











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.62 no.2 San José abr./jun. 2014
Morphology and genetics of Anadenanthera colubrina var. cebil (Fabaceae) tree from Salta (Northwestern Argentina)
Morfología y genética del árbol Anadenanthera colubrina var. cebil (Fabaceae) en Salta (noroeste de Argentina)
Morfología y genética del árbol Anadenanthera colubrina var. cebil (Fabaceae) en Salta (noroeste de Argentina)
*Dirección para correspondencia:
Abstract
Anadenanthera colubrina var. cebil is an important tree species for its cultural, economic, and medicinal uses in South America. In order to characterize A. colubrina populations, we collected fruits from four different sites (San Bernardo, El Cebilar, Metán and El Gallinato) within the species distribution area in Salta Province, Northwestern Argentina. For this, a total of 75 fruits and seeds per site were collected and described using morphological (fruits size and weight; seed weight and number per fruit) and genetic descriptors (ribo-somic DNA extraction and PCR; nucleotide alignment and phylogenetic analysis) with standard protocols. Our results showed that the San Bernardo population had the heaviest fruits and seeds (7.89±0.2g and 0.19±0.002, respectively), and the Cebilar population the lightest (6.25±0.18g and 0.15±0.002g, respectively). Fruits and seeds from Metán and El Gallinato showed similar and intermediate values. The proportion viable (39 to 55%) and aborted (43 to 57%) seeds was different, while the proportion of predated (1.7 to 4.2%) seeds was similar among populations. The genetic analysis showed variability of ITS sequences within the especies, and also when compared with the same Brazilian species. Both, morphologic and genetic descriptors showed a high level of similarity between San Bernardo and Metán, and between El Cebilar and El Gallinato populations. Further studies are needed to assess levels of phenotypic and genetic variability within and between populations of different plant species, since this information is crucial for biodiversity and germplasm long-term conservation.
Key words: biodiversity, germplasm bank, molecular markers, nucleotidic sequence, plant conservation.
Resumen
Anadenanthera colubrina var. cebil es una especie arbórea de importancia cultural, económica y medicinal en Sur América. Para estudiar las poblaciones de A. colubrina, recolectamos frutos de cuatro sitios diferentes dentro del área de distribución de la especie en la provincia de Salta (Noroeste de Argentina) y se caracterizaron con base en descriptores morfológicos (tamaño de frutos, semillas y peso y número de semillas por fruto) y genéticos (ADN ribosómico). La población de San Bernardo presentó los frutos y semillas más pesados y la de El Cebilar los más livianos. Los frutos y semillas de Metán y El Gallinato fueron similares e intermedios. La proporción de semillas viables y abortadas fue similar en todas las poblaciones, mientras que la de semillas depredadas fue diferente. El análisis genético mostró variabilidad de las secuencias ITS dentro de la especie y también en comparación con la misma especie de Brasil. Los descriptores morfológicos y genéticos mostraron un mayor nivel de similitud entre las poblaciones de San Bernardo y Metán y entre El Cebilar y El Gallinato. Se necesitan más estudios para evaluar los niveles de variabilidad fenotípica y genética dentro y entre poblaciones de diferentes especies de plantas, ya que esta información es fundamental para la conservación de la biodiversidad y del germoplasma a largo plazo.
Palabras clave: biodiversidad, banco de germoplasma, marcadores moleculares, secuencia nucleotídica, conservación de especies vegetales.
Anadenanthera colubrina var. cebil (Vell.) Brenan belongs to the Fabaceae/Leguminosae family (Lewis, Schrire, MacKinder, & Lock, 2005), Mimosoideae subfamily. This tree species has economic, medicinal, and cultural applications in South America. It is widely distributed in Brazil, Paraguay, Bolivia and Argentina, where it is found in eleven provinces in the Northeast, Northwest and center, in mountain and transition forests and in Chaco Serrano (Siri Von Reis, 1964). It measures up to 35m in height and can reach 60-80cm in diameter. The wood contains tannins, is hard and resistant to termites and is used in construction and furniture, as poles and for firewood. In popular medicine, it is used to treat respiratory problems and inflammations. It is considered a sacred tree by local cultures. Its seeds have been used for over 3 000 years by shamans in rituals and popular medicine as they contain alkaloids derived from dimethyl tryptamine (Carod-Artal & Vásquez-Cabrera, 2007; Demaio, Karlin, & Medina, 2002; Jus-tiniano & Fredericksen, 1998; Martinez & Andrade, 2006; Monteiro et al., 2006). This species is categorized as “least concern” with low risk of extinction (IUCN, 2012).
The characterization of germplasm collections allows the assessment of the variability of the preserved material, although this information is poorly documented in seed banks. When the characterizations are morphological, descriptors that exhibit high heritability are used due to their stability in different environments (de Viana, Morandini, Giamminola, & Diaz, 2011; Souza & Sorrells, 1991).
Studies carried out on wild plant species have revealed that the distribution of morphological and chemical characteristics present patterns can be related to geographic regions (Nevo, Beiles, & Krugman, 1988; Nevo, Noy-Meir, Beiles, Krugman, & Agami, 1991). These variations may reflect the phenotypic plasticity of the individuals, genetic adaptations of the populations to different environments or both (Bradshaw, 1984; Matthies; Schmid, & Smid Hempel, 1995). For instance, Wulff (1986) reported that the degree of differentiation in populations of the genus Plantago was greater for morphological than for isoenzymatic markers, due to its great phenotypic plasticity in response to environmental differences. Makkar & Becker (1998) found that plants in semiarid environments contain more phenolic compounds than in humid African environments. Morphological differences were also reported in relation to genetic and environmental characteristics, such as soil moisture, nutrient levels, precipitation, radiation, temperature and elevation (Baker, 1972; Kuiper, 1985; Bond, Hoing, & Maze, 1999; Bu et al., 2007) and Monteiro et al (2006) reported a positive relationship between A. colubrina bark tannin content and environmental humidity.
