











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.62 no.2 San José abr./jun. 2014
Feeding and migration habits of white shark Carcharodon carcharias (Lamniformes: Lamnidae) from Isla Guadalupe inferred by analysis of stable isotopes δ15N and δ13C
Hábitos alimenticios y migratorios del tiburón blanco Carcharodon carcharias (Lamniformes: Lamnidae) de Isla Guadalupe inferidos por el análisis de isótopos estables de δ15N and δ13C
Hábitos alimenticios y migratorios del tiburón blanco Carcharodon carcharias (Lamniformes: Lamnidae) de Isla Guadalupe inferidos por el análisis de isótopos estables de δ15N and δ13C
*Dirección para correspondencia:
Abstract
Stable isotope composition of marine top predator’s tissues provides insight information of its trophic ecology and migratory behavior. Previous reports have shown that dermal tissues could record longer patterns of hunting and movement. Based on this, the aim of this study was to describe the feeding and migratory habits of the white shark from Isla Guadalupe, using stable isotopic analysis of dermis. We considered a small subset of many possible prey taxa that the sharks could have eaten throughout their migration: pinnipeds, squid and tuna. We grouped the data in five focal areas: Gulf of California, Coast of California, Isla Guadalupe, SOFA and Hawaii. We performed a Bayesian mixing model to study the trophic ecology of this top predator. Average isotopic values for dermis tissue of white shark were δ13C (-14.5‰) and δ15N (19.1‰). Corrected white shark dermal mean values to resemble muscle were δ13C (-16.6‰) and δ15N (21.2‰). Mixing model data from dermis showed predation in offshore areas such the SOFA and a main importance of pinnipeds as prey of the white shark in Isla Guadalupe. Rev. Biol. Trop. 62 (2): 637-647. Epub 2014 June 01.
Key words: white shark, feeding habits, stable isotopes, mixing models, Isla Guadalupe, Pinnipeds.
Resumen
La composición isotópica de los tejidos de los depredadores tope en el ambiente marino provee información sobre su ecología trófica y su comportamiento migratorio. Estudios previos han mostrado que el tejido dérmico puede registrar patrones largos de movimiento y caza. El objetivo de este estudio fue describir los hábitos tróficos y migratorios de los tiburones blancos de Isla Guadalupe realizando un análisis de isótopos estables de su tejido dérmico. Consideramos un pequeño grupo de muchos posibles taxa que los tiburones pudieron haber comido a lo largo de su migración: pinípedos, calamares y atunes. Estas presas fueron agrupadas en cinco áreas focales: Golfo de California, Isla Guadalupe, Costa de California, SOFA y Hawái. Realizamos un modelo de mezcla bayesiano para estudiar la ecología trófica de este depredador tope. Los promedios del valor isotópico de la dermis del tiburón blanco fueron δ13C (-14.5‰) y δ15N (19.1‰). Los promedios del valor isotópico de la dermis transformada para semejar músculo fueron δ13C (-16.6 ‰) y δ15N (21.2‰). El modelo de mezcla mostró una probable depredación de los tiburones en áreas oceánicas como el SOFA y confirmó la importancia de los pinnípedos como presa principal del tiburón blanco en Isla Guadalupe.
Palabras clave: tiburón blanco, isótopos estables, modelos de mezcla, Isla Guadalupe, pinnípedos.
The white shark Carcharodon carcharias occupies a high trophic level in marine ecosystems (Tricas & McCosker, 1984), and is known to aggregate in offshore and nearshore environments. Sharks tagged at Isla Guadalupe and the Farallon Islands are known to move offshore to an aggregation site called the Shark Offshore Feeding Area (SOFA) between 15° N and 30° N - 130° W and 140° W, near Hawaii (Weng et al., 2007; Domeier & Nasby-Lucas, 2008) and some individuals travel from pelagic areas to the Gulf of California (Domeier, 2012). Stomach content analysis and observational studies have shown that adults mainly feed on marine mammals, especially pinnipeds (Ainley et al., 1981; Morey et al., 2003; Hammerschlag et al., 2006); cetaceans, including dolphins and porpoises (Long & Jones, 1996; Morey et al., 2003; Wcisel et al., 2010), and fishes, including tuna (Fergusson et al., 2000; Ellis & McCosker, 1991). However, those approaches have some limitations that may include an emphasis on recently ingested prey and over representation/retention of prey with hard parts. Field observations at nearshore aggregation sites seem to uphold this perception, while in offshore aggregation sites these are only assumptions based on spatial/temporal patterns of potential prey distribution. For example, at nearshore aggregation sites, there is a high availability of marine mammals. In Isla Guadalupe there is an abundance of the Northern elephant seal Mirounga angustirostris, Guadalupe fur seal Arctocephalus townsendi, and California sea lion Zalophus californianus (Gallo-Reynoso et al., 2004); while in Farallon Islands, there is an abundance of the Northern elephant seal Mirounga angustirostris, and the harbor seal, Phoca vitulina (Ainley et al., 1981). Between spring and summer, the offshore environment in the Central Pacific contains the biggest aggregations of tuna (Okamoto & Bayliff, 2003) so there can be reasonably high numbers near the SOFA. The reported spawning area of Ommastrephid squid overlaps temporally and spatially with white sharks in offshore waters so they may be an important prey resource (Carlisle et al., 2012). Yellowfin tuna, Thunnus albacares are abundant near the Hawaiian Islands during summer months (Boggs & Ito 1993; Itano, 2000) when white sharks are in offshore habitats (Weng et al., 2007; Domeier & Nasby-Lucas, 2008).
