archives of Biology and Technology, 52, 679-688. [ Links ]
Hildebrand, S. F. (1963). Family Engraulidae. Memoiries Sears Foundation for Marine Research, 1, 152-249. [ Links ]
Jablonski, S., Azevedo, A. F., & Moreira, L. H. A. (2006). Fisheries and conflicts in Guanabara Bay, Rio de Janeiro, Brazil. Brazilian Archives of Biology and Technology, 49, 79-91. [ Links ]
Kjerfve, B., Ribeiro, C. H. A., Dias, G. T. M., Filippo, A. M., & Quaresma, V. S. (1997). Oceanographic characteristics of an impacted coastal bay: Baía de Guanabara, Rio de Janeiro, Brazil. Continental Shelf Research, 17, 1609-1643. [ Links ]
Köse, S. & Erdem, M. E. (2004). An investigation of quality changes in anchovy (Engraulis encrasicolus, L. 1758) stored at different temperatures. Turkish Journal of Veterinary and Animal Sciences, 28, 575-82. [ Links ]
Krumme, U., Keuthen, H., Barletta, M., Saint-Paul, U., & Villwock, W. (2008). Resuspended intertidal micro- phytobenthos as major diet component of planktivorous Atlantic anchoveta Cetengraulis edentulus (Engraulidae) from equatorial mangrove creeks. Ecotropica, 14, 121-128. [ Links ]
Lavrado, H. P., Carvalho, V., Mayr, L. M., & Paranhos, R. (1991). Evolution (1980-1990) of ammonia and dissolved oxygen in Guanabara Bay, RJ, Brazil. In O. T. Magoon, H. Converse, V. Tippie, L. T. Thomas, & D. Clark (Eds.), Coastal Zone ‘91: Proceedings of the 7th Symposium on Coastal and Ocean Management (pp. 3234-3245). New York: American Society of Civil Engineers. [ Links ]
MacGregor, J. M. & Houde, E. D. (1996). Onshore- Offshore pattern and variability in distribution and abundance of bay anchovy Anchoamitchilli eggs and larvae in Cheasapeake Bay. Marine Ecology Progress Series, 138, 15-25. [ Links ]
Mcgowan, M. F. & Berry, F. H. (1983). Clupeiformes: Development and Relationships. In: H. G. Moser, W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall, & S. L. Richardson (Eds.), Ontogeny and Systematics of Fishes (pp. 108-126). New York: American Society of Ichthyologists and Herpetologists. [ Links ]
Paranhos, R. & Mayr, L. M. (1993). Seasonal Patterns of Temperature and Salinity in Guanabara Bay, Brazil. Fresenius Environmental Bulletin, 2, 647-652. [ Links ]
Ribeiro, C. H. A. & Kjerfve, B. (2002). Anthropogenic influence on the water quality in Guanabara Bay, Rio de Janeiro, Brazil. Regional Environmental Change, 3, 13-19. [ Links ]
Rossano, R., Mastrangelo, L., Ungaro, N., & Riccio, P. (2006). Influence of storage temperature and freezing time on histamine level in the European anchovy Engraulis encrasicolus (L., 1758): a study by capi- llary electrophoresis. Journal of Chromatography B, 830, 161-164. [ Links ]
Sergipense, S. & Sazima, I. (1995). Variações sazonais de ocorrência e tamanho em duas espécies de Engraulididae (Osteichthyes) na Baía de Sepetiba, Rio de Janeiro. Revista Brasileira de Biologia, 55, 491-501. [ Links ]
Sergipense, S., Caramaschi, E. P., & Sazima, I. (1999). Morfologia e hábitos alimentares de duas espécies de Engraulidae (Teleostei, Clupeiforme) na Baía de Sepetiba, Rio de Janeiro. Revista Brasileira de Oceanografia, 47, 173-188. [ Links ]