Morphological characteristics are related to genetic traits, but this information is poorly documented. The genetic diversity of the Brazilian A. colubrina was studied using micro-satellite markers, and polymorphisms were found in the studied populations (Feres et al., 2012), as well as in A. colubrina var. cebil inArgentina (Barrandeguy, Prinz, García, & Fin-keldey, 2012).
Different molecular markers have been used to study plant genetic diversity, such as ISSR (Inter Simple Sequence Repeat) (Lombardo, Schicchi, Marino, & Palla, 2012). Other molecular markers of great importance are the Intergenic Transcripted Spacer (ITS) regions of ribosomal rRNA (rRNA). ITS refers to a piece of non-functional RNA situated between rRNAs. This polycistronic rRNA transcript contains the 5’ external transcribed sequence (ETS), 18S rRNA, ITS1, 5.8S rRNA, ITS2, 28S rRNA and finally the 3’ ETS. Genes encoding rRNA occur in tandem repeats that are in thousands of copies long, each separated by non-transcribed DNA regions termed intergenic spacers (IGS) or non-transcribed spacers (NTS) (Rogers & Bendich, 1987). They have an important biological meaning in rRNA processing and in RNAs during ribosome maturation (Hausner & Wang, 2005). In genomes, the ITS regions vary greatly in size and sequence (Korabecna, 2007).
Sequence comparison of the ITS region is widely used in taxonomy and molecular phylogeny. This can be explained by the relatively low evolutionary pressure acting on such non-functional sequences. For example, ITS has been proven especially useful for elucidating relationships among congeneric spe-cies and closely related genera in Asteraceae and Cyperaceae (Baldwin, 1992; Shekhovtsov, Shekhovtsova, & Peltec, 2012).
The aim of this work was to study the morphological descriptors and the genetic diversity of A. colubrina var. cebil, collected from different locations over its distribution area in Salta Province (North Argentina) and conserved in the Germplasm Bank of Native Species (BGEN-INEAH) of the National University of Salta by analyzing, for the first time, the ITS of rDNA region.
Materials and Methods
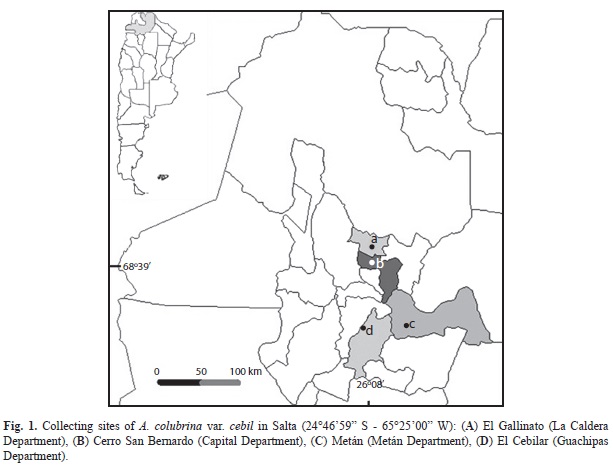
Collection of fruits: Initially four populations of A. colubrina var. cebil (Ac) wereselected from different sites over the distribution area of the species in Salta province: El Gallinato (G), San Bernardo (B), El Cebilar (C) and Metán (M) (Fig. 1), all with different elevation and rainfall (Table 1). From each population, we collected mature fruits from 10 trees (separated by at least 30m from each other). The fruits were processed in the INEAH laboratory, separating two groups: one for long term conservation in the seed bank, and the other for morphological and genetic characterization.
Morphological approach of fruits and seeds: Morphological characterization was performed by randomly sampling 75 fruits from each population. We recorded the weight (Denver Instrument APX analytical balance-200, 0.1mg accuracy), the length and the width (with digital gauge) of each fruit and counted the number of predated, aborted and plain seeds per fruit. The plain seeds (viable) were individually measured (width, length and thickness) and weighed. The weight distributions of fruits and seeds were analyzed with the Shapiro - Wilks normality test as well as skewness and kurtosis tests. The morphological descriptors were compared between populations with ANOVA and analysis of conglomerates (Euclidean distance) using InfoStat (2009).
Isolation and sequencing of ITS-rDNA regions: Genomic DNA was extracted from the seeds of one tree from each population following the protocol of Doyle & Doyle (1990). PCR was performed according to Esteve-Zarzoso, Belloch, Uruburu, and Querol (1999). Briefly, the primers ITS1 (5´-TCC-GTAGGTGAACCTTGCGG-3´) and ITS4 (5´-TCCTCCGCTTATTGATATGC -3´) were used for the PCR. The PCR mix contained 0.5μmol/L of each primer, 10μmol/L deoxy-nucleotides, 1.5mmol/L MgCl2 and 1×PCR buffer, 1 unit of Taq enzyme (AmpliTaq Gold® PCR Master Mix). The suspension was heated at 95°C for 5min in a PE 2400 thermocycler (Applied Biosystems, USA). PCR conditions were: 35 cycles of denaturing (94°C, 1min), annealing (55°C, 1min) and extension (72°C, 2min), followed by a final extension at 72°C for 10min. The fragments, ranging from 390-490nt. obtained by PCR, were purified by resin columns (Qiagen) and sent to sequencing to MWG company (Germany).
Nucleotide alignment and phylogenetic analysis: Multiple alignment of the ITS-rRNA region nucleotidic sequences was performed with Clustal W2 and Boxshade programs, manually refined (Thompson, Higgins, & Gib-son, 1994). A phylogenetic tree was generated by using the Gene Bee service, Tree-Top program choosing a Phylip format, with boot-strap values, in agreement with alignment (Saitou & Nei, 1987; Brodsky et al., 1995). Sequence accession numbers are AcBr (Brazilian): DQ787408; AcM: JQ910930; AcB: HF564640; AcG: HF564641; AcC: HF564642.