Stable isotope analysis (SIA) provides information about assimilated food, diet preferences, and location of foraging in consumers (Schell et al., 1998; Hobson & Sease, 1998; Lesage et al., 2001). The isotopic composition can be used as a natural tag to track movements through isotopically-distinct habitats (Graham et al., 2010). These variations can be recorded in the tissues if the animals move between nearshore/offshore environments. The dietary information retained by tissues of predators depends upon its tissue specific elemental turnover rates, the isotopic composition of food webs used by the consumer, and the prey items in the diet (Tieszen et al., 1983; Hobson & Clark, 1992a). It is commonly accepted that active tissues such as plasma incorporate dietary signatures very rapidly due to high tissue turnover rates; in contrast, less active tissues such as bone and dermis incorporate dietary signatures very slowly and therefore, averages the composition of the assimilated diet over a longer time scale (Hobson & Clark, 1992a). Stable isotope analysis of slow turnover tissues has been used to investigate foraging in nearshore and offshore habitats of different predators, including the harbor seal, California sea lion, Northern elephant seal, and Northern fur seal, Callorhinus ursinus (Burton & Koch, 1999), salmon, Salmo salar (Kennedy et al., 1997), loggerhead sea turtle, Caretta caretta (McClellan et al., 2010), hammerhead shark Sphyrna lewini and dusky shark Carcharhinus obscurus (Hussey et al., 2011). Currently mixing models use stable isotope data to estimate the contribution of sources to a mixture, because stable isotopes are conserved through time (Phillips & Gregg, 2003). Mixing models using a Bayesian framework offer a powerful mean to interpret data because they can incorporate prior information, integrate across sources of uncertainty, and explicitly compare the strength of support for competing models or parameter values (Moore & Semmens, 2008).
Studies employing stable isotope analysis of white shark tissues to understand its feeding ecology are scarce. Some studies focus on ontogenic feeding shifts in vertebrae (Kerr et al., 2006; Estrada et al., 2006; Kim et al., 2012a). Recently one study related the migration and trophic ecology of Northeastern Pacific white sharks with biopsies sampled in California (Carlisle et al., 2012). Shark skin is composed of two collagen regions; the epidermis into which the bases of the denticles protrude, and the dermis, which is a deeper layer of collagen (Motta, 1977). It is feasible that dermis reflects the feeding history of white sharks, and although its turnover rate remains unknown; if the dermis has a slow turnover rate it would reflect a longer term diet integration.
White sharks aggregating at Isla Guadalupe and the coasts of California may be a single population (Jorgensen et al., 2009), this supports the idea that both have similar feeding habits (Carlisle et al., 2012) and they appear to have similar spatial and temporal behavior patterns (Domeier & Nasby-Lucas, 2008). Acoustic tags of some sharks that traveled from Central California to Isla Guadalupe showed some connectivity between these areas (Jorgensen et al., 2012). In California, white sharks are known to aggregate at pinnipeds rookeries during autumn and winter (September-February), when the abundance of immature elephant seals is at a maximum (Ainley et al., 1985); the same seasonal behavior was reported for white sharks at Isla Guadalupe (Domeier & Nasby-Lucas 2008). The aim of this study was to describe the feeding and migratory habits of white shark from Isla Guadalupe using stable isotopic analysis of dermis. For this we determined the extent of foraging in offshore vs. nearshore areas using a dietary mixing model that incorporated different tissue incorporation rates and satellite tagging data.
Materials and methods
White sharks dermis biopsies were obtained off Isla Guadalupe from September through December 2007 (N=15) and during September through December 2010 (N=16). We used a pole spear with steel tips to take biopsies in situ. Steel typed dart measuring 10cm length and 0.6cm in diameter with tapering edges based on a cutting head originally designed by Winn et al. (1973). Sharks were attracted with bait. In some cases, sharks were photo-identified with an underwater camera to determine sex and size. Isotope signatures of probable white shark prey within its migration range in the Eastern Pacific were obtained from field surveys at Isla Guadalupe and a literature review. During the same years we collected skin and hair samples of Northern elephant seal (N=32), California sea lion (N=4) and Guadalupe fur seal (N=17). We used a crossbow and aluminum arrows with steel tips to obtain samples from their backs. All samples were frozen at -80°C until analyzed.
Samples of white sharks were rinsed with deionized water in order to remove urea. The lipids were removed by rinsing the ground tissue several times with 2:1 chloroform: methanol mixture, following the method of Folch et al., 1957. Pinniped samples were rinsed with distilled water to eliminate salt and sand residues. The samples were washed again with distilled water and dried at 80°C for 12h to eliminate excessive moisture. Samples were put in flasks with Teflon caps and lyophilized. Subsamples of ~0.001g were encapsulated in tin capsules (8mm×5mm). Isotope values of carbon (C) and nitrogen (N) were obtained by mass spectrometry at the Continuous Flow-Isotope Ratio Mass Spectrometry Laboratory at the University of California, Santa Cruz. Isotope signatures were reported as delta values (δX), where X is the heavy isotope with reference to the standard, expressed as parts per thousand (‰). These data were calculated according to the formula:
δX=(Rsample/Rstandard-1)×1000,
where Rsample is the ratio between the heavy isotope and the light isotope and Rstandard is the standard for the isotopes. The standard for δ13C was carbon dioxide from calcium carbonate produced by the Cretaceous belemnoid fossil Belemnitella americana (PDB, Pee Dee Belemnite). The standard for δ15N was the atmospheric nitrogen.
Lipid content can alter δ13C values if the C: N ratio for samples is greater than 3.5 (Post et al., 2007). The C: N ratio for epidermis of Guadalupe pinnipeds was ~5.1, therefore lipid correction was necessary. To account lipid effects over the isotopic value of pinnipeds, we performed a mathematical normalization of δ13C to simplify sample preparation following the equations proposed by Post et al. (2007) for terrestrial mammals.
