











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.62 no.2 San José abr./jun. 2014
Los cromosomas de las tortugas tropicales: Kinosternon leucostomum, Trachemys scripta y Staurotypus triporcatus (Testudines: Kinosternidae/Emydidae)
Tropical turtles chrosomes: Kinosternon leucostomum, Trachemys scripta y Staurotypus triporcatus (Testudines: Kinosternidae/Emydidae)
Tropical turtles chrosomes: Kinosternon leucostomum, Trachemys scripta y Staurotypus triporcatus (Testudines: Kinosternidae/Emydidae)
*Dirección para correspondencia:
Abstract
Mexico is a biodiverse country in several taxa as reptiles, that include several species of freshwater and marine turtles. Eventhough most of this group species are under protection, Tabasco State has nine native freshwater turtles, like Kinosternon leucostomum, Trachemys scripta and Staurotypus triporcatus that are very important in traditional dishes. This has resulted in a critical level of their populations, together with little biological knowledge for their conservation. Therefore, this study was dedicated to turtle cytogenetics. The study was conducted using the conventional methods for cytogenetics. The results showed the modal diploid and haploid number for K. leucostomum of 2n=56 (2n=56+3 microchromosomes “B”) and 1n=28 chromosomes in mitosis and meiosis, respectively. In T. scripta 2n=50 chromosomes (2n=50+2 microchromosomes “B”) and 1n=25 chromosomes were also characterized. Whereas in S. triporcatus we only report the 2=54 chromosomes (2n=54+2 microchromosomes “B”). The karyological formula for K. leucostomum was integrated by 12 metacentric-submetacentric chromosomes “msm”/”A”+22 subtelocentrictelocentric chromosomes “stt”/”B”+22 telocentric chromosomes “T”/”C” with fundamental number (FN) of 90 chromosome arms. While T. scripta karyotype was integrated by 32 “msm/”A”+10 “stt”/”B”+8”T/”C” chromosomes, with FN of 92 arms. S. triporcatus karyotype formula was built up by 20 chromosomes “msm”/”A”+34 chromosomes “T”/”C” with FN of 74. The variation in chromosome classification, the fundamental number and the presence of supernumerary microchromosomes “B” in the studied species, were evidence of a particular chromosome cytotypes in Tabasco. We considered that the presence of microchromosomes “B” probably has different origins, and they may be very important as a pattern for the formation or separation of new species. This study also showed the absence of heterologous chromosomes between the females and males karyotypes from the studied species. Rev. Biol. Trop. 62 (2): 671-688. Epub 2014 June 01.
Key words: cytogenetic, karyotype, B microchromosomes, turtles, Tabasco.
Resumen
México es un país biodiverso en varios grupos taxonómicos incluyendo a los reptiles, por ello en el país existen varias especies de tortugas dulceacuícolas y marinas. Las especies que integran dicho grupo se encuentran dentro del listado de especies sujetas a protección. El estado de Tabasco cuenta con nueve especies de tortugas de agua dulce, de las cuales Kinosternon leucostomum, Trachemys scripta y Staurotypus triporcatus son de las más importantes dentro de la tradición culinaria, hecho que las ha llevado a niveles críticos en sus poblaciones; aunado al poco conocimiento biológico que sobre dichas especies existe para conservarlas. Por lo anterior, el presente estudio de citogenética es el primero en tortugas de agua dulce en la región. El estudio se realizó, empleando el método convencional de citogenética. Los resultados muestran, el número modal diploide y haploide de K. leucostomum de 2n=56 (2n=56+3 microcromosomas “B”) y 1n=28 cromosomas 686 en mitosis y meiosis, respectivamente. En T. scripta de 2n=50 cromosomas (2n=50+2 microcromosomas “B”) y 1n=25 cromosomas. Mientras que en S. triporcatus solo se reporta el 2n=54 cromosomas (2n=54+2 microcromosomas “B”). La fórmula cromosómica en K. leucostomum, fue de 12 cromosomas metacéntricos submetacéntricos “msm”/“A”+22 cromosomas subtelocéntricos-telocéntricos “stt”/“B”+22 cromosomas telocéntricos “T”/“C”, con número fundamental (NF) de 90 brazos cromosómicos. En T. scripta fue de 32 cromosomas “msm”/“A”+10 cromosomas “stt”/“B”+8 cromosomas “T”/“C”, con NF de 92 y en S. triporcatus 20 cromosomas “msm”/“A”+34 cromosomas “T”/“C” con NF de 74. La variación en la clasificación cromosómica, el número fundamental y la presencia de microcromosomas “B” supernumerarios en las tres especies, son evidencia de citotipos cromosómicos particulares de las tortugas de Tabasco. Se argumenta que la presencia de los microcromosomas “B” tiene diferentes orígenes y de su importancia como pauta para la formación o separación de nuevas especies. En el estudio se descarta la presencia de cromosomas heterólogos entre las hembras y los machos de las especies estudiadas.
Palabras clave: citogenética cariotipo, microcromosomas B, tortugas, Tabasco.
México es un país con amplia biodiversidad de plantas, animales, hongos y microorganismos (McCoy, 1982; Mittermeier & Goettsch, 1992). Sólo en el caso de los reptiles, ocupa el primer lugar mundial con 707 especies (Mittermeier & Goettsch, 1992); de ellas se han reportado que existen 177 especies de tortugas, distribuidas en 13 familias que habitan diversos ecosistemas entre ellos los acuáticos y terrestres (Lee, 2000). En el sureste de México, se localizan los estados de Veracruz, Oaxaca, Chiapas, Tabasco, Campeche, Yucatán y Quintana Roo, donde se ha reportado, con distribución común, la presencia de nueve especies de tortugas dulceacuícolas (West, Psuty & Thom, 1987; Lee, 2000).
Las tortugas de México particularmente en el sureste del país, son un grupo de reptiles aprovechados como alimento, mascotas y ornato de modo tradicional (West et al., 1987). Ello ha ocasionado, que la gran mayoría de las especies, se encuentran listadas en la NOM- 059 de la Secretaria de Medio Ambiente y Recursos Naturales en México (SEMARNAT, 2010) y en la lista roja de especies en peligro de extinción (IUCN, 2012). A pesar de dicha situación, los estudios sobre la biología básica de las tortugas tropicales (West et al., 1987; Lemos-Espinal & Smith, 2009), han abordado solo aquellos temas encaminados a la descripción geográfica, evaluación de la variación de los caracteres taxonómicos que hacen énfasis solo en la distribución espacial. Así como aquellas investigaciones relacionadas con los usos y costumbres en países como México, donde varias especies forman parte de las tradiciones arraigadas a varias culturas entre ellas la Olmeca, Maya y Tolteca (West et al., 1987). Por otro lado, estudios recientes de biología que permitan dar mayor comprensión a la ecología y diversidad genética de las especies, han sido limitados solo a especies de origen marino, dada su importancia compartida entre los países, donde varias especies se distribuyen y reproducen; no siendo dicho fenómeno el caso de las especies de agua dulce y terrestres (Reed et al., 1991; Camacho-Mosquera & Amorocho, 2008; López, Hernández Fernández, & Bernal-Villegas, 2008). Desde la perspectiva de la genética básica y molecular, se le ha dado mayor importancia a los anfibios (Hillis, 1991; Schmid et al., 2010), siendo limitados los estudios en el grupo de los reptiles dulceacuícolas y terrestres.
Al considerar lo anterior, se han realizado estudios de citogenética convencional en algunos miembros de la familia Kinosternidae por Bull, Moon & Legler, (1974), Killebrew (1975), Bickham & Baker (1976a); Sites, Bickham, Haiduk, & Iverson (1979a) y Bickham & Carr (1983). Al igual que en algunas especies de la familia Emydidae, principalmente aquellas del género Trachemys, representados por los trabajos de Cleiton & Giuliano-Caetano (2008), Martínez et al., (2009) y Kasai, O´Brien, Martin & Ferguson-Smith (2012). En México, los análisis cariotípicos en especies de quelonios nativos son escasos, ya que sólo existe el reporte para la tortuga dulceacuícola Dermatemys mawii (Carr, Bickham, & Dean, 1981). En los estudios de citogenética anteriores, subyace la problemática de que las descripciones de los cariotipos se han basado solo en parámetros citológicos altamente subjetivos y redundantes, debido a que estos solo expresan en sus resultados, el número de cromosomas en mitosis, sin señalar la variación intrínseca que existe debido a la alta incidencia de microcromosomas “B” de tipo monorrámeo. Mientras que desde la perspectiva morfológica, esta solo se ha abordado con base en el aspecto visual de cada par cromosómico, sin el empleo normativo de las medidas reales en micrómetros de las longitudes de los brazos cortos (p) y largos (q), que es de amplio uso en un sin número de vertebrados y plantas, que ha hecho de ellos comparables y sujetos al establecimiento de citotipos especiales (Levan, Fredga, & Sandberg, 1964; Denton 1973; Arai 2011). Por lo anterior, en este estudio se describen los números cromosómicos y la estructura cariotípica en mitosis y meiosis de tres especies de tortugas tropicales en riesgo crítico del estado de Tabasco en México, K. leucostomum, T. scripta y S. triporcatus.
Materiales y métodos
Sitios de recolecta, determinación taxonómica y mantenimiento de especímenes: La recolecta de las tortugas, K. leucostomum (cinco machos y cinco hembras con longitud y peso promedio de 12.4±1.52cm y 273.3±68.2g), T. scripta (cinco machos y cinco hembras con longitud y peso promedio de 13.3±2.54cm y 217.2±13.8g) y S. triporcatus (cuatro machos y seis hembras con 15.8±3.8cm y 495.5±16.2g en promedio), se llevó a cabo de modo manual. Los especímenes fueron extraídos de las orillas de la laguna del Pueblo de la Villa Luis Gil Pérez, la zona de humedales de los alrededores de la División Académica de Ciencias Biológicas-UJAT ambos del municipio del Centro, y de los humedales de la Villa Benito Juárez del municipio de Cárdenas, en todos los casos del estado de Tabasco. Los especímenes adultos, fueron identificados basados en los caracteres merísticos y morfométricos recomendados por Lee (2000). Los individuos recolectados, fueron mantenidos por tres meses en tanques de fibra de vidrio bajo las condiciones necesarias de alimentación y calidad de agua.