Results
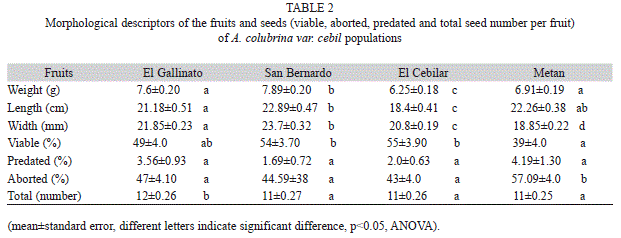
Morphological approach of fruits and seeds: The weight distributions of fruits were normal in the four populations studied (SW, p>0.05). San Bernardo population presented the heaviest fruits, El Cebilar the lightest, and the fruits from Metán and El Gallinato were similar and intermediate in weight. The same trend was observed in the length of the fruits. The width was also highest in the San Bernardo fruits, lowest in Metán, and intermediate in El Gallinato and El Cebilar. Seed num-ber per fruit was significantly higher in the El Gallinato population.
In the four populations, the proportion of plain seeds was low and ranged from 39% (Metán) to 55% (El Cebilar). The proportion of aborted seeds was high in all populations, although Metán had the highest level. The pro-portion of predated seeds was low and similar among populations (Table 2).
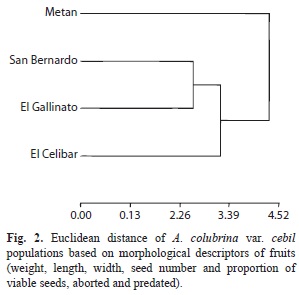
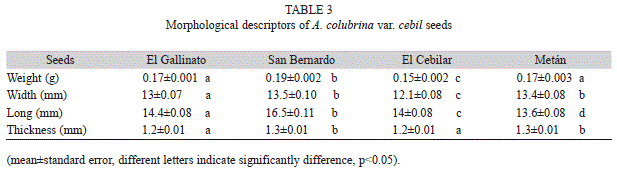
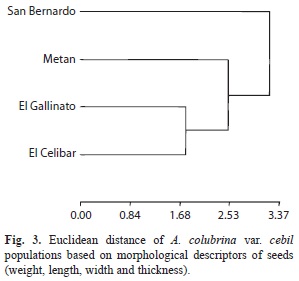
The analysis of the morphological traits of the fruits showed that the most distant populations were Metán and El Cebilar (Euclidean distance 4.49), whereas San Bernardo and El Gallinato were closer. In addition, these two populations were more distant from Metán than from El Cebilar (Fig. 2). The distributions of seed weights were normal in El Cebilar and El Gallinato, whereas in San Bernardo and Metán, seed weight was skewed and negative leptokurtic. Seed weight varied between populations and was significantly higher in San Bernardo, lower in El Cebilar and inter-mediate in El Gallinato and Metán, similar to the reported distributions in fruits. The other morphological descriptors (width, length and thickness) varied between populations. Overall, the San Bernardo population had the largest seeds (Table 3). The analysis of seed morphological descriptors showed that the more distant populations were San Bernardo and El Cebilar (Euclidean distance 4.09), and the closest were El Gallinato and El Cebilar. In addition, these two populations were more distant from San Bernardo than from Metán (Fig. 3).
Genetic approach of A. colubrina var. cebil seeds: We isolated, for the first time, the ITS of rDNA from fresh seeds of four individual A. colubrina var. cebil trees (one from each population) from Salta Province. They were long: 1 067, 960, 861, 580 base pairs, and were registered in GenBank. The sizes of the specificparts in the isolated regions of the individual trees were: ITS1=158bp; 5.8SrRNA=166bp; ITS2=170bp. We selected a restricted region of ITS, equal to San Bernardo (AcB) 450bp, El Cebilar (AcC) 448bp, El Gallinato (AcG) 448bp, Metán (AcM) 450bp, and aligned the nucleotidic sequences with a 397bp long sequence (AcBr, Brazilian), the only one found in Genbank, which was isolated from A. colubrina in Brazil. The restricted regions contain the following parts: from nucleotide 1 to 95, the partial “ITS1”; from 96 to 261, the complete gene “5.8S rRNA”; from 262 to 450, the partial “ITS2”. The nucleotidic sequence comparison showed a high identity between San Bernardo (AcB) and Metán (AcM) with a similarity of 98.1%, as well as between the El Cebilar (AcC) and the El Gallinato (AcG) sequences. The identity decreased to 90.4% when the AcB sequence was compared with AcC and AcG. The Brazilian sequence showed a lower identi-ty, ranging from 83.6%, if compared with AcC and AcG, to 84.1% with AcB and 84.6% with AcM. Moreover, the restriction analysis data showed variability in ITS region; for example three SmaI sites in the Metán and San Bernardo sequences were present opposite to those of the El Cebilar, El Gallinato and Brazilian sequences which had only one SmaI site. The variability is higher in the Brazilian sequence.
Then, we investigated the relationships among the ITS-rDNA regions of different individuals from the same species of A. colubrina var. cebil. The nucleotidic alignment showed a very high conservation of the nucleotidic sequence among the different individuals (Fig. 4A).
Nevertheless, some differences were evident: transitions (among purine or among pyrimidines) and transversions (from purine to pyrimidines and vice versa). Moreover there was a deletion of 12 nucleotides at the beginning of the ITS2 region in the four sequences from Argentina. Nine nucleotidic changes were present in all the analyzed accessions, while there were other changes in two out of four individuals (Fig. 4A). These differences can be attributed to intraspecific polymorphism, perhaps due to the different environments in which the trees grew.