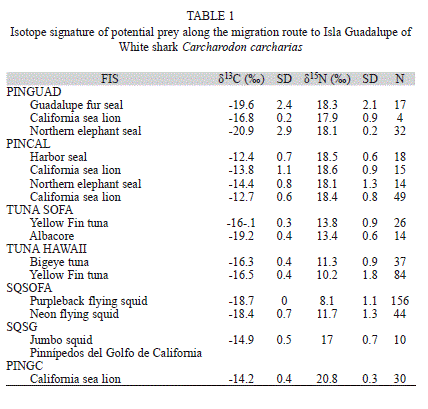
From the literature, we obtained stable isotopic signatures available of the potential prey items that white sharks of Isla Guadalupe could predate along its migratory route. We obtained signatures of harbor seals, California sea lions and Northern elephant seals (Burton & Koch, 1999). California sea lion from the coast of Baja California (Newsome et al., 2006).Yellow fin tuna, bigeye tuna Thunnus obesus from Hawaii (Graham et al., 2007; Carlisle et al., 2012). Yellow fin tuna and albacore Thunnus alalunga values near the SOFA (Olson et al., 2010; Carlisle et al., 2012). Purpleback flying squid Sthenoteuthis oualaniensis, neon flying squid Ommastrephes bartrami values near the SOFA (Carlisle et al., 2012).
Due to the lack of isotopic information, we only considered a small subset of many possible prey taxa that the sharks could have eaten throughout their migration. Also, data from stomach contents are not available for this population of sharks; so we used the prey isotope data grouped by region rather than by species. We grouped the data in five focal areas: Gulf of California, Coast of California, Isla Guadalupe, SOFA and Hawaii. We used mean values as an estimate of generalized prey isotopic composition for each region.
Substantial proportions of the long-term diet determined from shark tissue are likely provided from several regions by different prey. There are distinct regionally-specific isotope values, but very similar isotope values within those regions (West et al., 2010). We used pinnipeds as indicators of nearshore environments because they are the principal prey of white sharks in Central California and Isla Guadalupe (Long et al., 1996; Gallo-Reynoso et al., 2004; Hoyos-Padilla, 2009) and once sharks return to coastal aggregation sites they rapidly gain mass by feeding on pinnipeds (Chapple et al., 2011). We used tunas and squid as indicators of offshore environments of Hawaii and pelagic habitats near the SOFA. Hence, isotopic signatures of Isla Guadalupe pinnipeds, California pinnipeds, Isla Guadalupe pinnipeds, tuna and squid from the SOFA, and tuna from Hawaii were averaged to obtain unique values that represent each feasible isotope source (FIS).
To compare the prey values of the different tissue types, all of which have very different discrimination factors, we standardized them to muscle values. We adjusted the pinniped’s skin δ13C values to resemble muscle by subtracting 1.5‰, and fur δ13C values to resemble muscle by subtracting 1.4‰ (Hobson et al., 1996). The isotopic signatures of feasible prey at Isla Guadalupe and those obtained from the literature are listed in table 1.
Because the stable isotope ratio for an animal’s tissue (δtissue) is directly related to that of its diet (δdiet) as δtissue=δdiet+Δdt (DeNiro & Epstein, 1981), we used this equation and the white shark dermis values of C and N (δXtissue) to estimate the corresponding δdiet values. In order to compare our results with the study of Carlisle we used discrimination factors from leopard shark, Triakis semifasciata; 3.7‰±0.4 SD for nitrogen, 1.7‰±0.5 SD for carbon (Carlisle et al., 2012, Kim et al., 2012b).
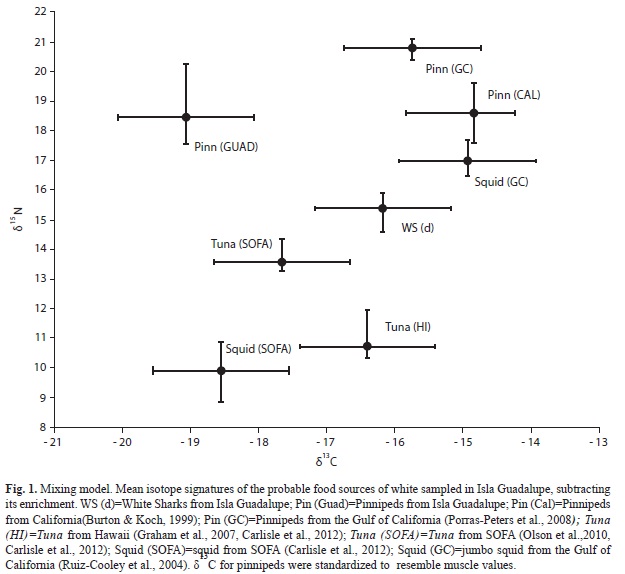
We plotted the isotopic values δ15N versus δ13C of each feasible isotope source with the white shark δdiet values subtracting its enrichment (Fig. 1). To determine the probable contribution of each feasible isotope source to shark tissue, we performed a Bayesian mixing models approach, with the software package SIAR (Stable Isotope Analysis in R; Parnell et al., 2010). The SIAR model is fit via Markov Chain Monte Carlo (MCMC) methods producing simulations of plausible values of dietary proportions of sources consistent with the data using a Dirichlet prior distribution (Jackson et al., 2009, Parnell et al., 2010). The SIAR MCMC were run for 500 000 iterations, the resulting probability density function distributions of the feasible foraging solutions produced by SIAR allowed direct identification of the most probable solution (i.e., the median value).