Procedimiento citológico, análisis microscópico y armado de cariotipos: Los especímenes fueron procesados con el protocolo propuesto por Arias-Rodriguez, Ibarra-Castro & Páramo-Delgadillo (2008); Arias-Rodriguez, Páramo-Delgadillo, Contreras- Sánchez & Álvarez-González (2009) y Hernández-Guzmán, Arias-Rodriguez & Indy (2011): se inyectó intraperitonealmente 50μg/g de colchicina diluida en citrato de sodio al 0.1%, a cada espécimen con seis horas de exposición. Posteriormente, fueron sacrificados por hipotermia y luego se removieron los tejidos (páncreas, riñones y gónadas) para la hidratación en 1mL de citrato de sodio al 2%, con lo cual fueron incubados a 37.0±1.0°C por una hora, y prefijados por 96 horas con la solución fijadora preparada en proporción 4:1 (metanol a 4°C: ácido acético). Luego se reemplazó el prefijador por la solución fijadora 4:1, y se centrifugó las muestras de tejido hasta que tuvieran apariencia blanquecina, por la eliminación de restos de proteínas del tejido hematopoyético. Finalmente, las muestras fueron goteadas sobre portaobjetos previamente enfriados con etanol absoluto a 4°C y a una altura de 1.70m, después deslizando gentilmente por tres ocasiones por el lado que no tiene el tejido; la flama del mechero de alcohol.
Las preparaciones cromosómicas, fueron teñidas con Giemsa al 10%, en buffer de fosfatos a pH 7.0 (Kligerman & Bloom, 1977) por 30min (Arias-Rodriguez et al., 2008; 2009; 2011). Se analizaron las mejores dispersiones cromosómicas mitóticas y meióticas (metafases con los cromosomas no traslapados o encimados) con los objetivos 10X y 40X; y fueron fotodigitalizadas a 100X+1.25X del optovar del microscopio AxioScope-A1, con la AxioCam ERc5s y el programa ZEN/2011 (Carl Zeizz® microscopy GmbH, 2011).
El número cromosómico modal diploide en mitosis y meiosis, se estimó con base en el análisis de frecuencias del número de cromosomas mostrado en las metafases digitalizadas, fotos impresas en alta resolución y por conteo directo con el microscopio óptico.
Para armar los cariotipos, cinco metafases en mitosis y meiosis por sexo/especie, con sus respectivos cromosomas individuales, fueron analizados y recortados con el programa Photoshop CS 8.0.1 (Adobe®). Mientras que los cromosomas, fueron insertados individualmente, en base a su longitud para ensamblar el cariotipo con las herramientas de dibujo del programa Microsoft Word 2011©.
Se calcularon las longitudes relativas (L.R) (Al-Aish, 1969), L.R=longitud promedio del par cromosómico en micrómetros (μm)/ la longitud total del complemento cromosómico (100). El cariotipo se clasificó con base en los parámetros citogenéticos de Levan et al. (1964), utilizando la proporción de brazos r=q/p (q= brazo largo/p= brazo corto), índice centromérico i=100(p/p+q), la diferencia entre brazos cromosómicos d=r-1(10)/r+1. El cariotipo típico se armó en orden descendente de longitud. Además, se utilizó la clasificación por grupos recomendada por Bickham (1975): donde el primer grupo “A” se integró de los macrocromosomas que tienen el centrómero en posición media o submedia “msm” de acuerdo con Levan et al. (1964), mientras que el grupo “B” consideró aquellos macrocromosomas con centrómero en posición terminal o subterminal “stt” basado en Levan et al. (1964), y el grupo “C” se identificó por mostrar los elementos más pequeños (o microcromosomas) y de morfología monorrámea por ello la ubicación del centrómero fue en algunos casos difícil de establecer.
Resultados
En el pochitoque tropical K. leucostomum, se lograron contabilizar 230 dispersiones cromosómicas, de las cuales 184 metafases fueron de origen mitótico o pancreático y 46 correspondieron a campos cromosómicos bivalentes en meiosis I que se derivaron del tejido gonádico. El análisis del tejido mitótico permitió observar 162 metafases, con conteos cromosómicos de 2n=56 cromosomas, lo que fue congruente con el 88.0% del total de campos analizados, y por su mayor frecuencia se le asignó la moda diploide (M2n) de cromosomas de la especie. También, se observaron conteos mitóticos variables de 47 (8.1%) y 42 (3.8%) cromosomas. Mientras en meiosis I, 38 dispersiones metafásicas haploides con cromosomas bivalentes, correspondieron a la moda haploide (M1n) de 1n=28 cromosomas, lo que correspondió con el 82.6% del total de dispersiones cromosómicas analizadas. Metafases de origen meiótico, con conteos variables fueron también observadas y estas fluctuaron entre 42 (2.1%), 47 (2.1%) y 56 (13.0%) cromosomas respectivamente. Adicionalmente, fue posible observar presencia de microcromosomas “B” en las metafases de origen mitótico y meiótico.
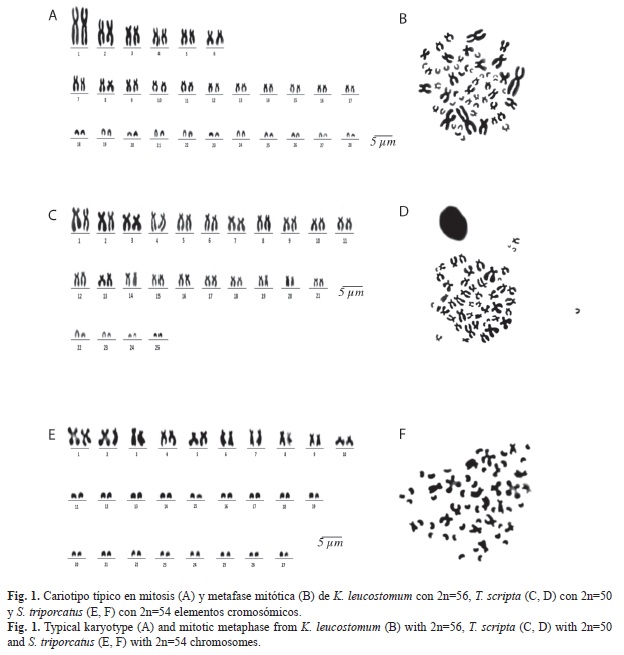
Los cromosomas mitóticos del cariotipo de K. leucostomum, mostraron en promedio longitudes (p+q) desde 9.58±3.13μm en el primer par cromosómico birrámeo, 4.26±0.61μm en el par cromosómico birrámeo noveno y 3.59±0.22μm en el par cromosómico birrámeo diecisiete (Cuadro 1, Fig. 1A, Fig. 1B). Mientras el primer par cromosómico dieciocho de tipo monorrámeo mostró en promedio 4.02±0.20μm de su único brazo cromosómico pequeño (q), mientras que el último par monorrámeo veintiocho logró definirse en promedio con 2.52±0.33μm (Cuadro 1).
Las longitudes del complemento cromosómico bivalente haploide mostraron las siguientes longitudes promedio, para el primer par cromosómico de tipo birrámeo 6.95±1.08μm, en el noveno cromosoma con 4.77±0.53μm, y en el cromosoma diecisiete 3.02±0.55μm. En el cromosoma dieciocho de tipo monorrámeo, el promedio fue de 3.47±0.55μm en el brazo pequeño (q) y en el último cromosoma monorrámeo número veintiocho, el promedio fue de 1.71±0.27μm (Cuadro 1).
La longitud promedio total del complemento cromosómico diploide en mitosis (CCDMI) mostró 239.53±16.5μm y del complemento cromosómico haploide bivalente en meiosis (CCHME) fue de 111.99±17.8μm (Cuadro 1).
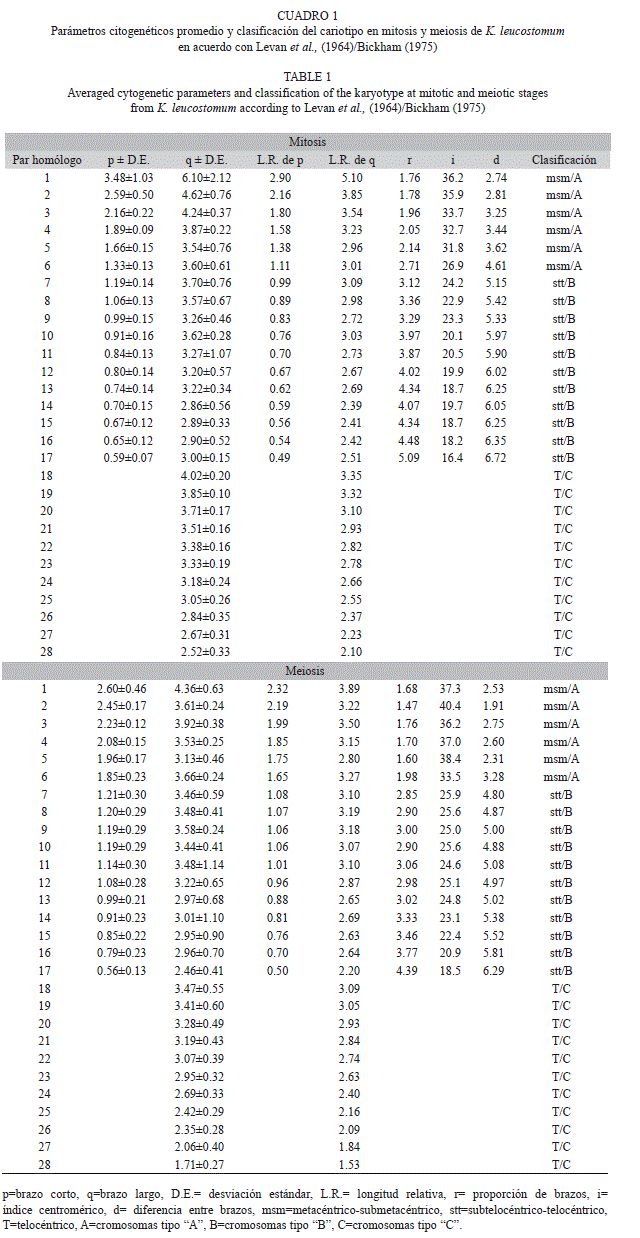
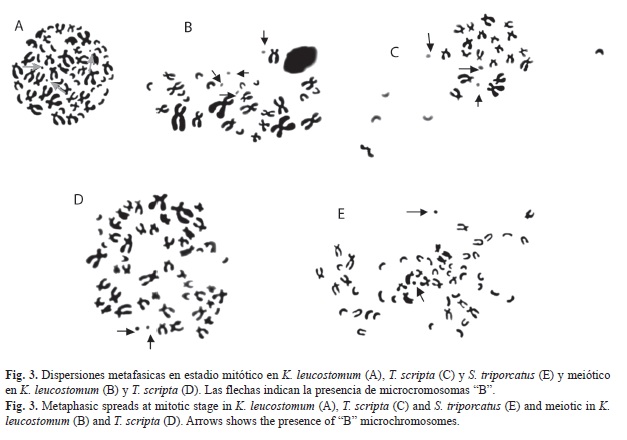
El cariotipo típico de K. leucostomum en mitosis, se caracterizó por mostrar la siguiente fórmula cromosómica basado en Levan et al. (1964)/Bickham (1975): 12 cromosomas metacéntrico-submetacéntrico “msm”/“A”+22 cromosomas subtelocéntrico-telocéntrico “stt”/“B”+22 cromosomas telocéntrico “T”/“C”, con número fundamental (NF) de 90 (Fig. 1A, Fig. 1B). En meiosis se identificó fórmula cromosómica que correspondió con la mitad en mitosis (Fig. 2A, Fig, 2B). No fue posible identificar diferencias entre las metafases de los machos y hembras. Adicionalmente se encontraron dispersiones cromosómicas con presencia de microcromosomas “B”. En condición mitótica se identificó la presencia desde cero, uno, dos y hasta tres microcromosomas en algunas metafases; tales microcromosomas mostraron en promedio 0.47±0.06μm. Mientras, en meiosis se logró observar también nula presencia en algunas metafases y en otras desde uno, dos, tres y hasta cuatro microcromosomas que en promedio midieron 0.70±0.14μm (Fig. 3A, Fig. 3B).