Figure 4B shows a graphic phylogram, with cluster algorithm and bootstrap values. In this phylogenetic tree, it is clear that Argentin-ian and Brazilian ITS genes form different branches; then, the Argentinian branch divides into two: one for AcB and AcM, and another giving rise to AcC and AcG. The Brazilian A. colubrina is an outgroup. The high bootstrap values (100), indicated the accuracy of the data. Another difference, regarding the GC content, is that Argentinian A. colubrina var. cebil has a similar percentage (ranging from 59 to 60%), while the Brazilian one presents a higher GC content (70%).
Discussion
The studies of phenotypic and genetic diversity, as well as the detection of local varieties adjusted to different environments, are crucial issues in long-term (in situ and ex situ) biodiversity conservation. They are also impor-tant for the understanding of genetic change, adaptation or speciation in plant populations (Arzate-Fernández et al., 2005).
Although theoretical models suggest that balancing selection should be heavy on features related to biological fitness (McGinley, Temme, & Geber, 1987), numerous plant species have documented wide variations in the morphology of fruits and seeds also related to geographic variability (Baker, 1972; Busso & Perryman, 2005; Guo, Mazer, & Guozhen, 2010).
In this work, we found variations in the morphological traits of fruits and seeds in the four studied populations of Argentinian A. colubrina var. cebil. The variation was recorded within populations, and it was higher in fruits than in seeds. In short, for fruits descriptors, we found a greater variability between Metán and the other populations, whereas those of San Bernardo and El Gallinato were more similar. Instead, the distance of seed morphological descriptors was highest between the San Bernardo and El Cebilar populations, while El Gallinato and El Cebilar were more similar.
Regarding the genetic characterization, ITS sequences are used for phylogenetic studies due to their ease of use. In fact, first they are subjected to evolution, and then they can be easily cloned, even from preserved material.
It is possible to use gene trees to obtain information about species history. It is known that ITS-rDNA genes are good species evolution markers because they contain both conserved and variable regions. They are present and have similar functions in all organisms and are easy to clone (Ribeiro, Rapini, Silva, & Berg, 2012). We used this genetic analysis as a complement to the phenotypic descriptors of A. colubrina var. cebil because of the few individual trees studied.
ITS region is a high variable region, so it is a particularly valuable resource for plant systematists. In fact, ITS based studies improved our knowledge on plant phylogeny providing direct evidence of species relationships, such as calcydenia or glycine (Baldwin, 1992, 1993; Baldwin et al., 1995; Kollipara, Singh, & Hymowitz, 1997). Also in other species such as fungi, as an example, high variability in ITS region allowed to better understand the infraspecific variability (Simon & Weiß, 2008; Schoch et al., 2012). Althouhgt different markers are used to understand the genetic differentiation and relationships in various related species such as inter simple sequence repeats (ISSR) (Barrandeguy et al., 2012), we used ITS region, even if we think that could be important to do a comparison among different markers.
In the present study, we used ITS-rDNA sequences to address one main objective: to reconstruct the phylogenetic relationships among the four individuals of A. colubrina var.cebil from Salta province. We isolated the ITS-rDNA from A. colubrina var. cebil, and we found variability within the species. The highest similarity of nucleotidic sequence was found between San Bernardo and Metán, and between El Cebilar and El Gallinato, as demonstrated by phylogenetic analysis. This is in agreement with our analysis of seed morphological descriptors. The only sequence recorded for the same species, from a plant in Brazil, presented a lower similarity to all Argentinian sequences, suggesting a divergent evolution. Furthermore, GC content was found to be different from the Argentinian species when compared to the Brazilian one. In fact, the GC content is a variability parameter that can contribute to variation in selection, mutational proneness and DNA repair.
Even though the present genetic study is limited solely to A. colubrina var. cebil species within a radius of 100km in the Salta province of Northern Argentina, we have demonstrated a morphological and genetic intraspecific variability. It is perhaps due to geographic characteristics of the areas in which the tree live, as described by Ellison, Buckley, Miller, and Gotelli (2004) on the plant Sarracenia purpurea. Our results agree with the concept that there is great plant phenotypic plasticity that allows the assessment of diversity in response to the environmental variations (Mondini, Noorani, & Pagnotta, 2009). Further studies are needed to explore this topic, through the investigation of a large number of individuals and expanding the target zones in order to determine the evolution of the A. colubrina var. cebil species.
In this study, we concluded that the Argentinian tree A. colubrina var. cebil shows significant morphological and genetic variability and that ITS-rDNA molecular markers are useful tools for detecting genetic variability in plants.
The documentation and recording of phenotypic and genetic variability is crucial for the maintaining of the long-term conservation of biodiversity and for germplasm conservation.
In order to preserve the greatest possible diversity (morphological and genetic) of the accessions, with ex situ conservation purposes, it is important to collect seeds from many populations, especially for widespread species that grow in a variety of environments, such as A. colubrina var. cebil. However, further studies are needed to assess levels of variability within and between populations. A. colubrina var. cebil is an important plant species because of its ornamental, medicinal, cultural, ecological and economic uses (wood, honey, alkaloids, tannin and gum extraction, restoration). These reasons point to the need for A. colubrina var. cebil conservation and propagation.
Acknowledgments
We thank the Research Council of the National University of Salta and INEAH for supporting this work and Maria Emilia Muñoz, Griselda Salas Barboza and Valeria Pastrana Ignes for the collaboration with the morphological characterization. We acknowledge Franco Palla for critical reading of the manuscript and for very helpful suggestions.