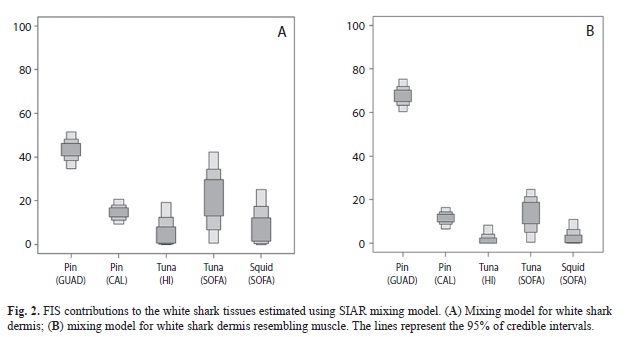
As most of the sharks migrate from Isla Guadalupe to SOFA we developed a mixing model of dermis and dermis resembling muscle only with the FIS from the Northeast Pacific. In order to compare the isotopic values with those of other studies, dermal isotope values were corrected to resemble muscle according to tissue-specific differences following the methods of Carlisle et al., (2012). However it has been demonstrated that some sharks that enter the Gulf of California come from Isla Guadalupe (Domeier, 2012). We developed a mixing model of dermis and dermis resembling muscle with the FIS of the Northeast Pacific plus the FIS of the Gulf of California. White sharks have been spotted near the colonies of California sea lions at San Pedro Mártir and San Pedro Nolasco Islands (Aurioles & Zavala, 1994), which is why we chose isotopic values of California sea lion pups from these two islands (Porras-Peters et al., 2008). Large concentrations of jumbo squid Dosidicus gigas in the Gulf of California occur during April and May (Morales-Bojórquez et al., 2012). This period of big squid concentration coincides with the occurrence of white sharks in the Gulf of California (Galván-Magaña et al., 2011; Domeier, 2012). We used the stable isotopic values available of jumbo squid published for the Gulf of California (Ruiz-Cooley et al., 2004).
Results
We obtained 28 values of δ13C and δ15N from the 31 sharks sampled. Using photo identification we identified 13 adults (3.5-5m total length) and 1 subadult (2.5m total length). We only identified 10 males and 6 females. Female size average was 4.2m and male size average was 3.6m.
The C:N ratio of dermal tissues of white shark was lower than 3.5 (2.5±0.08‰, n=33, mean±SD), hence the lipid content was minimal. The average isotopic value for dermal tissue was -14.4‰ (SD=0.5, n=28) for δ13C and 19.1‰ (SD=0.7, n=28) for δ15N. The corrected white shark dermal mean value to resemble muscle was: -16.6‰ (SD=0.5, n=28) for δ13C, and 21.2‰ (SD=0.9, n=28) for δ15N. There were no significant differences between males and females for δ13C (ANOVA, F4.747=0.1, p>0.05) and for δ15N (ANOVA, F4.747=0.4, p>0.05).
There were no significant differences in δ13C and δ15N values between pinniped skin and fur (ANOVA, F4.747=0.4, p>0.05; ANOVA, F5.317=1.6, p>0.05 respectively). Corrected epidermal main values of Guadalupe pinnipeds were: -19.06‰ (SD=0.9, n=89) for δ13C and 18.45‰ (SD=1.8, n=89 for δ13N. Corrected bone main values of California pinnipeds were: -15.98‰ (SD=1.0, n=96) for δ13C and 18.60 ‰ (SD=0.6, n=96) for δ13N. Corrected fur main values of Gulf of California pinnipeds were: -14.23 ‰ (SD=0.4, n=30) for δ13C and 20.80‰ (SD=0.3, n=30) for δ13N.
Pinnipeds had the highest δ13N values while tuna had the lowest. Squid of the Gulf of California showed higher δ13N values than squid of the SOFA. Pinnipeds and squid from the Gulf of California were more enriched in δ13C than pinnipeds of Isla Guadalupe and squid of the SOFA. Squid of the Gulf of California had higher δ13N values than squid of the SOFA. Tuna of Hawaii were more enriched in δ13C than tuna of the SOFA.
The range of the probable contribution of the FIS to white shark dermis is shown in table 2. Pinnipeds of Isla Guadalupe and Tuna of the SOFA were the two main FIS for white shark dermis (Fig. 2A). For dermis resembling muscle, pinnipeds of Isla Guadalupe showed a great feasible contribution of 60-77% while the other FIS had a contribution less than 30% (Fig. 2B). Incorporating the FIS of the Gulf of California the probable contribution changed. For dermis, Jumbo squid showed the greatest contribution (Fig. 3.A) while Pinnipeds of Isla Guadalupe remained the greatest contribution to the dermis resembling muscle (Fig. 3B).
Discussion
The contribution of the tuna from the SOFA of the white shark dermis samples collected from Isla Guadalupe is very similar to that of the shark dermis samples collected near the Coast of California (Carlisle et al. 2012). This implies that the trophic habits of the white sharks of Isla Guadalupe are similar to those of the sharks of California in the Northeast Pacific.
The mixing model of the dermis showed that predation in offshore areas is important in the shark’s long-term diet. These findings are consistent with a recent study of Northeastern Pacific white sharks (Carlisle et al., 2012). Near the SOFA, abundance of tuna peaks in the winter and spring, the same seasons when the white sharks are present there (Okamoto & Bayliff, 2003; Weng et al., 2007). The contribution to the dermis of the white sharks of tuna from Hawaii and pinnipeds from Isla Guadalupe suggests displacement between nearshore and offshore environments. Sharks migrating to Hawaii remained near the islands for up to 122 days, potentially feeding on pelagic fishes and marine mammals that are concentrated around the islands (Weng et al., 2007). Some studies with mathematical models propose that white shark tissues need approximately 40 to 50 years to reach isotopic equilibrium to a new diet. Adult white sharks would probably reach their maximum estimated longevity before reaching such isotopic equilibrium (Bruce, 1992; Malpica-Cruz, 2012). Under this assumption dermal collagen of white sharks incorporates dietary signatures slowly; hence white shark dermis has the possibility to record longer patterns of hunting and movement.