En la tortuga T. scripta, se contabilizaron 314 dispersiones cromosómicas, de ellas 141 metafases de campos cromosómicos en mitosis del páncreas y 173 metafases correspondieron a campos cromosómicos bivalentes en meiosis I de las gónadas. El análisis del tejido mitótico de T. scripta, permitió observar 141 metafases con conteos cromosómicos de 2n=50 cromosomas, lo que fue congruente con el 70% del total de campos analizados, por su alta frecuencia se le asignó la moda diploide (M2n) de 2n=50 cromosomas. También, se logró observar conteos mitóticos variables de 40 (15%) y 58 (15%) cromosomas, respectivamente.
En los conteos en meiosis I, fueron identificadas 97 dispersiones metafásicas haploides con cromosomas bivalentes, correspondieron a la moda haploide (M1n) 1n=25 cromosomas, con el 74% del total de dispersiones cromosómicas analizadas. Mientras que en el resto de metafases de origen meiótico, se pudo observar conteos variables que oscilaron entre 20 (6%), 30 (17%) y 50 cromosomas respectivamente (3%).
Las longitudes de los cromosomas mitóticos, presentan medidas de (p+q) 6.58±0.65μm en el primer par cromosómico de tipo birrámeo, así como 2.62±0.78μm en el último par cromosómico de tipo birrámeo. Los cromosomas monorrámeos en mitosis presentan medidas de 3.12±0.45μm a 2.37±0.29μm (Cuadro 2, Fig. 1C, Fig. 1D).
Las medidas en micrómetros (p+q) de los cromosomas meióticos de la tortuga T. scripta corresponden a 7.92±2.32μm en su primer cromosoma de tipo birrámeo, así como 2.78±0.72μm en el último cromosoma de tipo birrámeo. Mientras que los cromosomas de tipo monorrámeo presentaron medidas de 2.52±0.30μm a 1.96±0.29μm (Cuadro 2, Fig. 1).
La longitud promedio total del complemento cromosómico diploide en mitosis, mostró 188.88±12.2μm y del complemento cromosómico haploide bivalente en meiosis fue de 108.99±11.8μm.
El cariotipo típico en mitosis presentó la siguiente clasificación cromosómica basado en Levan et al., (1964)/Bickham (1975) fue de 32 cromosomas metacéntrico-submetacéntrico “msm”/“A”+10 cromosomas subtelocéntrico- -telocéntrico “stt”/“B”+8 cromosomas telocéntricos “T”/“C” (Cuadro 2) con número fundamental (NF) de 92 (Fig. 1C, Fig. 1D). En meiosis se identificó fórmula cromosómica que correspondió con la mitad de la mostrada en mitosis (Fig. 2C, Fig. 2D). Adicional al complemento cromosómico del cariotipo típico en T. scripta, se identificaron dispersiones cromosómicas con presencia de microcromosomas tipo “B”. En fase mitótica se identificaron de cero, uno y hasta dos microcromosomas con medidas promedio de 0.80±0.28μm. En fase meiótica se identificó la presencia de cero, uno, dos y hasta tres microcromosomas con medidas promedio de 0.87±0.23μm (Fig. 3C, Fig. 3D).
En la tortuga S. triporcatus, se contabilizaron 104 dispersiones cromosómicas de origen mitótico de tejido pancreático, de las cuales 81 metafases correspondieron a campos cromosómicos de 2n=54 cromosomas, lo que corresponde al 78% modal diploide (M2n) de las dispersiones analizadas. También, se identificó la presencia de dispersiones cromosómicas con 58 (10%) cromosomas y 111 (5%) cromosomas. En el caso de material celular procedente del tejido gonádico, este no cumplió con la calidad requerida para este tipo de estudios, por ello no fueron utilizados ni reportados.
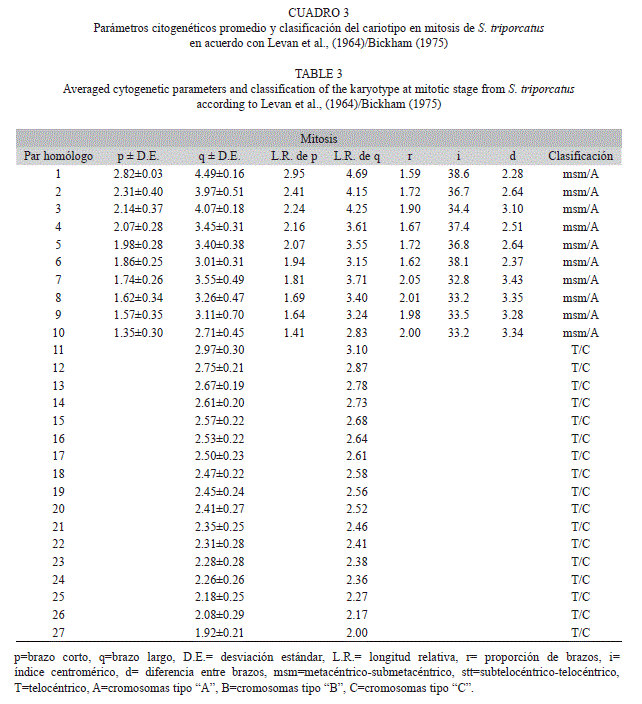
Las medidas en micrómetros de los cromosomas mitóticos fueron de 7.31±0.19μm en el primer par cromosómico de tipo birrámeo, mientras que en el último par de cromosomas de este tipo; las medidas fueron de 3.96±0.75μm. Por otro lado, los cromosomas monorrámeos presentaron medidas de 2.97±0.30μm a 1.92±0.21μm (Cuadro 3, Fig. 1E, Fig. 1F).
La longitud promedio total del complemento cromosómico diploide en mitosis mostró 95.7±.10.9μm.
El cariotipo típico, presentó la siguiente clasificación cromosómica basado en Levan et al. (1964)/Bickham (1975) fue de 20 cromosomas metacéntrico submetacéntrico “msm”/“A”+34 cromosomas telocéntricos “T”/“C” (Cuadro 3), con número fundamental (NF) de 74 (Fig. 1E, Fig. 1F). En S. triporcatus, también se identificó la presencia de microcromosomas “B”, siendo únicamente observables en condición mitótica, donde se descubrió la presencia de cero, uno, dos y hasta tres microcromosomas con medida promedio de 0.65±0.07μm (Fig. 3E).
Discusión
El número modal diploide de 2n=56 cromosomas confirmado por el número modal haploide de 1n=28 cromosomas bivalentes obtenidos, fue similar al reportado tempranamente en especímenes de K. leucostomum de origen desconocido por Gorman (1973), Moon (1974) y Killebrew (1975).
Los estudios citogenéticos reportados en los kinosternidos, muestran números cromosómicos variables de 2n=56 en K. leucostomum por Gorman (1973), Moon (1974) y Killebrew (1975); de 2n=54 en S. triporcatus por Bull et al. (1974) y Sites et al. (1979a) Sites, Bickham & Haiduk (1979b); y 2n=50 cromosomas solo en Sternotherus odoratus (Risley, 1936). Lo que hace pensar en presencia de números cromosómicos especie específicos.
A pesar de las diferencias existentes en los números cromosómicos en los kinosternidos, como bien lo mencionan los antecedentes cariológicos de este grupo en particular, es posible mencionar que el 2n=56 y 2n=54 cromosomas es el número diploide típico en la familia Kinosternidae (Bickham & Carr, 1983; Mayer-Goyenechea, 2001).