Silva, M. A., Araújo, F. G., Azevedo, M. C. C., & Mendonça, P. (2003). Distribuição espacial e temporal de Cetengraulis edentulus (Cuvier) (Actinopterygii, Engraulidae) na Baía de Sepetiba, Rio de Janeiro, Brasil. Revista Brasileira de Zoologia, 20, 577-581. [ Links ]
Souza-Conceição, J. M., Rodrigues-Ribeiro, M., & Castro-Silva, M. A. (2005). Dinâmica populacional, biologia reprodutiva e o ictioplâncton de Cetengraulis edentulus na enseada do Saco dos Limões, Florianópolis, Santa Catarina, Brasil. Revista Brasileira de Zoologia, 22, 953-961. [ Links ]
Tubino, R. A., Monteiro-Neto, C., Moraes, L. E. S., & Paes, E. T. (2007). Artisanal fisheries production in the coastal zone of Itaipu, Niterói, RJ, Brazil. Brazilian Journal of Oceanography, 55, 187-197. [ Links ]
Valentin, J. L., Tenenbaum, D. R., Bonecker, A., Bonecker, S. L. C., Nogueira, C. R., Paranhos, R., & Villac, M. C. (1999). Caractéristiques hydrologiques de La Baie de Guanabara (Rio de Janeiro, Brésil). Journal de Recherche Océanographique, 24, 33-41. [ Links ]
Vianna, M. (2009). Diagnóstico da cadeia produtiva da pesca marítima no Estado do Rio de Janeiro: Relatório da pesquisa. Rio de Janeiro: FAERJ: SEBRAE-RJ. Retrieved from http://www.que-rodiscutiromeuestado.rj.gov.br/CMS/uploads/publicacoes/09.08.13-09.37.56-diagnostico_pesca_maritima_comercial_rio_de_janeiro.pdf. [ Links ]
Visciano, P., Campana, G., Annunziata, L., Vergara, A., & Ianieri, A. (2007). Effect of storage temperature on histamine formation in Sardina pilchardus and Engraulis encrasicolus after catch. Journal of Food Biochemistry, 31, 577-588. [ Links ]
Whitehead, P. J. P. (1977). Engraulidae. In: W. Fischer (Ed.), FAO species identification sheets for fishery purposes. Western Central Atlantic (Fishing Area 31) (Vol 2). Rome: Food and Agriculture Organization of the United Nations. [ Links ]
Whitehead, P. J. P., Nelson, G. J., & Wongratana, T. (1988). FAO species catalogue. Clupeoid of the world (Suborder Clupeoidei). An annotated and illustrated catalogue of the herrings, sardines, pilchards, sprats, shads, anchovies and wolf-herrings. (Vol. 7, pp. 1-579). Rome: FAO Fisheries Synopsis, Rome. [ Links ]
Burnham, K. P. & Anderson, D. R. (1998). Model selection and multimodel inference: a practical information- theoretic approach. New York: Springer. [ Links ]
Caddy, J. F. (2000). Marine catchment basin effects versus impacts of fisheries on semi-enclosed seas. Journal of Marine Sciences, 57, 628-640. [ Links ]
Castro, M. S., Bonecker, A. C. T., & Valentin, J. L. (2005). Seasonal variation in fish larvae at the entrance of Guanabara Bay, Brazil. Brazilian Archieves of Biology and Technology, 48(1): 121-128. [ Links ]
Cergole, M. C., Saccardo, S. A., & Rossi-Wongtschowski, C. L. D. B. (2002). Fluctuations in the spawning stock biomass and recruitment of the Brazilian sardine (Sardinella brasiliensis): 1977-1997. Revista Brasileira de Oceanografia, 50, 13-25. [ Links ]
Gay, D., Sergipense, S., & Rocha, C. F. D. (2000). Ocorrên- cia e distribuição de Cetengraulis edentulus (Cuvier) (Teleostei, Engraulidae) na Laguna de Itaipu, Niterói, Rio de Janeiro Brasil. Revista Brasileira de Zoologia, 17, 463-472. [ Links ]