The mixing model of dermis and dermis resembling muscle without the Gulf of California FIS reflects the importance of pinnipeds as prey in Isla Guadalupe. Movements of white sharks in Isla Guadalupe are potentially associated with foraging and the seasonal cycles of pinnipeds (Domeier et al., 2012), specifically the Northern elephant seals (Hoyos-Padilla, 2009). White sharks in Isla Guadalupe behead pups of Guadalupe fur seals and ambush Northern elephant seals in deep areas (Gallo-Reynoso et al., 2004, Hoyos-Padilla, 2009). Our results confirm that pinnipeds of Isla Guadalupe are an important prey item for white sharks. The white shark has an ecological role that links food webs in Pacific offshore and nearshore areas. Results of the mixing model show a potential food assimilation index (Ben-David & Schell, 2001). Both environments are part of the trophic niche of this top predator. However, to determine if shark dermis mathematically converted to muscle can be used to understand the trophic ecology of the white shark, muscle biopsies are needed to corroborate previous assumptions.
Mixing model of dermis and dermis resembling muscle with the FIS of the Northeast Pacific plus the FIS of the Gulf of California reflects the importance of squid as prey and supports the evidence that some sharks from Isla Guadalupe move into the Gulf of California (Domeier, 2012). This evidence could be linked to white shark trophic ecology. The isotopic contribution of the California sea lions from these zones to the white sharks dermis was lower than jumbo squid. Therefore, the latter can be considered a main prey item there.
Large cephalopods are an important diet component of white sharks and also are indicators of habitat use (Smale & Cliff, 2012). For example, jumbo squid was found as a prey item in the stomach of a 4.6m female caught near California (Ellis & McCosker, 1991). Moreover, jumbo squid recruitment in the Gulf of California occurs during April and May and its population size in April is estimated at ~136 million squid (Morales-Bojórquez et al., 2012).This is the same season when most white sharks have been caught in the gulf (Galván- Magaña et al., 2011). Therefore, there is a high probability that white sharks could be feeding on squid. We infer that the high contribution of jumbo squid from the Gulf of California in the dermis of the white shark could show the trophic use of the Gulf by some white sharks that inhabit waters around Isla Guadalupe; however dermal and muscle biopsies from Gulf of California populations are needed to support this assumption.
Acknoledgements
Funding was provided by the WWF/Telcel Aliance, PROMEXICO, project “Movements of white sharks in Isla Guadalupe” (Project KZ85), Instituto Politécnico Nacional (COFAA, EDI), Centro de Investigaciones Biológicas del Noroeste (CIBNOR) and CONACYT. Special thanks to Guillermo García-Cortés and Jorge Cobos-Anaya for the manufacture of the darts.
References
Ainley, D. G., Strong, C. S. Huber, H. R. Lewis, T. J., & Morrell, S. H. (1981). Predation by sharks on pinnipeds at the Farallon Islands. Fishery Bulletin, 78, 941-945. [ Links ]
Ainley, D. G., Henderson, R. P., Huber, H. R., Boekelheide, R. J., Allen, S. G., & McElroy, T. L. (1985). Dynamics of white shark/pinniped interactions in the Gulf of the Farallones. Memoirs of the California Academy of Sciences, 9, 109-122. [ Links ]
Aurioles-Gamboa, D., & Zavala-González, A. (1994). Algunos factores ecológicos que determinan la distribución y abundancia del lobo marino Zalophus californianus, en el Golfo de California. Ciencias Marinas, 20, 535-553. [ Links ]
Ben-David, M., & Schell, D. M. (2001). Mixing models in analyses of diet using multiple stable isotopes: a response. Oecologia, 127,180-184. [ Links ]
Bruce, B. (1992). Preliminary observations on the biology of the White Shark, Carcharodon carcharias, in South Australian waters. Marine & Freshwater Research, 43, [ Links ] 1-11.