Basados en la información de los trabajos citogenéticos en el orden Testudines, es difícil hacer una comparación precisa, de las fórmulas cromosómicas de las especies que integran el orden. En acuerdo con Ortiz et al. (2005) para la clasificación de los cromosomas y el establecimiento del cariotipo se ha utilizado ampliamente la nomenclatura propuesta por Bickham (1975). Debido a que el criterio que se emplea comúnmente, solamente se basa en la morfología de los cromosomas y toma solamente en cuenta a los birrámeos, ya que estos son los elementos con mayor tamaño en el cariotipo. Por lo anterior, los elementos menores o microcromosomas quedan descartados del criterio de clasificación, lo que incrementa la pérdida de información derivada de los cariotipos. Adicionalmente, los estudios antiguos y los más recientes en el orden Testudines; se han venido reportando sin adicionar las medidas de los elementos que integran el cariotipo, por ello no pueden ser comparados. Sin embargo, aquellos estudios en los que se reportan las longitudes de los brazos cortos (p) y largos (q) como es el caso de este trabajo, permite emplear la nomenclatura recomendada por Levan et al. (1964) y la de Bickham (1975) simultáneamente, lo que genera una mayor comprensión de la estructura cariotípica de las especies estudiadas.
Se comprobó que la tortuga S. triporcatus de Tabasco, presenta número diploide de 2n=54 cromosomas, al igual que los estudios previos en la subfamilia Staurotypinae (Bull et al., 1974; Moon, 1974; Killebrew, 1975; Sites et al., 1979a); en los estudios previos en dichas especies, se describe aisladamente que la especie S. triporcatus y C. angustatus presentan el número diploide de 2n=56 cromosomas (Gorman, 1973). La variación en los números cromosómicos, también ha sido observada en K. subrubrum con 2n=54 y S. odoratus con 2n=50 que pertenecen a la subfamilia Kinosterninae (Risley, 1936; Forbes, 1966).
Estructuralmente, los estudios citogenéticos en la subfamilia Staurotypinae muestran que la especie S. salvinii presenta la composición cromosómica de siete pares de cromosomas tipo “A”, cinco pares de cromosomas “B”, y 15 pares de cromosomas “C”, con fórmula Bickham 7A:5B:15C (Sites et al., 1979a), presentando diferencias en el número de microcromosomas con respecto a lo reportado en S. triporcatus de Tabasco. La variación en la estructura del complejo cromosómico, es una manifestación de la diversidad genética que existe en relación con la distribución geográfica como ocurre en la familia Kinosternidae (Frair, 1972). Lo que hace suponer de la importancia biológica que tienen los microcromosomas como mecanismo genético para ampliar la diversidad genética presente en varias especies (Olmo, 2005; Hernández-Guzmán et al., 2011).
Los cromosomas heteromórficos o cromosomas sexuales encontrados en S. triporcatus por Bull et al. (1974) y en S. salvinii por Sites et al. (1979a, b), no fue demostrado con evidencia tangible en los organismos de Tabasco evaluados para el presente estudio. Lo anterior, hace necesaria la aplicación de herramientas de citogenética molecular, que permitan comparar el genoma total (CGT) entre sexos: hembra/ macho o viceversa en todas las especies de tortugas con el fin de identificar evidencias tangibles entre pares cromosómicos.
La reducción o incremento en el número de cromosomas, se puede deber también, probablemente a la fusión centromérica de cromosomas monorrámeos y fisión centromérica de cromosomas birrámeos (Baker & Bickham, 1986). Dicho planteamiento toma credibilidad en las tortugas, porque sus cariotipos en la mayoría de los casos muestran cromosomas birrámeos y monorrámeos lo que hace factible que ocurra cualquiera de los fenómenos señalados con anterioridad (Bull et al., 1974; Moon, 1974; Killebrew, 1975; Sites et al., 1979a).
El número fundamental en las tortugas de Tabasco K. leucostomum con NF=90 y S. triporcatus NF=74 demuestra otra de las diferencias citogenéticas en la familia Kinosternidae. Dicha diferencia en el número fundamental se debe a que en la especie S. triporcatus existe mayor número de microcromosomas monorrámeos (17 pares de microcromosomas), mientras que en K. leucostomum sólo existen 11 pares de microcromosomas en su cariotipo, lo que origina que el número fundamental sea más elevado en K. leucostomum que en S. triporcatus.
El análisis citogenético de los cromosomas de T. scripta de Tabasco, ha presentado diferencias en la morfología y clasificación de los cromosomas en el complemento cariotípico con relación a otros estudios en Trachemys y otros taxones de la familia (Cleiton & Giuliano-Caetano, 2008; Martínez et al., 2009; Kasai et al., 2012)
El estudio más reciente en dicha especie en Brasil, indica que las tortugas de esta región en particular presentan el número diploide de 2n=50 cromosomas y clasificación cromosómica de los dos primeros pares de cromosomas como metacéntricos, seis pares de cromosomas submetacéntricos, cinco pares de cromosomas acrocéntricos y 12 pares de microcromosomas y el número fundamental calculado fue de NF=62 (Cleiton & Giuliano Caetano, 2008). Las diferencias mostradas con los individuos de T. scripta de Tabasco es notoria ya que se logró identificar 42 cromosomas submetacéntricos “sm”+8 cromosomas telocéntricos “T”, con número fundamental calculado de NF=92. El número fundamental es un parámetro citogenético clave para la comparación de las fórmulas cariotípicas, ya que el número fundamental de T. scripta de Tabasco, México presentó mayor número de macrocromosomas birrámeos y menor número de microcromosomas, que el cariotipo reportado para los especímenes de Brasil, lo que es evidencia de la amplia variabilidad genética en la especie.
Otro estudio citogenético en el género Trachemys es presentado por los mismos autores Cleiton & Giuliano-Caetano (2008) en la especie T. dorbigni, donde se identificó el número diploide de 2n=50 cromosomas con clasificación de dos pares de cromosomas metacéntricos y seis pares de cromosomas submetacéntricos, cinco pares de cromosomas acrocéntricos y 12 pares de microcromosomas, fórmula cromosómica idéntica a la de T. scripta de la misma región de Brasil, por lo que las diferencias en la estructura del cariotipo y número fundamental en T. scripta de Tabasco, México son las mismas con respecto a T. scripta y T. dorbigni del Brasil. Tales resultados generan la idea de posible presencia de errores taxonómicos en la identificación de las especies o bien de ocurrencia de especies crípticas que aún mantienen cariotipos compartidos.
Los estudios citogenéticos en la familia Emydidae, demuestran al igual que en la familia Kinosternidae, presencia de variabilidad en el número de cromosomas entre las especies de este grupo en particular como en Clemmys insculpta con 2n=48 cromosomas por Forbes (1966), Cuora amboinensis, Emys orbicularis, Chrysemys picta, Graptemys geographica, Emydoidea blandingi Malayemys subtrijuga, Orlitia borneensis, Siebenrockiella crassicollis, T. scripta, T. dorbigni con 2n=50 cromosomas por Van Brink (1959), Gorman (1973), Ivanov (1973), Carr & Bickham (1986), Cleiton & Giuliano-Caetano (2008), Martínez et al. (2009) Callagur borneoensis, Chinemys kwangtungensis, Cuora ambioenensis, Cyclemis dentata, Heosemys grandis y Mauremys japonica con 2n=52 cromosomas, siendo el número 2n=50 el de mayor frecuencia. Sin embargo, el grupo de los emídidos en comparación con los kinosternidos presentan números diploides muy discrepantes entre sus miembros; un claro ejemplo es la especie Terrapene carolina, en la cual Jordan (1914) reportó 2n=32 cromosomas.
De acuerdo a la revisión de los estudios citogenéticos en las tortugas de la familia Emydidae, se pueden identificar cuatro grupos con niveles de ploidía diferentes: el primero con 2n=50 cromosomas como número diploide en las especies Trachemys, Emys, Chrysemys, Graptemys, Terrapene, Deirochelys, Malaclemys, Emydoidea. El segundo con 2n=52 cromosomas en las especies Chrysemys scripta elegans, C. s. callirostris, Graptemys barbouri y G. pulchra. El tercero con 2n=48 cromosomas en las especies Clemmys insculpta y C. guttata. Mientras que el cuarto está conformado por una única especie Terrapene carolina, en la cual se reportó en 1914 el número diploide de 32 cromosomas.
La familia Emydidae presenta dos subfamilias caracterizadas por dos tipos de cariotipos. La primera subfamilia, Emydinae o tortugas del nuevo mundo con 2n=50 cromosomas y los Batagurinae o tortugas antiguas con 2n=52 cromosomas (Bickham & Carr, 1983). Otros autores afirman que el número de cromosomas 2n=52 es un representante del cariotipo primitivo en las tortugas relacionadas con la subfamilia Batagurinae, es decir, con tortugas de origen genético antiguo como Sacalia bealei y Siebenrockiella crassicollis (Bickham & Baker, 1976b). Esta información es respaldada por otros estudios en donde las bandas cariotípicas identificadas en Chynemys reevesi y otras especies de este grupo son similares con los miembros de tortugas de la subfamilia Batagurinae (Dowler & Bickham, 1982; Carr & Bickham, 1986). Además, se ha identificado que las Regiones Organizadoras Nucleolares (RON´s) en el grupo primitivo de los batagurinos se localizan en el par nueve de los macrocromosomas del grupo “A” o del complemento cromosómico autosómico, mientras que en los emídidos se localizan solo en un microcromosoma (Carr & Bickham, 1986), condición citológica que resalta la importancia evolutiva de los microcromosomas como un probable elemento para separar grupos taxonómicos en las tortugas.
Los cromosomas adicionales al complemento cariotípico son considerados como cromosomas supernumerarios o cromosomas tipo “B” (Jones & Rees, 1982; King & Stansfield, 2002). La caracterización de presencia y ausencia de microcromosomas “B”, en el complemento cariotípico en mitosis y meiosis de los organismos pueden ser también parte de un citotipo particular en las especies, ocasionados por las actividades antropogénicas (Arias-Rodriguez et al., 2007; 2008; Camacho, Sharbel & Beukeboom, 2000). No obstante, también existe la posibilidad de que la presencia de microcromosomas sea causada por hibridación, siendo estas las dos vías o condiciones para la presencia de microcromosomas “B” en las especies (Mestriner et al., 2000; Perfectti & Werren 2001; Cabrero et al., 2003). En diversas especies de tortugas y en la mayoría de los reportes citogenéticos, se tiene el registro de presencia de microcromosomas como complemento de los cromosomas “A” del cariotipo típico (Bickham & Baker, 1976b; 1979; Porter, Haiduk & De Queiroz 1994; Badenhorst, Stanyon, Engstrom & Valenzuela, 2013).