References
Arzate-Fernández, A. M., Miwa, M., Shimada, T., Yonekura, T., & Ogawa, K. (2005). Genetic diversity of Miyamasukashi-yuri (Lilium maculatum Thunb. var. bukosanense), an endemic and endangered species at Mount Buko, Saitama, Japan. Plant Species Biology, 20, 57-65. [ Links ]
Baker, G. (1972). Seed weight in relation to environmental conditions in California. Ecology, 53, 997-1010. [ Links ]
Baldwin, B. G. (1992). Phylogenetic utility of the internal transcribed spacers of nuclear ribosomal DNA in plants: An example from the Compositae. Molecular Phylogenetics and Evolution, 1, 3-16. [ Links ]
Baldwin, B. G. (1993). Molecular phylogenetics of Calcydenia (Compositae) based on ITS sequences of nuclear ribosomal DNA: Chromosomal and morphological evolution reexamined. American Journal of Botanic, 80, 222-238. [ Links ]
Baldwin, B. G., Sanderson, M. J., Porter, J. M., Wojciechowski, M. F., Campbell, C. S., & Donoghue, M. J. (1995). The ITS region of nuclear ribosomal DNA: a valuable source of evidence on angiosperm phylogeny. Annals of the Missouri Botanical Garden, 82, 247-277. [ Links ]
Barrandeguy, M. E., Prinz, K., García, M. V., & Finkeldey, R. (2012). Development of microsatellite markers for Anadenanthera colubrina var. cebil (fabaceae), a native tree from South America. American Journal of Botany, 99, e372-e374. [ Links ]
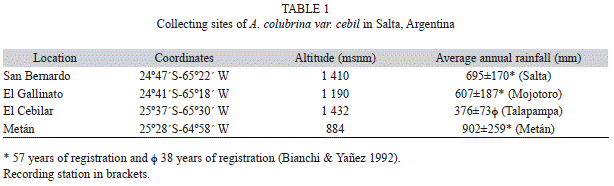
Bianchi, A. R., & Yanez, Y. C. (1992). Las precipitaciones en el Noroeste Argentino. Instituto Nacional de Tecnología Agropecuaria. Estación Experimental Agropecuaria. Salta: INTA. [ Links ]
Bond, W. J., Hoing, M., & Maze, K. E. (1999). Seed size and seedling emergence: an allometric relationship and some ecological implications. Oecologia, 120, 120-132. [ Links ]
Bradshaw, A. D. (1984). Ecological significance of genetic variation between populations. In R. Dirzo, & J. Sarukhan (Eds.), Perspectives on plant population ecology (pp. 213-228). Sunderland: Sinauer Associates Inc. Publishers. [ Links ]
Brodsky, L. I., Ivanov, V. V., Kalaidzidis, Y. L., Leontovich, A. M., Nikolaev, V. K., Feranchuk, S. I., & Drachev, V. A. (1995). GeneBee-NET: Internet-based server for analyzing biopolymers structure. Biochemistry, 60, 923-928. [ Links ]
Bu, H., Chen, X., Xu, X., Liu, K., Jia, P., & Du, G. (2007). Seed mass and germination in an alpine meadow on the eastern Tsinghai Tibet Plateau. Plant Ecology, 191, 127-149. [ Links ]
Busso, C. A., & Perryman, B. L. (2005). Seed weight variation of Wyoming sagebrush in Northern Nevada. Biocell, 29, 279-285. [ Links ]
Carod-Artal, F., & Vásquez-Cabrera, A. C. (2007). Usos rituales de la semilla de Anadenanthera sp. entre los indígenas sudamericanos. Neurología, 22, 410-415. [ Links ]
Demaio, P., Karlin, U. O., & Medina, M. (2002). Árboles nativos del centro de Argentina. Córdoba: L.O.L.A Editorial. [ Links ]
de Viana, M. L., Morandini, M. N., Giamminola, E. M., & Diaz, R. (2011). Conservación ex situ: un banco de germoplasma de especies nativas. Lhawet, 1, 35-41. [ Links ]
Doyle, J. J., & Doyle, J. L. (1990). Isolation of plant DNA from fresh tissue. Focus, 12, 13-15. [ Links ]
Ellison, A. M., Buckley, H. L., Miller, T. E., & Gotelli, N. J. (2004). Morphological variation in Sarracenia purpurea (Sarraceniaceae): geographic, environmental, and taxonomic correlates. American Journal of Botany, 91, 1930-1935. [ Links ]
Esteve-Zarzoso, B., Belloch, C., Uruburu, F., & Querol, A. (1999). Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. International Journal of Syste-matic Bacteriology, 49, 329-337. [ Links ]
Feres, J. M., Monteiro, M., Zucchi, M. I., Pinheiro, J. B., Mestrineir, M. A., & Alzate Marin, A. L. (2012). Development of microsatellite markers for Anadenanthera colubrina (leguminosae), a neotropical tree species. American Journal of Botany, 99, e154-e156. [ Links ]
Guo, H., Mazer, S. J., & Guozhen, D. (2010). Geographic variation in seed mass within and among nine species of Pedicularis (Orobanchaceae): effects of elevation, plant size and seed number per fruit. Journal of Ecology, 98, 1232-1242. [ Links ]
Hausner, G., & Wang, X. (2005). Unusual compact rDNA gene arrangements within some members of the Ascomycota: evidence for molecular co-evolution between ITS1 and ITS2. Genome, 48, 648. [ Links ]
INFOSTAT versión. (2009). Grupo InfoStat, FCA. Universidad Nacional de Córdoba: Editorial Brujas. [ Links ]
IUCN. (2012). Red List of Threatened Species. Retrieved from http://www.iucnredlist.org. [ Links ]
Justiniano, M. J., & Fredericksen, T. S. (1998). Ecología y Silvicultura de Especies Menos Conocidas-Curupaú Anadenanthera colubrina (Vell.Conc.) Benth, Mimo-soideae. Santa Cruz: BOLFOR. [ Links ]
Kollipara, K. P., Singh, R. J., & Hymowitz, T. (1997). Phylogenetic and genomic relationship in the genus Glycine Wild. Based on sequences from the ITS region rDNA. Genome, 40, 57-68. [ Links ]