Boggs, C. H., & Ito, R. Y. (1993). Hawaii’s pelagic fisheries. Marine Fisheries Review, 55, 69-82. [ Links ]
Burton, R. K., & Koch, P. L. (1999). Isotopic tracking of foraging and long-distance migration in northeastern Pacific pinnipeds. Oecologia, 119, 578-585. [ Links ]
Carlisle, A. B., Kim, S. L., Semmens, B. X., Madigan, D. J., Jorgensen, S. J., Perle, C. R., Anderson, S. D., Chapple, T. K., Kanive, P. E., & Block, B. A. (2012). Using Stable Isotope Analysis to Understand the Migration and Trophic Ecology of Northeastern Pacific White Sharks (Carcharodon carcharias). PLoS ONE, 7(2):e30492. [ Links ]
Chapple, T. K., Jorgensen, S. J., Anderson, S. D., Kanive, P. E., Klimley, A. P., Botsford, L. W., & Block, B. A. (2011). A first estimate of white shark, Carcharodon carcharias, abundance off Central California. Biology Letters, 7, 581-583. [ Links ]
DeNiro, M. J., & Epstein, S. (1981). Influence of the diet on the distribution of nitrogen isotopes in animals. Geochimica et Cosmochimica Acta, 45, 341-351. [ Links ]
Domeier, M., & Nasby-Lucas, N. (2008). Migration patterns of white sharks Carcharodon carcharias tagged at Isla Guadalupe, Mexico, and identification of an eastern Pacific shared offshore foraging area. Marine Ecology Progress Series, 370, 221-237. [ Links ]
Domeier, M. L. (2012). A new Life-History Hypothesis for White Sharks, Carcharodon carcharias, in the Northeastern Pacific. In M. L. Domeier (Eds.), Global Perspectives on the Biology and Life History of the White Shark (pp. 199-223). Florida: CRC Press. [ Links ]
Ellis, R., & McCosker, J. E. (1991). Great White Shark. Stanford: Stanford University Press. [ Links ]
Estrada, J. A., Rice, A. N., Natanson, L. J., & Skomal, G. B. (2006). Use of isotopic analysis of vertebrae in reconstructing ontogenetic feeding ecology in white sharks. Ecology, 87, 829-834. [ Links ]
Fergusson, I. K., Compagno, L. J. V., & Marks, M. A. (2000). Predation by White Sharks; Carcharodon carcharias; (Chondrichthyes: Lamnidae) Upon Chelonians, with New Records from the Mediterranean Sea and a First Record of the Ocean Sunfish; Mola mola; (Osteichthyes: Molidae) as Stomach Contents. Environmental Biology of Fishes, 5, 447-453. [ Links ]
Folch, J., Lees, M., & Stanley, G. H. (1957). A simple method for the isolation and purification of total lipids from animal tissues. The Journal of Biological Chemistry 226, 497-509. [ Links ]
Gallo-Reynoso, J. P., Figueroa-Carranza, A. L., & Blanco-Parra, M. P. (2004). Shark wounds in the seals of Isla Guadalupe, México. Annual Meeting of the American Elasmobranchs Society. Norman, Oklahoma USA. [ Links ]
Galván-Magaña, F., Hoyos-Padilla, E. M., Navarro-Serment, C. J., & Márquez-Farías, F. (2011). Records of white shark, Carcharodon carcharias, in the Gulf of California, Mexico. Marine Biology Records, 3, e111. [ Links ]
Graham, B. S., Grubbs, D., Holland, K., & Popp, B. (2007). A rapid ontogenetic shift in the diet of juvenile yellowfin tuna from Hawaii. Marine Biology, 150, 647-658. [ Links ]
Graham, B. S., Koch, P. L., Newsome, S. D., McMahon, K. W., & Aurioles, D. (2010). Using isoscapes to trace the movements and foraging behavior of top predators in oceanic ecosystems. In J. B. West, G. Bowen, T. Dawson, & K. Tu (Eds.), Isoscapes: understanding movement, pattern, and process on earth through isotope mapping. (pp. 299-318). New York: Springer-Verlag. [ Links ]
Hammerschlag, N., Martin, R. A., & Fallows, C. (2006). Effects of environmental conditions on predator-prey interactions between white sharks (Carcharodon carcharias) and Cape fur seals (Arctocephalus pusillus pusillus) at Seal Island, South Africa. Environmental Biology of Fishes, 76, 341-350. [ Links ]
Hobson, K. A., & Clark, R. G. (1992a). Assessing Avian Diets Using Stable Isotopes I: Turnover of 13C in Tissues. The Condor, 94, 181-188. [ Links ]
Hobson, K. A., Schell, D. M., Renouf, D., & Noseworthy, E. (1996). Stable carbon and nitrogen fractionation between diet and tissues of captive seals: Implications for dietary reconstructions involving marine mammals. Canadian Journal of Fisheries and Aquatic Sciences, 53, 528-533. [ Links ]
Hobson, K. A., & Sease, J. L. (1998). Stable isotope analyses of tooth annuli reveal temporal dietary records: an example using Steller sea lions. Marine Mammal Science, 14, 116-129. [ Links ]
Hoyos-Padilla, E. M. (2009). Patrones de movimiento del tiburón blanco (Carcharodon carcharias) en isla Guadalupe, México (Tesis de doctorado, CICIMAR, IPN). [ Links ]
Hussey, N. E., Dudley, S. F. J., McCarthy, I. D., Geremy, C., & Fisk, A. T. (2011). Stable isotope profiles of large marine predators: viable indicators of trophic position, diet, and movement in sharks. Canadian Journal of Fisheries and Aquatic Sciences, 68, 2029-2045. [ Links ]
Itano, D. G. (2000). The reproductive biology of yellowfin tuna (Thunnus albacares) in Hawaiian waters and the western tropic Pacific Ocean: project summary. SOEST 00-01 JIMAR Contribution 00-328. Pelagic Fisheries Research Program, JIMAR, University of
Hawaii. [ Links ]
Jackson, A. L., Inger, R., Bearhop, S., & Parnell, A. (2009). Erroneous behaviour of MixSIR, a recently published Bayesian isotope mixing model: a discussion of Moore & Semmens (2008). Ecology Letters, 12, E1-E5. [ Links ]