Sin embargo, en la literatura actual no se ha reportado presencia de microcromosomas tipo “B” en las tortugas. En México se han identificado microcromosomas “B” en diversas especies de organismos acuáticos como los caracoles de tinte Plicopurpura pansa y Plicopurpura columellaris (Arias-Rodriguez, González-Hermoso, Fletes-Regalado, Rodríguez-Ibarra & Del Valle-Pignataro, 2007), en los peces tropicales Petenia splendida y Atractosteus tropicus (Arias-Rodriguez et al., 2008; 2009) y en la rana arborícola Smilisca baudinii (Hernández-Guzmán et al., 2011), por lo que la presencia de microcromosomas “B” en las tortugas K. leucostomum, T. scripta y S. triporcatus de Tabasco incrementa el número de incidencia de especies con presencia de cromosomas supernumerarios, encontrando una relación constante en las especies con este citotipo cariológico específico, en donde todos los organismos con microcromosomas “B” son acuáticos o semiacuáticos. La hipótesis del origen de los microcromosomas “B” presentes en la biodiversidad mexicana, puede tener relación con la presencia de componentes xenobióticos en niveles o concentraciones elevadas (Arias-Rodriguez et al., 2007; 2008; 2009; Hernández-Guzmán et al., 2011); esta idea es reforzada por los antecedentes históricos de presencia de metales pesados en sistemas acuáticos y terrestres del Golfo de México y principalmente del estado de Tabasco, de donde son procedentes las tres especies de tortugas K. leucostomum, T. scripta y S. triporcatus (Rosas et al., 1983; Villanueva & Botello, 1992; Fiedler et al., 2009; De la Cruz-Pons et al., 2012). También algunos estudios han demostrado la relevancia de los microcromosomas, por ejemplo en la tortuga Pelodiscus sinensis en la que el 50% de los genes funcionales se encuentran alojados en los microcromosomas (Kuraku et al., 2006), los cuales son ricos en secuencias de Guanina-Citosina más que en los macrocromosomas, a pesar de que los microcromosomas sólo representan el 23% del total de ADN (Olmo, Capriglione & Odierna, 2002; Olmo, 2008). La suma total de las longitudes del complemento cromosómico en K. leucostomum en mitosis (239.53±16.5μm) y en meiosis (111.99±17.8μm) está correlacionado con el contenido total de ADN diploide de 2.8 pico gramos (pg) y de 2C=2.6 pg reportados para la tortuga K. subrubrum por Olmo (1976) y De Smet (1981), respectivamente. Mientras que en S. triporcatus el CCDMI sumó en total 95.7±.10.9μm que demostró un tamaño del genoma intermedio en comparación con el reportado en los miembros de la familia Kinosternidae. Mientras que en T. scripta el tamaño del CCDMI (188.88±12.2μm) y el CCHME (108.99±11.8μm) también resultó correlacionado al complemento diploide 2C=1.9pg reportado en la misma especie por De Smet (1981). Sin embargo, existe grandes diferencias con el CCDMI de T. scripta con 2C=2.6pg por Tiersch, Chandler, Wachtel & Elias (1989) y 2C=2.5pg por Lockwood, Seavey, Dillinger & Bickham (1991). Las similitudes y las diferencias reportados en el tamaño del genoma en la familia Kinosternidae y Emydidae, retoma importancia probablemente por la presencia y ausencia de cierto número de microcromosomas tipo “B” en el complemento cromosómico de los cariotipos típicos, ya que los microcromosomas “B” pueden disminuir o aumentar el tamaño del genoma a través de la suma total de las longitudes del complemento cromosómico mediante fusiones de uno o varios microcromosomas “B” que no cuentan con un par homologo. La fusión de microcromosomas vs microcromosomas, microcromosomas vs macrocromosomas se ha observado como un fenómeno esencial durante la evolución cariotípica de varias especies de aves (Tegelström & Ryttman, 1981; Vos et al., 2011), reptiles (Olmo, 2008; Organ, Moreno & Edwards, 2008; Srikulnath et al., 2009) y anfibios (Vos et al., 2011).
En conclusión, las tortugas son un grupo de vertebrados que citogenéticamente se vuelve complicado correlacionar con los parámetros cromosómicos tradicionales, esto debido a la variabilidad cromosómica que existe en número cromosómico y número fundamental (NF) en una especie, además por similitudes entre las diferentes familias de Testudines. Por otro lado, este taxón se vuelve un ejemplo interesante para el estudio de fenómenos de especiación a través de la evolución cromosómica.
También, se descarta la presencia de heteromorfismo cromosómico por falta de evidencia tangible entre los cromosomas del cariotipo típico de las especies estudiadas.
La presencia de microcromosomas tipo “B” en las tortugas de Tabasco demuestra el incremento de incidencia de este citotipo particular en la biodiversidad mexicana, relacionándose en todos sus casos con ambientes acuáticos contaminados. Además, es el primer registro de la presencia de microcromosomas “B” en este grupo de reptiles para la región sureste de México.
El presente estudio demuestra la presencia de variación en la fórmula cromosómica de las especies con respecto a los estudios citogenéticos reportados, por ello, se recomienda ampliar los estudios con un procedimiento citológico estandarizado, que permita hacer comparaciones detalladas de la estructura cariotípica de especies que habitan áreas geográficas diversas, considerando sobre todo el significado y la relevancia evolutiva que pudieran tener la presencia y ausencia de microcromosomas “B” como pauta para la formación o separación de nuevas especies (Hernández-Guzmán et al., 2011; Ferro et al., 2012).
Agradecimientos
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca para estudios de posgrado del primer autor (JHG) y el apoyo parcial mediante los proyectos 167686/204264 a LAR. El permiso federal de recolecta en México, fue el otorgado por la SEMARNAT con número de oficio: SGPA/DGVS/04315/11. El estudio recibió apoyo en especies del rancho Las Amazonas.
Referencias
Al-Aish, M. (1969). Human chromosome morphology I. Studies on normal chromosome characterization, classification and karyotyping. Canadian Journal of Genetics and Cytology, 11, 370-381. [ Links ]
Arai, R. (2011). Fish karyotypes a check list. Springer. [ Links ]
Arias-Rodriguez, L., González-Hermoso, J. P., Fletes- Regalado, H., Rodríguez-Ibarra, L. E., & Del Valle- Pignataro, G. (2007). Cariotipos de los caracoles de tinte Plicopurpura pansa y Plicopurpura columellaris (Gastropoda: Muricidae). Revista de Biología Tropical, 55(3-4), 853-866. [ Links ]
Arias-Rodriguez, L., Ibarra-Castro, L., & Páramo-Delgadillo, S. (2008). Los cromosomas mitóticos y meióticos del pez tropical Petenia splendida (Cichlidae). Revista de Biología Tropical, 56(2), 895-907. [ Links ]
Arias-Rodriguez, L., Páramo-Delgadillo, S., Contreras- Sánchez, W. M., & Álvarez González, C. A. (2009). Cariotipo del pejelagarto tropical Atractosteus tropicus (Lepisosteiformes: Lepisosteidae) y variación cromosómica en sus larvas y adultos. Revista de Biología Tropical, 57(3), 529-539. [ Links ]
Arias-Rodriguez, L., Indy, J. R., Ahumada-Hernández, R. I., Barragán-Cupido, H., Ávalos Lázaro, A. A., & Páramo-Delgadillo, S. (2011). Caracterización cariotípica en mitosis y meiosis del robalo blanco Centropomus undecimalis (Pisces: Centropomidae). Revista de Biología Tropical, 59(2), 683-692. [ Links ]
Badenhorst, D., Stanyon, R., Engstrom, T., & Valenzuela, N. (2013). A ZZ/ZW microchromosome system in the spiny softshell turle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae. Chromosome Research, 21(2), 137-147. [ Links ]
Baker, R. J. & Bickham, J. W. (1986). Speciation by monobrachial centric fusions. Proceedings of the National Academy of Sciences, Unites States of America, 83, 8245-8248. [ Links ]
Bickham, J. W. (1975). A cytosystematic study of turtles in the genera Clemmys, Mauremys and Sacalia. Herpetologica, 31(2), 198-204. [ Links ]
Bickham, J. W. & Baker, R. J. (1976a). Karyotypes of some neotropical turtles. Copeia, 1976(4), 704-707. [ Links ]
Bickham, J. W. & Baker, R. J. (1976b). Chromosome homology and evolution of emydid turtles. Chromosoma, 54(3), 201-219. [ Links ]
Bickham, J. W. & Baker, R. J. (1979). Canalization model of chromosomal evolution. Bulletin of the American Museum of Natural History, 13(1), 70-84. [ Links ]
Bickham, J. W. & Carr, J. L. (1983). Taxonomy and phylogeny of the higher categories of cryptodiran turtles based on a cladistic analysis of chromosomal data. Copeia, 4, 918-932. [ Links ]
Bull, J. J., Moon, R. G., & Legler, J. M. (1974). Male heterogamety in kinosternid turtles (genus Staurotypus). Cytogenetics and Cell Genetics, 13(5), 419-425. [ Links ]
Cabrero, J., Bakkali, M., Bugrov, A., Warchalowska-Sliwa, E., López-León, M. D., Perfectti, F., & Camacho, J. P. M. (2003). Multiregional origin B chromosomes in the grasshopper Eyprepocnemis plorans. Chromosoma, 112, 207-211. [ Links ]