Korabecna, M. (2007). The Variability in the Fungal Ribo-somal DNA (ITS1, ITS2, and 5.8 S rRNA Gene): Its Biological Meaning and Application in Medical Mycology. Communicating Current Research and Educational Topics and Trends in Applied Microbio-logy, 105, 783-787. [ Links ]
Kuiper, D. (1985). Genetic differentiation and phenotypic plasticity in populations of Plantago lanceolata in response to nutrient levels. In P. Jacquard, G. Heim, & J. Antonovics (Eds.), Genetic differentiation and dispersal in plants (pp. 251-269). Berlin: Springer. [ Links ]
Lewis, G., Schrire, B., MacKinder, B., & Lock, M. (Eds.). (2005). Legumes of the world. UK: Royal Botanical Gardens, Kew. [ Links ]
Lombardo, G., Schicchi, R., Marino, P., & Palla, F. (2012). Genetic analysis of Citrus aurantiu L. (Rutaceae) cultivars by ISSR molecular markers. Plant Biosys-tems, 146, 19-26. [ Links ]
Makkar, H. P. S., & Becker, K. (1998). Jatropha curcas toxicity: identification of toxic principle. In T. Garland & A. C. Barr (Eds.), Toxic plants and other natural toxicants (pp. 554-558). New York: CAB International. [ Links ]
Martinez, S. M., & Andrade, D. J. (2006). Guía de árboles nativos de la Provincia de Salta, Noroeste Argentino. Ministerio de Educación de la Provincia de Salta, Secretaría de Cultura. [ Links ]
Matthies, D., Schmid, B., & Schmid-Hempel, P. (1995). The importance of population processes for the maintenance of biological diversity, Gaia, 4, 199-209. [ Links ]
McGinley, M., Temme, D., & Geber, M. (1987). Parental investment in off springs in variable environments, theoretical and empirical considerations. The American Naturalist, 130, 370-398. [ Links ]
Mondini, L., Noorani, A., & Pagnotta, M. A. (2009). Review Assessing Plant Genetic Diversity by Molecular Tools. Diversity, 1, 19-35. [ Links ]
Monteiro, J. M., Albuquerque, U. P., Linsneto, E. M. F., Araujo, E. L., Albuquerque, M. M., & Amorim, E. L. C. (2006). The effects of seasonal climate changes in the caatinga on tannin levels in Myracrodruon urundeuva (Engl.) Fr. All. and Anadenanthera colubrina (Vell.) Brenan. Revista Brasileira de Farmacognosia, 16, 338-344. [ Links ]
Nevo, E., Beiles, A., & Krugman, T. (1988). Natural selection of allozyme polymorphims: a microgeographical differentiation by edaphic, topographical, and temporal factors in wild emmer wheat (Triticum dicoccoides). Theoretical and Applied Genetics, 76, 737-752. [ Links ]
Nevo, E., Noy-Meir, I., Beiles, A., Krugman, T., & Agami, M. (1991). Natural selection of allozyme polymorphisms: microgeographical spatial and temporal ecological differentiations in wild emmer wheat. Israel Journal of Botany, 40, 419-449. [ Links ]
Ribeiro, P. L., Rapini, A., Silva, U. C., & Berg, C. (2012). Using multiple analytical methods to improve phylogenetic hypotheses in Minaria (Apocynaceae). Molecular Phylogenetics and Evolution, 65, 915-925. [ Links ]
Rogers, S. O., & Bendich, A. J. (1987). Ribosomal RNA genes in plants: variability in copy number and in the intergenic spacer. Plant Molecular Biology, 9, 509-520. [ Links ]
Saitou, N., & Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4: 406-425. [ Links ]
Schoch, C. L., Seifert, K. A., Huhndorf, S., Robert, V., Spouge, J. L., Levesque, C. A., Chen, W., & Fun-gal Barcoding Consortium. (2012). Nuclear ribosomal internal transcribed spacer (ITS) region as universal DNA barcode marker for Fungi. PNAS, 109, 6241-6246. [ Links ]
Shekhovtsov, S. V., Shekhovtsova, I. N., & Peltek, S. E. (2012). Phylogeny of Siberian species of Carex sect. Vesicariae based on nuclear and plastid markers. Nordic Journal of Botany, 30, 343-351. [ Links ]
Simon, U. K., & Weiß, M. (2008). Intragenomic variation of fungal ribosomal genes is higher than previously thought. Molecular Biology and Evolution, 25, 2251-2254. [ Links ]
Siri von Reis, A. (1964). Taxonomic Study of Anadenanthera. Contributions from the Gray Herbarium of Harvard University, 193, 3-65. [ Links ]
Souza, E., & Sorrells, M. E. (1991). Relationships among 70 North American oat germplasms: II. Cluster analysis using qualitative characters. Crop Science, 31, 505-612. [ Links ]
Thompson, J. D., Higgins, D. G., & Gibson, T. J. (1994). Clustal W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22, 4673-4680. [ Links ]
Wulff, R. D. (1986). Seed size variation in Desmodium paniculatum. I Factors affecting seed size. Journal of Ecology, 74, 87-97. [ Links ]
Baker, G. (1972). Seed weight in relation to environmental conditions in California. Ecology, 53, 997-1010. [ Links ]
Baldwin, B. G. (1992). Phylogenetic utility of the internal transcribed spacers of nuclear ribosomal DNA in plants: An example from the Compositae. Molecular Phylogenetics and Evolution, 1, 3-16. [ Links ]
Baldwin, B. G. (1993). Molecular phylogenetics of Calcydenia (Compositae) based on ITS sequences of nuclear ribosomal DNA: Chromosomal and morphological evolution reexamined. American Journal of Botanic, 80, 222-238. [ Links ]