Giarrizzo, T. & Krumme, U. (2009). Temporal patterns in the occurrence of selected tropical fishes in mangrove creeks: implications for the fisheries management in the North Brazil. Brazilian Archives of Biology and Technology, 52, 679-688. [ Links ]
Hildebrand, S. F. (1963). Family Engraulidae. Memoiries Sears Foundation for Marine Research, 1, 152-249. [ Links ]
Jablonski, S., Azevedo, A. F., & Moreira, L. H. A. (2006). Fisheries and conflicts in Guanabara Bay, Rio de Janeiro, Brazil. Brazilian Archives of Biology and Technology, 49, 79-91. [ Links ]
Kjerfve, B., Ribeiro, C. H. A., Dias, G. T. M., Filippo, A. M., & Quaresma, V. S. (1997). Oceanographic characteristics of an impacted coastal bay: Baía de Guanabara, Rio de Janeiro, Brazil. Continental Shelf Research, 17, 1609-1643. [ Links ]
Köse, S. & Erdem, M. E. (2004). An investigation of quality changes in anchovy (Engraulis encrasicolus, L. 1758) stored at different temperatures. Turkish Journal of Veterinary and Animal Sciences, 28, 575-82. [ Links ]
Krumme, U., Keuthen, H., Barletta, M., Saint-Paul, U., & Villwock, W. (2008). Resuspended intertidal micro- phytobenthos as major diet component of planktivorous Atlantic anchoveta Cetengraulis edentulus (Engraulidae) from equatorial mangrove creeks. Ecotropica, 14, 121-128. [ Links ]
Lavrado, H. P., Carvalho, V., Mayr, L. M., & Paranhos, R. (1991). Evolution (1980-1990) of ammonia and dissolved oxygen in Guanabara Bay, RJ, Brazil. In O. T. Magoon, H. Converse, V. Tippie, L. T. Thomas, & D. Clark (Eds.), Coastal Zone ‘91: Proceedings of the 7th Symposium on Coastal and Ocean Management (pp. 3234-3245). New York: American Society of Civil Engineers. [ Links ]
MacGregor, J. M. & Houde, E. D. (1996). Onshore- Offshore pattern and variability in distribution and abundance of bay anchovy Anchoamitchilli eggs and larvae in Cheasapeake Bay. Marine Ecology Progress Series, 138, 15-25. [ Links ]
Mcgowan, M. F. & Berry, F. H. (1983). Clupeiformes: Development and Relationships. In: H. G. Moser, W. J. Richards, D. M. Cohen, M. P. Fahay, A. W. Kendall, & S. L. Richardson (Eds.), Ontogeny and Systematics of Fishes (pp. 108-126). New York: American Society of Ichthyologists and Herpetologists. [ Links ]
Paranhos, R. & Mayr, L. M. (1993). Seasonal Patterns of Temperature and Salinity in Guanabara Bay, Brazil. Fresenius Environmental Bulletin, 2, 647-652. [ Links ]
Ribeiro, C. H. A. & Kjerfve, B. (2002). Anthropogenic influence on the water quality in Guanabara Bay, Rio de Janeiro, Brazil. Regional Environmental Change, 3, 13-19. [ Links ]
Rossano, R., Mastrangelo, L., Ungaro, N., & Riccio, P. (2006). Influence of storage temperature and freezing time on histamine level in the European anchovy Engraulis encrasicolus (L., 1758): a study by capi- llary electrophoresis. Journal of Chromatography B, 830, 161-164. [ Links ]
Sergipense, S. & Sazima, I. (1995). Variações sazonais de ocorrência e tamanho em duas espécies de Engraulididae (Osteichthyes) na Baía de Sepetiba, Rio de Janeiro. Revista Brasileira de Biologia, 55, 491-501. [ Links ]