Jorgensen, S. J., Reeb, C. A., Chapple, T. K., Anderson, S., Perle, C., Van Sommeran, S. R., Fritz-Cope, C., Brown, A. C., Klimley, A. P., & Block, B. A. (2009). Philopatry and migration of Pacific white sharks. Proceedings of the Royal Society B: Biological Sciences, 277, 679-688. [ Links ]
Jorgensen, S. J., Chapple, T. K., Anderson, S., Hoyos, M., Reeb, C., & Block, B. (2012). Connectivity among White Shark Coastal Aggregation Areas in Northeastern Pacific. In M. L. Domeier (Ed.), Global Perspectives on the Biology and Life History of the White Shark (pp. 159-167). Florida: CRC Press. [ Links ]
Kennedy, B. P., Blum, J. D., Folt, C. L., & Nislow, K. H. (1997). Natural isotope markers in salmon. Nature, 387, 766-767. [ Links ]
Kerr, L. A., Andrews, A. H., Cailliet, G. M., Brown, T. A., & Coale, K. H. (2006). Investigations of Δ14C, δ13C, and δ15N in vertebrae of white shark (Carcharodon carcharias) from the eastern North Pacific Ocean. Environmental Biology of Fishes,77, 337-353. [ Links ]
Kim, S. L., Tinker, T., Estes, J. A., & Koch, P. L. (2012a). Ontogenetic and Among-Individual Variation in Foraging Strategies of Northeast Pacific White Sharks Based on Stable Isotope Analysis. PLoS ONE, 7(9), e45068. [ Links ]
Kim, S. L., del Rio, C. M. N., Casper, D., & Koch, P. L. (2012b). Isotopic incorporation rates for shark tissues from a long-term captive feeding study. The Journal of Experimental Biology, 215, 2495-2500. [ Links ]
Lesage, V., Hammill, M. O., & Kovacs, K. M. (2001). Marine mammals and the community structure of the estuary and gulf of St Lawrence, Canada: evidence from stable isotope analysis. Marine Ecology Progress Series, 210, 203-221. [ Links ]
Long, J. D., & Jones, R. (1996). White Shark Predation and Scavening on Cetaceans in the Eastern North Pacific Ocean. In A. P. Klimley & D. G. Ainley (Eds.), Great white sharks: the biology of Carcharodon carcharias (pp. 161-177). San Diego: Academic Press. [ Links ]
Malpica-Cruz, L., Herzka, S. Z., Sosa-Nishizaki, O., Lazo, J. P., & Trudel, M. (2012). Tissue-specific isotope trophic discrimination factors and turnover rates in a marine elasmobranch: empirical and modeling results. Canadian Journal of Fisheries and Aquatic Sciences, 69, 551-564. [ Links ]
McClellan, C. M., Braun-McNeill, J., Avens, L., Wallace, B. P., & Read, A. J. (2010). Stable isotopes confirm a foraging dichotomy in juvenile loggerhead sea turtles. Journal of Experimental Marine Biology and Ecology, 387, 44-51. [ Links ]
Morales-Bojórquez, E., Hernández-Herrera, A., Nevárez-Martínez, M. O., & Díaz-Uribe, J. G. (2012). Population size of the jumbo squid Dosidicus gigas in the central Gulf of California, Mexico, based on markrecapture data. Aquatic Biology, 15, 27-34. [ Links ]
Morey, G., Martínez, M., Massutí, E., & Moranta, J. (2003). The Occurrence of White Sharks, Carcharodon carcharias, Around the Balearic Islands (Western Mediterranean Sea). Environmental Biology of Fishes, 68, 425-432. [ Links ]
Motta, Ρ. J. (1977). Anatomy and functional morphology of dermal collagen fibres in shark. Copeia, 3, 454-464. [ Links ]
Moore, J. W., & Semmens, B. X. (2008). Incorporating uncertainty and prior information into stable isotope mixing models. Ecology Letters, 11, 470-480. [ Links ]
Newsome, S. D., Koch, P. L., Etnier, M. A., & Aurioles-Gamboa, D. (2006). Using carbon and nitrogen isotope values to investigate maternal strategies in northeast Pacific otariids. Marine Mammal Science, 22, 556-572. [ Links ]
Okamoto, H., & Bayliff, W. H. (2003). A review of the Japanese longline fishery for tunas and billfishes in the eastern Pacific Ocean, 1993-1997. Inter-american tropical tuna commission bulletin, 22, 221-431. [ Links ]
Olson, R. J., Popp, B. N., Graham, B. S., Lopez-Ibarra, G. A., Galván-Magaña, F., Lennert-Cody, C. E., Bocanegra-Castillo, N., Wallsgrove, N. J., Gier, E., Alatorre-Ramirez, V., Balance, L. T., & Fry, B. (2010). Food-web inferences of stable isotope spatial patterns in copepods and yellowfin tuna in the pelagic eastern Pacific Ocean. Progress in Oceanography, 86, 124-138. [ Links ]
Parnell, A. C., Inger, R., Bearhop, S., & Jackson, A. L. (2010). Source partitioning using stable isotopes: coping with too much variation. PLoS ONE, 5, e9672. [ Links ]
Phillips, D. L., & Gregg, J. W. (2003). Source partitioning using stable isotopes: coping with too many sources. Oecologia, 136, 261-269. [ Links ]
Porras-Peters, H., Aurioles-Gamboa, D., Cruz-Escalona, V. H., & Koch, P. L. (2008). Trophic level and overlap of sea lions (Zalophus californianus) in the Gulf of California, Mexico. Marine Mammal Science, 24, 554-576. [ Links ]
Post, D., Layman, C., Arrington, D., Takimoto, G., Quattrochi, J., & Montaña, C. (2007). Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia, 152, 179-189. [ Links ]
Ruiz-Cooley, R. I., Gendron, D., Aguiñiga, S., Mesnick, S., & Carriquiry, J. D. (2004). Trophic relationships between sperm whales and jumbo squid using stable isotopes of C and N. Marine Ecology Progress Series, 277, 275-283. [ Links ]