Baldwin, B. G., Sanderson, M. J., Porter, J. M., Wojciechowski, M. F., Campbell, C. S., & Donoghue, M. J. (1995). The ITS region of nuclear ribosomal DNA: a valuable source of evidence on angiosperm phylogeny. Annals of the Missouri Botanical Garden, 82, 247-277. [ Links ]
Barrandeguy, M. E., Prinz, K., García, M. V., & Finkeldey, R. (2012). Development of microsatellite markers for Anadenanthera colubrina var. cebil (fabaceae), a native tree from South America. American Journal of Botany, 99, e372-e374. [ Links ]
Bianchi, A. R., & Yanez, Y. C. (1992). Las precipitaciones en el Noroeste Argentino. Instituto Nacional de Tecnología Agropecuaria. Estación Experimental Agropecuaria. Salta: INTA. [ Links ]
Bond, W. J., Hoing, M., & Maze, K. E. (1999). Seed size and seedling emergence: an allometric relationship and some ecological implications. Oecologia, 120, 120-132. [ Links ]
Bradshaw, A. D. (1984). Ecological significance of genetic variation between populations. In R. Dirzo, & J. Sarukhan (Eds.), Perspectives on plant population ecology (pp. 213-228). Sunderland: Sinauer Associates Inc. Publishers. [ Links ]
Brodsky, L. I., Ivanov, V. V., Kalaidzidis, Y. L., Leontovich, A. M., Nikolaev, V. K., Feranchuk, S. I., & Drachev, V. A. (1995). GeneBee-NET: Internet-based server for analyzing biopolymers structure. Biochemistry, 60, 923-928. [ Links ]
Bu, H., Chen, X., Xu, X., Liu, K., Jia, P., & Du, G. (2007). Seed mass and germination in an alpine meadow on the eastern Tsinghai Tibet Plateau. Plant Ecology, 191, 127-149. [ Links ]
Busso, C. A., & Perryman, B. L. (2005). Seed weight variation of Wyoming sagebrush in Northern Nevada. Biocell, 29, 279-285. [ Links ]
Carod-Artal, F., & Vásquez-Cabrera, A. C. (2007). Usos rituales de la semilla de Anadenanthera sp. entre los indígenas sudamericanos. Neurología, 22, 410-415. [ Links ]
Demaio, P., Karlin, U. O., & Medina, M. (2002). Árboles nativos del centro de Argentina. Córdoba: L.O.L.A Editorial. [ Links ]
de Viana, M. L., Morandini, M. N., Giamminola, E. M., & Diaz, R. (2011). Conservación ex situ: un banco de germoplasma de especies nativas. Lhawet, 1, 35-41. [ Links ]
Doyle, J. J., & Doyle, J. L. (1990). Isolation of plant DNA from fresh tissue. Focus, 12, 13-15. [ Links ]
Ellison, A. M., Buckley, H. L., Miller, T. E., & Gotelli, N. J. (2004). Morphological variation in Sarracenia purpurea (Sarraceniaceae): geographic, environmental, and taxonomic correlates. American Journal of Botany, 91, 1930-1935. [ Links ]
Esteve-Zarzoso, B., Belloch, C., Uruburu, F., & Querol, A. (1999). Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. International Journal of Syste-matic Bacteriology, 49, 329-337. [ Links ]
Feres, J. M., Monteiro, M., Zucchi, M. I., Pinheiro, J. B., Mestrineir, M. A., & Alzate Marin, A. L. (2012). Development of microsatellite markers for Anadenanthera colubrina (leguminosae), a neotropical tree species. American Journal of Botany, 99, e154-e156. [ Links ]
Guo, H., Mazer, S. J., & Guozhen, D. (2010). Geographic variation in seed mass within and among nine species of Pedicularis (Orobanchaceae): effects of elevation, plant size and seed number per fruit. Journal of Ecology, 98, 1232-1242. [ Links ]
Hausner, G., & Wang, X. (2005). Unusual compact rDNA gene arrangements within some members of the Ascomycota: evidence for molecular co-evolution between ITS1 and ITS2. Genome, 48, 648. [ Links ]
INFOSTAT versión. (2009). Grupo InfoStat, FCA. Universidad Nacional de Córdoba: Editorial Brujas. [ Links ]
IUCN. (2012). Red List of Threatened Species. Retrieved from http://www.iucnredlist.org. [ Links ]
Justiniano, M. J., & Fredericksen, T. S. (1998). Ecología y Silvicultura de Especies Menos Conocidas-Curupaú Anadenanthera colubrina (Vell.Conc.) Benth, Mimo-soideae. Santa Cruz: BOLFOR. [ Links ]
Kollipara, K. P., Singh, R. J., & Hymowitz, T. (1997). Phylogenetic and genomic relationship in the genus Glycine Wild. Based on sequences from the ITS region rDNA. Genome, 40, 57-68. [ Links ]
Korabecna, M. (2007). The Variability in the Fungal Ribo-somal DNA (ITS1, ITS2, and 5.8 S rRNA Gene): Its Biological Meaning and Application in Medical Mycology. Communicating Current Research and Educational Topics and Trends in Applied Microbio-logy, 105, 783-787. [ Links ]
Kuiper, D. (1985). Genetic differentiation and phenotypic plasticity in populations of Plantago lanceolata in response to nutrient levels. In P. Jacquard, G. Heim, & J. Antonovics (Eds.), Genetic differentiation and dispersal in plants (pp. 251-269). Berlin: Springer. [ Links ]
Lewis, G., Schrire, B., MacKinder, B., & Lock, M. (Eds.). (2005). Legumes of the world. UK: Royal Botanical Gardens, Kew. [ Links ]
Lombardo, G., Schicchi, R., Marino, P., & Palla, F. (2012). Genetic analysis of Citrus aurantiu L. (Rutaceae) cultivars by ISSR molecular markers. Plant Biosys-tems, 146, 19-26. [ Links ]