Camacho, J. P. M., Sharbel, T. F., & Beukeboom, L. W. (2000). B-Chromosome evolution. Philosophical Transactions of the Royal Society B: Biological Sciences, 355(1394), 163-178. [ Links ]
Camacho-Mosquera, L. & Amorocho, D. (2008). Caracterización genética de la colonia reproductiva de la tortuga marina golfina Lepidochelys olivácea en el parque nacional natural Gorgona (Pacífico Colombiano) a partir de secuencias de ADN mitocondrial. Boletín de Investigaciones Marinas y Costeras, 37(1), 77-92. [ Links ]
Carr, J. L., Bickham, J. W., & Dean, R. H. (1981). The karyotype and chromosomal banding patterns of the Central American river turtle Dermatemys mawii. Herpetologica, 37(2), 92-95. [ Links ]
Carr, J. L. & Bickham, J. W. (1986). Philogenetic implications of karyotypic variation in the Batagurinae (Testudines: Emydidae). Genetica, 70, 89-106. [ Links ]
Cleiton, F. & Giuliano Caetano, L. (2008). Cytogenetic characterization of two turtle species: Trachemys dorbigni and Trachemys scripta elegans. Caryologia, 61(3), 253-257. [ Links ]
De la Cruz-Pons, A., Zavala-Cruz, J., Guerrero-Peña, A., Salgado-García, S., Lagunes-Espinoza, L. C., & Gavi-Reyes, F. (2012). Metales pesados en suelos cultivados con caña de azúcar en la Chontalpa Tabasco. Universidad y Ciencia, 28(2), 119-130. [ Links ]
De Smet, W. H. O. (1981). The nuclear feulgen-DNA content of the vertebrates (especially reptiles), as measured by fluorescence cytophotometry, with notes on the cell and chromosome size. Acta Zoologica et Pathologica Antverpiensia, 76, 119-167. [ Links ]
Denton, T. E. (1973). Fish chromosome methodology. Chicago, Illinois, Estados Unidos de América: Charles C. Tomas. [ Links ]
Dowler, R. C. & Bickham, J. W. (1982). Chromosomal relationships of the tortoises (family Testudinidae). Genetica, 58, 189-197. [ Links ]
Ferro, J. M., Marti, D. A., Bidau, C. J., Suárez, P., Nagamachi, C. Y., Pieczarka, J. C., & Baldo, D. (2012). B chromosome in the tree frog Hybsiboas albopunctatus (Anura: Hylidae). Herpetologica, 68(4), 482-490. [ Links ]
Fiedler, S., Siebe, C., Herre, A., Roth, B., Cram, S., & Stahr, K. (2009). Contribution of oil industry activities to environmental loads of heavy metals in the Tabasco lowlands, Mexico. Water, Air and Soil Pollution, 197, 35-47. [ Links ]
Forbes, W. R. Jr. (1966). A cytological study of the Chelonia. (Tesis de Doctorado). University of Connecticut. [ Links ]
Frair, W. (1972). Taxonomic relations among chelydrid and kinosternid turtles elucidated by serological tests. Copeia, 1, 97-108. [ Links ]
Gorman, G. C. (1973). The chromosomes of the reptilian, a cytotaxonomic interpretation. In A. B. Chiarelli & E. Capanna (Eds.), Cytotaxonomy and vertebrate evolution (pp. 347 424). New York, USA.: Editorial Academic Press. [ Links ]
Hernández-Guzmán, J., Arias-Rodriguez, L., & Indy, J. R. (2011). Los cromosomas meióticos de la rana arborícola Smilisca baudinii (Anura: Hylidae). Revista de Biología Tropical, 59(1), 355-362. [ Links ]
Hillis, D. M. (1991). The phylogeny of amphibians: Current knowledge and the role of cytogenetics. In D. M. Green & S. K. Sessions (Eds.), Amphibian cytogenetics and evolution (pp. 7-31). San Diego, California, USA: Academic Press Inc. [ Links ]
IUCN. 2012. IUCN red list of threatened species. Retrieved from www.iucnredlist.org [ Links ]
Ivanov, V. G. (1973). Description of the karyotype of Emys orbicularis L. (Testudines, Emydidade). Tsitologiya, 15, 1059-1061. [ Links ]
Jones, R. & Rees, H. (1982). B chromosomes. London, England: Academic. [ Links ]
Jordan, H. E. (1914). Spermatogenesis in Chrysemys marginata and Cistudo carolina. Science, 39, 438. [ Links ]
Kasai, F., O´Brien, P. C., Martin, S., & Ferguson-Smith, M. A. (2012). Extensive homology of chicken macrochromosomes in the karyotypes of Trachemys scripta elegans and Crocodylus niloticus revealed by chromosome painting despite long divergence times. Cytogenetics Genome Research, 136(4), 303-307. [ Links ]
Killebrew, F. C. (1975). Mitotic chromosomes of turtles, III The Kinosternidae. Herpetologica, 31, 398-403. [ Links ]
King, R. C. & Stansfield, W. D. (2002). A dictionary of genetics. USA: Oxford University Press. [ Links ]
Kligerman, A. D. & Bloom, S. E. (1977). Rapid chromosome preparations from solid tissues of fishes. Journal of the Fisheries Research Board of Canada, 34, 266-269. [ Links ]
Kuraku, S., Ishijima, J., Nishida-Umehara, C., Agata, K., Kuratini, S., & Matsuda, Y. (2006). cDNA-based gene mapping and GC3 profiling in the soft-shelled turtle suggest a chromosomal size-dependent GC bias shared by sauropids. Chromosome Research, 14(2), 187-202. [ Links ]
Lee, J. C. (2000). A field guide to the amphibians and reptiles of the Maya World: the lowlands of Mexico, Northern Guatemala, and Belize. USA: Cornell University Press. [ Links ]
Lemos-Espinal, J. A. & Smith, H. M. (2009). Anfibios y reptiles del estado de Chihuahua, México. Universidad Nacional Autónoma de México, University of Colorado at Boulder, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México, D.F. [ Links ]
Levan, A., Fredga, K., & Sandberg, A. A. (1964). Nomenclature for centromeric position on chromosomes. Hereditas, 52, 201-220. [ Links ]
Lockwood, S. F., Seavey, B. T., Dillinger, R. E., & Bickham, J. W. (1991). Variation in DNA content among age classes of broad whitefish (Coregonus nasus) from the Sagavanirktok river delta. Canadian Journal of Zoology, 69, 1335-1338. [ Links ]
López, E. A., Hernández-Fernández, J., & Bernal-Villegas, J. (2008). Condiciones óptimas de cultivo de linfocitos y análisis parcial del cariotipo de la tortuga cabezona, Caretta caretta (Testudines: Cheloniidae) en Santa Marta, Caribe Colombiano. Revista de Biología Tropical, 56(3), 1459-1469. [ Links ]
Martínez, P. A., Boeris, J. M., Sánchez, J., Pastori, M. C., Bolzán, A. D., & Ledesma, M. A. (2009). Karyotypic characterization of Trachemys dorbigni (Testudines: Emydidae) and Chelonoidis (Geochelone) donosobarrosi (Testudines: Testudinidae), two species of Cryptodiran turtles from Argentina. Genetica, 137, 277-283. [ Links ]
Mayer-Goyenechea, I. G. (2001). Sistemática de reptiles y citogenética. Boletín de la Sociedad Herpetológica Mexicana, 9(1), 13-21. [ Links ]
McCoy, C. J. (1982). Reptilia. In S. H. Hurlbert & A. Villalobos-Figueroa (Eds.), Aquatic biota of Mexico, Central America and the West Indies (pp. 515-520). Aquatic Biota-SDSU Foundation. [ Links ]
Mestriner, C. A., Galetti Jr, P. M., Valentini, S. R., Ruiz, I. R. G., Abel, L. D. S., Moreira Filho, O., & Camacho, J. P. M. (2000). Structural and functional evidence that a B chromosome in the characid fish Astyanax scabripinnis is an isochromosome. Heredity, 85, 1-9. [ Links ]
Mittermeier, R. A. & Goettsch, C. (1992). La importancia de la diversidad biológica de México. In J. Sarukhán & R. Dirzo (Eds.), México ante los retos de la biodiversidad. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México D.F. [ Links ]
Moon, R. G. (1974). Heteromorphism in a kinosternid turtle. Mammalian chromosomes newsletter, 15, 10-11. [ Links ]
Olmo, E. (1976). Genome size in some reptiles. Journal of Experimental Zoology, 195, 305-310. [ Links ]
Olmo, E., Capriglione, T., & Odierna G. (2002). Different genomic evolutionary rates in the various reptile lineages. Gene, 295(2), 317-321. [ Links ]
Olmo, E. (2005). Rate of chromosome changes and speciation in reptiles. Genetica, 125, 185-203. [ Links ]
Olmo, E. (2008) Trends in the evolution of reptilian chromosomes. Integrative and Comparative Biology, 48(4), 486-493. [ Links ]
Organ, C. L., Moreno, R. G., & Edwards, S. V. (2008). Three tiers of genome evolution in reptiles. Integrative and Comparative Biology, 48(4), 494-504. [ Links ]
Ortiz, M. L., Rodríguez, P. A., & Bueno, M. L. (2005). Caracterización citogenética de la tortuga sabanera Podocnemis vogli (Reptilia: Testudinata: Podocnemididae). Acta Biológica Colombiana, 10(1), 19-33. [ Links ]
Perfectti, F. & Werren, J. H. (2001). The interspecific origin of B chromosomes: experimental evidence. Evolution, 55(5), 1069-1073. [ Links ]
Porter, C. A., Haiduk, M. W., & De Queiroz, K. (1994). Evolution and phylogenetic significance of ribosomal gene location in chromosomes squamate reptiles. Copeia, 2, 302-313. [ Links ]
Reed, K. M., Hanks, B. G., Bickham, J. W., Rhodin, A. G. J., Greenbaum, I. F., Mittermeier, R. A., & Fedullo, L. P. (1991). Cytogenetic analysis of the pleurodine turtle Phrynops hogei and its taxonomic implications. Amphibia-Reptilia, 12, 203-212. [ Links ]
Risley, P. L. (1936). The chromosomes of the male musk turtle, Sternotherus odoratus L. Cytologia, 7, 232-241. [ Links ]
Rosas, I., Báez, A., & Belmont, R. (1983). Oyster (Crassostrea virginica) as indicator of heavy metal pollution in some lagoons of the Gulf of Mexico. Water, Air and Soil Pollution, 20(1), 127-135. [ Links ]