Sergipense, S., Caramaschi, E. P., & Sazima, I. (1999). Morfologia e hábitos alimentares de duas espécies de Engraulidae (Teleostei, Clupeiforme) na Baía de Sepetiba, Rio de Janeiro. Revista Brasileira de Oceanografia, 47, 173-188. [ Links ]
Silva, M. A., Araújo, F. G., Azevedo, M. C. C., & Mendonça, P. (2003). Distribuição espacial e temporal de Cetengraulis edentulus (Cuvier) (Actinopterygii, Engraulidae) na Baía de Sepetiba, Rio de Janeiro, Brasil. Revista Brasileira de Zoologia, 20, 577-581. [ Links ]
Souza-Conceição, J. M., Rodrigues-Ribeiro, M., & Castro-Silva, M. A. (2005). Dinâmica populacional, biologia reprodutiva e o ictioplâncton de Cetengraulis edentulus na enseada do Saco dos Limões, Florianópolis, Santa Catarina, Brasil. Revista Brasileira de Zoologia, 22, 953-961. [ Links ]
Tubino, R. A., Monteiro-Neto, C., Moraes, L. E. S., & Paes, E. T. (2007). Artisanal fisheries production in the coastal zone of Itaipu, Niterói, RJ, Brazil. Brazilian Journal of Oceanography, 55, 187-197. [ Links ]
Valentin, J. L., Tenenbaum, D. R., Bonecker, A., Bonecker, S. L. C., Nogueira, C. R., Paranhos, R., & Villac, M. C. (1999). Caractéristiques hydrologiques de La Baie de Guanabara (Rio de Janeiro, Brésil). Journal de Recherche Océanographique, 24, 33-41. [ Links ]
Vianna, M. (2009). Diagnóstico da cadeia produtiva da pesca marítima no Estado do Rio de Janeiro: Relatório da pesquisa. Rio de Janeiro: FAERJ: SEBRAE-RJ. Retrieved from http://www.que-rodiscutiromeuestado.rj.gov.br/CMS/uploads/publicacoes/09.08.13-09.37.56-diagnostico_pesca_maritima_comercial_rio_de_janeiro.pdf. [ Links ]
Visciano, P., Campana, G., Annunziata, L., Vergara, A., & Ianieri, A. (2007). Effect of storage temperature on histamine formation in Sardina pilchardus and Engraulis encrasicolus after catch. Journal of Food Biochemistry, 31, 577-588. [ Links ]
Whitehead, P. J. P. (1977). Engraulidae. In: W. Fischer (Ed.), FAO species identification sheets for fishery purposes. Western Central Atlantic (Fishing Area 31) (Vol 2). Rome: Food and Agriculture Organization of the United Nations. [ Links ]
Whitehead, P. J. P., Nelson, G. J., & Wongratana, T. (1988). FAO species catalogue. Clupeoid of the world (Suborder Clupeoidei). An annotated and illustrated catalogue of the herrings, sardines, pilchards, sprats, shads, anchovies and wolf-herrings. (Vol. 7, pp. 1-579). Rome: FAO Fisheries Synopsis, Rome. [ Links ]
1. UNIRIO - Universidade Federal do Estado do Rio de Janeiro, Programa de Pós-graduação em Biodiversidade Neotropical. Avenida Pasteur, 458 sala 314A, 22290-240, Rio de Janeiro, Brasil; anaclara306@gmail.com, luciano.santos@unirio.br
2. UVA – Universidade Veiga de Almeida, Departamento de Ciências da Saúde. Rua Ibituruna, 108, 20271-901, Rio de Janeiro, Brasil; danshima@ig.com.br
3. UERJ – Universidade do Estado do Rio de Janeiro, Departamento de Oceanografia Física. Rua São Francisco Xavier 524, sala 4025-E, 20020-100, Rio de Janeiro, Brasil; davidzee.mw@gmail.com
Received 22-XI-2014. Corrected 30-IV-2014. Accepted 28-V-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}