Makkar, H. P. S., & Becker, K. (1998). Jatropha curcas toxicity: identification of toxic principle. In T. Garland & A. C. Barr (Eds.), Toxic plants and other natural toxicants (pp. 554-558). New York: CAB International. [ Links ]
Martinez, S. M., & Andrade, D. J. (2006). Guía de árboles nativos de la Provincia de Salta, Noroeste Argentino. Ministerio de Educación de la Provincia de Salta, Secretaría de Cultura. [ Links ]
Matthies, D., Schmid, B., & Schmid-Hempel, P. (1995). The importance of population processes for the maintenance of biological diversity, Gaia, 4, 199-209. [ Links ]
McGinley, M., Temme, D., & Geber, M. (1987). Parental investment in off springs in variable environments, theoretical and empirical considerations. The American Naturalist, 130, 370-398. [ Links ]
Mondini, L., Noorani, A., & Pagnotta, M. A. (2009). Review Assessing Plant Genetic Diversity by Molecular Tools. Diversity, 1, 19-35. [ Links ]
Monteiro, J. M., Albuquerque, U. P., Linsneto, E. M. F., Araujo, E. L., Albuquerque, M. M., & Amorim, E. L. C. (2006). The effects of seasonal climate changes in the caatinga on tannin levels in Myracrodruon urundeuva (Engl.) Fr. All. and Anadenanthera colubrina (Vell.) Brenan. Revista Brasileira de Farmacognosia, 16, 338-344. [ Links ]
Nevo, E., Beiles, A., & Krugman, T. (1988). Natural selection of allozyme polymorphims: a microgeographical differentiation by edaphic, topographical, and temporal factors in wild emmer wheat (Triticum dicoccoides). Theoretical and Applied Genetics, 76, 737-752. [ Links ]
Nevo, E., Noy-Meir, I., Beiles, A., Krugman, T., & Agami, M. (1991). Natural selection of allozyme polymorphisms: microgeographical spatial and temporal ecological differentiations in wild emmer wheat. Israel Journal of Botany, 40, 419-449. [ Links ]
Ribeiro, P. L., Rapini, A., Silva, U. C., & Berg, C. (2012). Using multiple analytical methods to improve phylogenetic hypotheses in Minaria (Apocynaceae). Molecular Phylogenetics and Evolution, 65, 915-925. [ Links ]
Rogers, S. O., & Bendich, A. J. (1987). Ribosomal RNA genes in plants: variability in copy number and in the intergenic spacer. Plant Molecular Biology, 9, 509-520. [ Links ]
Saitou, N., & Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4: 406-425. [ Links ]
Schoch, C. L., Seifert, K. A., Huhndorf, S., Robert, V., Spouge, J. L., Levesque, C. A., Chen, W., & Fun-gal Barcoding Consortium. (2012). Nuclear ribosomal internal transcribed spacer (ITS) region as universal DNA barcode marker for Fungi. PNAS, 109, 6241-6246. [ Links ]
Shekhovtsov, S. V., Shekhovtsova, I. N., & Peltek, S. E. (2012). Phylogeny of Siberian species of Carex sect. Vesicariae based on nuclear and plastid markers. Nordic Journal of Botany, 30, 343-351. [ Links ]
Simon, U. K., & Weiß, M. (2008). Intragenomic variation of fungal ribosomal genes is higher than previously thought. Molecular Biology and Evolution, 25, 2251-2254. [ Links ]
Siri von Reis, A. (1964). Taxonomic Study of Anadenanthera. Contributions from the Gray Herbarium of Harvard University, 193, 3-65. [ Links ]
Souza, E., & Sorrells, M. E. (1991). Relationships among 70 North American oat germplasms: II. Cluster analysis using qualitative characters. Crop Science, 31, 505-612. [ Links ]
Thompson, J. D., Higgins, D. G., & Gibson, T. J. (1994). Clustal W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22, 4673-4680. [ Links ]
Wulff, R. D. (1986). Seed size variation in Desmodium paniculatum. I Factors affecting seed size. Journal of Ecology, 74, 87-97. [ Links ]
*Correspondencia: Marta L. de Viana: Banco de Germoplasma de Especies Nativas, Instituto de Ecología y Ambiente Humano. Universidad Nacional de Salta. Avda. Bolivia 5150, 4400, Salta, Argentina; mldeviana@yahoo.com.ar
Eugenia Giamminola: Banco de Germoplasma de Especies Nativas, Instituto de Ecología y Ambiente Humano. Universidad Nacional de Salta. Avda. Bolivia 5150, 4400, Salta, Argentina; eugeniagiamminola@yahoo.com.ar
Roberta Russo: Istituto di Biomedicina e Immunologia Molecolare “A. Monroy”. Consiglio Nazionale delle Ricerche (CNR), Via Ugo La Malfa 153, 90146 Palermo, Italy; rrusso@ibim.cnr.it
Mirella Ciaccio: Istituto di Biomedicina e Immunologia Molecolare “A. Monroy”. Consiglio Nazionale delle Ricerche (CNR), Via Ugo La Malfa 153, 90146 Palermo, Italy; ciaccio@ibim.cnr.it
1. Banco de Germoplasma de Especies Nativas, Instituto de Ecología y Ambiente Humano. Universidad Nacional de Salta. Avda. Bolivia 5150, 4400, Salta, Argentina; mldeviana@yahoo.com.ar, eugeniagiamminola@yahoo.com.ar
2. Istituto di Biomedicina e Immunologia Molecolare “A. Monroy”. Consiglio Nazionale delle Ricerche (CNR), Via Ugo La Malfa 153, 90146 Palermo, Italy; ciaccio@ibim.cnr.it, rrusso@ibim.cnr.it
Received 13-vi-2013. Corrected 20-X-2013. Accepted 22-Xi-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}