Schmid, M., Steinlein, C., Bogart, J. P., Feichtinger, W., León, P., La Marca, E., Díaz, L. M., Sanz, A., Chen, S. H., & Hedges, S. B. (2010). The chromosomes of terraranan frogs: Insights into vertebrate cytogenetics. Cytogenetics Genome Research, 130-131(1-8), 1-568. [ Links ]
SEMARNAT. (2010). NORMA Oficial Mexicana NOM- 059-SEMARNAT-2010, Diario Oficial de la Federación, segunda sección, 30 de diciembre del 2010: 1-78. [ Links ]
Sites, J. W. Jr; Bickham, J. W., Haiduk, M. W., & Iverson, J. B. (1979a). Banded karyotypes of six taxa of kinosternid turtles. Copeia, 4, 692-698. [ Links ]
Sites, J. W., Bickham, J. W., & Haiduk, M. W. (1979b). Derived X chromosome in the turtle genus Staurotypus. Science, 206(4425), 1410-1412. [ Links ]
Srikulnath, K., Nishida, C., Matsubara, K., Uno, Y., Thongpan, A., Suputtitada, S., Apisitwanich, S., & Matsuda, Y. (2009). Karyotypic evolution in squamate reptiles: comparative gene mapping revealed highly conserved linkage homology between the butterfly lizard (Leiolepis reevesii rubritaeniata, Agamidae, Lacertilia) and the Japanese four-striped rat snake (Elaphe quadrivirgata , Colubridae, Serpentes). Chromosome Research, 17(8), 975-986. [ Links ]
Tegelström, H. & Ryttman, H. (1981). Chromosomes in birds (Aves): evolutionary implications of macro-and microchromosome numbers and length. Hereditas, 94(1), 225-233. [ Links ]
Tiersch, T. R., Chandler, R. W., Wachtel, S. S. & Elias, S. (1989). Reference standards for flow cytometry and application in comparative studies of nuclear DNA content. Cytometry, 10, 706-710. [ Links ]
Van Brink, J. M. (1959). L´expression morphologique de la digametie chex les sauropsides et les monotremes. Chromosoma, 10(1), 1-72. [ Links ]
Villanueva, S. & Botello, A. V. (1992). Metales pesados en la zona costera del Golfo de México y Caribe mexicano: una revisión. Revista Internacional de Contaminación Ambiental, 8(1), 47-61. [ Links ]
Voss, S. R., Kump, D. K., Putta, S., Pauly, N., Reynolds, A., Henry, R. J., Basa, S., Walker, J. A., & Smith, J. J. (2011). Origin of amphibian and avian chromosomes by fission, fusion, and retention of ancestral chromosomes. Genome Research, 21(8), 1306-1312. [ Links ]
West, R. C., Psuty, N. P., & Thom, B. (1987). Las tierras bajas de Tabasco. Biblioteca Básica Tabasqueña. Villahermosa, Tabasco, México. [ Links ]
Arai, R. (2011). Fish karyotypes a check list. Springer. [ Links ]
Arias-Rodriguez, L., González-Hermoso, J. P., Fletes- Regalado, H., Rodríguez-Ibarra, L. E., & Del Valle- Pignataro, G. (2007). Cariotipos de los caracoles de tinte Plicopurpura pansa y Plicopurpura columellaris (Gastropoda: Muricidae). Revista de Biología Tropical, 55(3-4), 853-866. [ Links ]
Arias-Rodriguez, L., Ibarra-Castro, L., & Páramo-Delgadillo, S. (2008). Los cromosomas mitóticos y meióticos del pez tropical Petenia splendida (Cichlidae). Revista de Biología Tropical, 56(2), 895-907. [ Links ]
Arias-Rodriguez, L., Páramo-Delgadillo, S., Contreras- Sánchez, W. M., & Álvarez González, C. A. (2009). Cariotipo del pejelagarto tropical Atractosteus tropicus (Lepisosteiformes: Lepisosteidae) y variación cromosómica en sus larvas y adultos. Revista de Biología Tropical, 57(3), 529-539. [ Links ]
Arias-Rodriguez, L., Indy, J. R., Ahumada-Hernández, R. I., Barragán-Cupido, H., Ávalos Lázaro, A. A., & Páramo-Delgadillo, S. (2011). Caracterización cariotípica en mitosis y meiosis del robalo blanco Centropomus undecimalis (Pisces: Centropomidae). Revista de Biología Tropical, 59(2), 683-692. [ Links ]
Badenhorst, D., Stanyon, R., Engstrom, T., & Valenzuela, N. (2013). A ZZ/ZW microchromosome system in the spiny softshell turle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae. Chromosome Research, 21(2), 137-147. [ Links ]
Baker, R. J. & Bickham, J. W. (1986). Speciation by monobrachial centric fusions. Proceedings of the National Academy of Sciences, Unites States of America, 83, 8245-8248. [ Links ]
Bickham, J. W. (1975). A cytosystematic study of turtles in the genera Clemmys, Mauremys and Sacalia. Herpetologica, 31(2), 198-204. [ Links ]
Bickham, J. W. & Baker, R. J. (1976a). Karyotypes of some neotropical turtles. Copeia, 1976(4), 704-707. [ Links ]
Bickham, J. W. & Baker, R. J. (1976b). Chromosome homology and evolution of emydid turtles. Chromosoma, 54(3), 201-219. [ Links ]
Bickham, J. W. & Baker, R. J. (1979). Canalization model of chromosomal evolution. Bulletin of the American Museum of Natural History, 13(1), 70-84. [ Links ]
Bickham, J. W. & Carr, J. L. (1983). Taxonomy and phylogeny of the higher categories of cryptodiran turtles based on a cladistic analysis of chromosomal data. Copeia, 4, 918-932. [ Links ]
Bull, J. J., Moon, R. G., & Legler, J. M. (1974). Male heterogamety in kinosternid turtles (genus Staurotypus). Cytogenetics and Cell Genetics, 13(5), 419-425. [ Links ]
Cabrero, J., Bakkali, M., Bugrov, A., Warchalowska-Sliwa, E., López-León, M. D., Perfectti, F., & Camacho, J. P. M. (2003). Multiregional origin B chromosomes in the grasshopper Eyprepocnemis plorans. Chromosoma, 112, 207-211. [ Links ]
Camacho, J. P. M., Sharbel, T. F., & Beukeboom, L. W. (2000). B-Chromosome evolution. Philosophical Transactions of the Royal Society B: Biological Sciences, 355(1394), 163-178. [ Links ]
Camacho-Mosquera, L. & Amorocho, D. (2008). Caracterización genética de la colonia reproductiva de la tortuga marina golfina Lepidochelys olivácea en el parque nacional natural Gorgona (Pacífico Colombiano) a partir de secuencias de ADN mitocondrial. Boletín de Investigaciones Marinas y Costeras, 37(1), 77-92. [ Links ]
Carr, J. L., Bickham, J. W., & Dean, R. H. (1981). The karyotype and chromosomal banding patterns of the Central American river turtle Dermatemys mawii. Herpetologica, 37(2), 92-95. [ Links ]
Carr, J. L. & Bickham, J. W. (1986). Philogenetic implications of karyotypic variation in the Batagurinae (Testudines: Emydidae). Genetica, 70, 89-106. [ Links ]
Cleiton, F. & Giuliano Caetano, L. (2008). Cytogenetic characterization of two turtle species: Trachemys dorbigni and Trachemys scripta elegans. Caryologia, 61(3), 253-257. [ Links ]
De la Cruz-Pons, A., Zavala-Cruz, J., Guerrero-Peña, A., Salgado-García, S., Lagunes-Espinoza, L. C., & Gavi-Reyes, F. (2012). Metales pesados en suelos cultivados con caña de azúcar en la Chontalpa Tabasco. Universidad y Ciencia, 28(2), 119-130. [ Links ]
De Smet, W. H. O. (1981). The nuclear feulgen-DNA content of the vertebrates (especially reptiles), as measured by fluorescence cytophotometry, with notes on the cell and chromosome size. Acta Zoologica et Pathologica Antverpiensia, 76, 119-167. [ Links ]
Denton, T. E. (1973). Fish chromosome methodology. Chicago, Illinois, Estados Unidos de América: Charles C. Tomas. [ Links ]
Dowler, R. C. & Bickham, J. W. (1982). Chromosomal relationships of the tortoises (family Testudinidae). Genetica, 58, 189-197. [ Links ]
Ferro, J. M., Marti, D. A., Bidau, C. J., Suárez, P., Nagamachi, C. Y., Pieczarka, J. C., & Baldo, D. (2012). B chromosome in the tree frog Hybsiboas albopunctatus (Anura: Hylidae). Herpetologica, 68(4), 482-490. [ Links ]
Fiedler, S., Siebe, C., Herre, A., Roth, B., Cram, S., & Stahr, K. (2009). Contribution of oil industry activities to environmental loads of heavy metals in the Tabasco lowlands, Mexico. Water, Air and Soil Pollution, 197, 35-47. [ Links ]
Forbes, W. R. Jr. (1966). A cytological study of the Chelonia. (Tesis de Doctorado). University of Connecticut. [ Links ]
Frair, W. (1972). Taxonomic relations among chelydrid and kinosternid turtles elucidated by serological tests. Copeia, 1, 97-108. [ Links ]
Gorman, G. C. (1973). The chromosomes of the reptilian, a cytotaxonomic interpretation. In A. B. Chiarelli & E. Capanna (Eds.), Cytotaxonomy and vertebrate evolution (pp. 347 424). New York, USA.: Editorial Academic Press. [ Links ]
Hernández-Guzmán, J., Arias-Rodriguez, L., & Indy, J. R. (2011). Los cromosomas meióticos de la rana arborícola Smilisca baudinii (Anura: Hylidae). Revista de Biología Tropical, 59(1), 355-362. [ Links ]
Hillis, D. M. (1991). The phylogeny of amphibians: Current knowledge and the role of cytogenetics. In D. M. Green & S. K. Sessions (Eds.), Amphibian cytogenetics and evolution (pp. 7-31). San Diego, California, USA: Academic Press Inc. [ Links ]
IUCN. 2012. IUCN red list of threatened species. Retrieved from www.iucnredlist.org [ Links ]
Ivanov, V. G. (1973). Description of the karyotype of Emys orbicularis L. (Testudines, Emydidade). Tsitologiya, 15, 1059-1061. [ Links ]
Jones, R. & Rees, H. (1982). B chromosomes. London, England: Academic. [ Links ]
Jordan, H. E. (1914). Spermatogenesis in Chrysemys marginata and Cistudo carolina. Science, 39, 438. [ Links ]
Kasai, F., O´Brien, P. C., Martin, S., & Ferguson-Smith, M. A. (2012). Extensive homology of chicken macrochromosomes in the karyotypes of Trachemys scripta elegans and Crocodylus niloticus revealed by chromosome painting despite long divergence times. Cytogenetics Genome Research, 136(4), 303-307. [ Links ]
Killebrew, F. C. (1975). Mitotic chromosomes of turtles, III The Kinosternidae. Herpetologica, 31, 398-403. [ Links ]
King, R. C. & Stansfield, W. D. (2002). A dictionary of genetics. USA: Oxford University Press. [ Links ]
Kligerman, A. D. & Bloom, S. E. (1977). Rapid chromosome preparations from solid tissues of fishes. Journal of the Fisheries Research Board of Canada, 34, 266-269. [ Links ]
Kuraku, S., Ishijima, J., Nishida-Umehara, C., Agata, K., Kuratini, S., & Matsuda, Y. (2006). cDNA-based gene mapping and GC3 profiling in the soft-shelled turtle suggest a chromosomal size-dependent GC bias shared by sauropids. Chromosome Research, 14(2), 187-202. [ Links ]
Lee, J. C. (2000). A field guide to the amphibians and reptiles of the Maya World: the lowlands of Mexico, Northern Guatemala, and Belize. USA: Cornell University Press. [ Links ]
Lemos-Espinal, J. A. & Smith, H. M. (2009). Anfibios y reptiles del estado de Chihuahua, México. Universidad Nacional Autónoma de México, University of Colorado at Boulder, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México, D.F. [ Links ]
Levan, A., Fredga, K., & Sandberg, A. A. (1964). Nomenclature for centromeric position on chromosomes. Hereditas, 52, 201-220. [ Links ]
Lockwood, S. F., Seavey, B. T., Dillinger, R. E., & Bickham, J. W. (1991). Variation in DNA content among age classes of broad whitefish (Coregonus nasus) from the Sagavanirktok river delta. Canadian Journal of Zoology, 69, 1335-1338. [ Links ]
López, E. A., Hernández-Fernández, J., & Bernal-Villegas, J. (2008). Condiciones óptimas de cultivo de linfocitos y análisis parcial del cariotipo de la tortuga cabezona, Caretta caretta (Testudines: Cheloniidae) en Santa Marta, Caribe Colombiano. Revista de Biología Tropical, 56(3), 1459-1469. [ Links ]
Martínez, P. A., Boeris, J. M., Sánchez, J., Pastori, M. C., Bolzán, A. D., & Ledesma, M. A. (2009). Karyotypic characterization of Trachemys dorbigni (Testudines: Emydidae) and Chelonoidis (Geochelone) donosobarrosi (Testudines: Testudinidae), two species of Cryptodiran turtles from Argentina. Genetica, 137, 277-283. [ Links ]
Mayer-Goyenechea, I. G. (2001). Sistemática de reptiles y citogenética. Boletín de la Sociedad Herpetológica Mexicana, 9(1), 13-21. [ Links ]
McCoy, C. J. (1982). Reptilia. In S. H. Hurlbert & A. Villalobos-Figueroa (Eds.), Aquatic biota of Mexico, Central America and the West Indies (pp. 515-520). Aquatic Biota-SDSU Foundation. [ Links ]
Mestriner, C. A., Galetti Jr, P. M., Valentini, S. R., Ruiz, I. R. G., Abel, L. D. S., Moreira Filho, O., & Camacho, J. P. M. (2000). Structural and functional evidence that a B chromosome in the characid fish Astyanax scabripinnis is an isochromosome. Heredity, 85, 1-9. [ Links ]
Mittermeier, R. A. & Goettsch, C. (1992). La importancia de la diversidad biológica de México. In J. Sarukhán & R. Dirzo (Eds.), México ante los retos de la biodiversidad. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México D.F. [ Links ]
Moon, R. G. (1974). Heteromorphism in a kinosternid turtle. Mammalian chromosomes newsletter, 15, 10-11. [ Links ]
Olmo, E. (1976). Genome size in some reptiles. Journal of Experimental Zoology, 195, 305-310. [ Links ]
Olmo, E., Capriglione, T., & Odierna G. (2002). Different genomic evolutionary rates in the various reptile lineages. Gene, 295(2), 317-321. [ Links ]
Olmo, E. (2005). Rate of chromosome changes and speciation in reptiles. Genetica, 125, 185-203. [ Links ]
Olmo, E. (2008) Trends in the evolution of reptilian chromosomes. Integrative and Comparative Biology, 48(4), 486-493. [ Links ]
Organ, C. L., Moreno, R. G., & Edwards, S. V. (2008). Three tiers of genome evolution in reptiles. Integrative and Comparative Biology, 48(4), 494-504. [ Links ]
Ortiz, M. L., Rodríguez, P. A., & Bueno, M. L. (2005). Caracterización citogenética de la tortuga sabanera Podocnemis vogli (Reptilia: Testudinata: Podocnemididae). Acta Biológica Colombiana, 10(1), 19-33. [ Links ]
Perfectti, F. & Werren, J. H. (2001). The interspecific origin of B chromosomes: experimental evidence. Evolution, 55(5), 1069-1073. [ Links ]
Porter, C. A., Haiduk, M. W., & De Queiroz, K. (1994). Evolution and phylogenetic significance of ribosomal gene location in chromosomes squamate reptiles. Copeia, 2, 302-313. [ Links ]
Reed, K. M., Hanks, B. G., Bickham, J. W., Rhodin, A. G. J., Greenbaum, I. F., Mittermeier, R. A., & Fedullo, L. P. (1991). Cytogenetic analysis of the pleurodine turtle Phrynops hogei and its taxonomic implications. Amphibia-Reptilia, 12, 203-212. [ Links ]
Risley, P. L. (1936). The chromosomes of the male musk turtle, Sternotherus odoratus L. Cytologia, 7, 232-241. [ Links ]
Rosas, I., Báez, A., & Belmont, R. (1983). Oyster (Crassostrea virginica) as indicator of heavy metal pollution in some lagoons of the Gulf of Mexico. Water, Air and Soil Pollution, 20(1), 127-135. [ Links ]
Schmid, M., Steinlein, C., Bogart, J. P., Feichtinger, W., León, P., La Marca, E., Díaz, L. M., Sanz, A., Chen, S. H., & Hedges, S. B. (2010). The chromosomes of terraranan frogs: Insights into vertebrate cytogenetics. Cytogenetics Genome Research, 130-131(1-8), 1-568. [ Links ]
SEMARNAT. (2010). NORMA Oficial Mexicana NOM- 059-SEMARNAT-2010, Diario Oficial de la Federación, segunda sección, 30 de diciembre del 2010: 1-78. [ Links ]
Sites, J. W. Jr; Bickham, J. W., Haiduk, M. W., & Iverson, J. B. (1979a). Banded karyotypes of six taxa of kinosternid turtles. Copeia, 4, 692-698. [ Links ]
Sites, J. W., Bickham, J. W., & Haiduk, M. W. (1979b). Derived X chromosome in the turtle genus Staurotypus. Science, 206(4425), 1410-1412. [ Links ]
Srikulnath, K., Nishida, C., Matsubara, K., Uno, Y., Thongpan, A., Suputtitada, S., Apisitwanich, S., & Matsuda, Y. (2009). Karyotypic evolution in squamate reptiles: comparative gene mapping revealed highly conserved linkage homology between the butterfly lizard (Leiolepis reevesii rubritaeniata, Agamidae, Lacertilia) and the Japanese four-striped rat snake (Elaphe quadrivirgata , Colubridae, Serpentes). Chromosome Research, 17(8), 975-986. [ Links ]
Tegelström, H. & Ryttman, H. (1981). Chromosomes in birds (Aves): evolutionary implications of macro-and microchromosome numbers and length. Hereditas, 94(1), 225-233. [ Links ]
Tiersch, T. R., Chandler, R. W., Wachtel, S. S. & Elias, S. (1989). Reference standards for flow cytometry and application in comparative studies of nuclear DNA content. Cytometry, 10, 706-710. [ Links ]
Van Brink, J. M. (1959). L´expression morphologique de la digametie chex les sauropsides et les monotremes. Chromosoma, 10(1), 1-72. [ Links ]
Villanueva, S. & Botello, A. V. (1992). Metales pesados en la zona costera del Golfo de México y Caribe mexicano: una revisión. Revista Internacional de Contaminación Ambiental, 8(1), 47-61. [ Links ]
Voss, S. R., Kump, D. K., Putta, S., Pauly, N., Reynolds, A., Henry, R. J., Basa, S., Walker, J. A., & Smith, J. J. (2011). Origin of amphibian and avian chromosomes by fission, fusion, and retention of ancestral chromosomes. Genome Research, 21(8), 1306-1312. [ Links ]
West, R. C., Psuty, N. P., & Thom, B. (1987). Las tierras bajas de Tabasco. Biblioteca Básica Tabasqueña. Villahermosa, Tabasco, México. [ Links ]
*Correspondencia a:
Javier Hernández-Guzmán. División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco, Villahermosa, CP. 86150, Tabasco, México; jhernandez-guzman@hotmail.com
Jeane Rimber Indy. División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco, Villahermosa, CP. 86150, Tabasco, México; jeanerimberindy@yahoo.com
George Shigueki Yasui. Universidade de São Paulo, Faculdade de Zootecnia e Engenharia de Alimentos, Laboratório de Teriogenologia. São Paulo, 13635-900, Brasil; yasui@usp.br
Lenin Arias-Rodriguez. División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco, Villahermosa, CP. 86150, Tabasco, México; leninariasrodriguez@hotmail.com
1. División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco, Villahermosa, CP. 86150, Tabasco, México; jhernandez-guzman@hotmail.com
2. División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco, Villahermosa, CP. 86150, Tabasco, México; jeanerimberindy@yahoo.com
3. Universidade de São Paulo, Faculdade de Zootecnia e Engenharia de Alimentos, Laboratório de Teriogenologia. São Paulo, 13635-900, Brasil; yasui@usp.br
4. División Académica de Ciencias Biológicas, Universidad Juárez Autónoma de Tabasco, Villahermosa, CP. 86150, Tabasco, México; leninariasrodriguez@hotmail.com
Recibido 29-vii-2013. Corregido 13-i-2014. Aceptado 30-i-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}