Schell, D. M., Barnett, B. A., & Vinette, K. A. (1998). Carbon and nitrogen isotope ratios in zooplankton of the Bering, Chukchi and Beaufort seas. Marine Ecology Progress Series, 162, 11-23. [ Links ]
Smale, M. J., & Cliff, G. (2012). White sharks and Cephalopod Prey. Indicators of Habitat use? In M.L Domeier (Ed.), Global Perspectives on the Biology and Life History of the White Shark (pp. 51-57). Florida: CRC Press. [ Links ]
Tieszen, L. L., Boutton, T. W., Tesdahl, K. G., & Slade, N. A. (1983). Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet. Oecologia, 57, 32-37. [ Links ]
Tricas, T. C., & McCosker, J. E. (1984). Predatory behavior of the white shark, Carcharodon carcharias, with notes on its biology. Proceedings of California Academy Science, 43, 221-238. [ Links ]
Wcisel, M., Chivelle, W., & Gottfried, M. D. (2010). A potential predation attempt by a great white shark on an Indo-Pacific humpback dolphin. South African Journal of Wildlife Research 40, 184-187. [ Links ]
Weng, K., Boustany, A., Pyle, P., Anderson, S., Brown, A., & Block, B. (2007). Migration and habitat of white sharks (Carcharodon carcharias) in the eastern Pacific Ocean. Marine Biology 152, 877-894. [ Links ]
West, A. G., Goldsmith, G., Brooks, P., & Dawson, T. (2010). Discrepancies between isotope ratio infrared spectroscopy and isotope ratio mass spectrometry for the stable isotope analysis of plant and soil waters. Rapid Communications in Mass Spectrometry 24, 1948-1954. [ Links ]
Winn, H. E., Bishoff, W. L., & Tarusky, A. G. (1973). Cytological sexing of Cetacea. Marine Biology 23: 343-346. [ Links ]
Ainley, D. G., Henderson, R. P., Huber, H. R., Boekelheide, R. J., Allen, S. G., & McElroy, T. L. (1985). Dynamics of white shark/pinniped interactions in the Gulf of the Farallones. Memoirs of the California Academy of Sciences, 9, 109-122. [ Links ]
Aurioles-Gamboa, D., & Zavala-González, A. (1994). Algunos factores ecológicos que determinan la distribución y abundancia del lobo marino Zalophus californianus, en el Golfo de California. Ciencias Marinas, 20, 535-553. [ Links ]
Ben-David, M., & Schell, D. M. (2001). Mixing models in analyses of diet using multiple stable isotopes: a response. Oecologia, 127,180-184. [ Links ]
Bruce, B. (1992). Preliminary observations on the biology of the White Shark, Carcharodon carcharias, in South Australian waters. Marine & Freshwater Research, 43, [ Links ] 1-11.
Boggs, C. H., & Ito, R. Y. (1993). Hawaii’s pelagic fisheries. Marine Fisheries Review, 55, 69-82. [ Links ]
Burton, R. K., & Koch, P. L. (1999). Isotopic tracking of foraging and long-distance migration in northeastern Pacific pinnipeds. Oecologia, 119, 578-585. [ Links ]
Carlisle, A. B., Kim, S. L., Semmens, B. X., Madigan, D. J., Jorgensen, S. J., Perle, C. R., Anderson, S. D., Chapple, T. K., Kanive, P. E., & Block, B. A. (2012). Using Stable Isotope Analysis to Understand the Migration and Trophic Ecology of Northeastern Pacific White Sharks (Carcharodon carcharias). PLoS ONE, 7(2):e30492. [ Links ]
Chapple, T. K., Jorgensen, S. J., Anderson, S. D., Kanive, P. E., Klimley, A. P., Botsford, L. W., & Block, B. A. (2011). A first estimate of white shark, Carcharodon carcharias, abundance off Central California. Biology Letters, 7, 581-583. [ Links ]
DeNiro, M. J., & Epstein, S. (1981). Influence of the diet on the distribution of nitrogen isotopes in animals. Geochimica et Cosmochimica Acta, 45, 341-351. [ Links ]
Domeier, M., & Nasby-Lucas, N. (2008). Migration patterns of white sharks Carcharodon carcharias tagged at Isla Guadalupe, Mexico, and identification of an eastern Pacific shared offshore foraging area. Marine Ecology Progress Series, 370, 221-237. [ Links ]
Domeier, M. L. (2012). A new Life-History Hypothesis for White Sharks, Carcharodon carcharias, in the Northeastern Pacific. In M. L. Domeier (Eds.), Global Perspectives on the Biology and Life History of the White Shark (pp. 199-223). Florida: CRC Press. [ Links ]
Ellis, R., & McCosker, J. E. (1991). Great White Shark. Stanford: Stanford University Press. [ Links ]
Estrada, J. A., Rice, A. N., Natanson, L. J., & Skomal, G. B. (2006). Use of isotopic analysis of vertebrae in reconstructing ontogenetic feeding ecology in white sharks. Ecology, 87, 829-834. [ Links ]
Fergusson, I. K., Compagno, L. J. V., & Marks, M. A. (2000). Predation by White Sharks; Carcharodon carcharias; (Chondrichthyes: Lamnidae) Upon Chelonians, with New Records from the Mediterranean Sea and a First Record of the Ocean Sunfish; Mola mola; (Osteichthyes: Molidae) as Stomach Contents. Environmental Biology of Fishes, 5, 447-453. [ Links ]
Folch, J., Lees, M., & Stanley, G. H. (1957). A simple method for the isolation and purification of total lipids from animal tissues. The Journal of Biological Chemistry 226, 497-509. [ Links ]
Gallo-Reynoso, J. P., Figueroa-Carranza, A. L., & Blanco-Parra, M. P. (2004). Shark wounds in the seals of Isla Guadalupe, México. Annual Meeting of the American Elasmobranchs Society. Norman, Oklahoma USA. [ Links ]
Galván-Magaña, F., Hoyos-Padilla, E. M., Navarro-Serment, C. J., & Márquez-Farías, F. (2011). Records of white shark, Carcharodon carcharias, in the Gulf of California, Mexico. Marine Biology Records, 3, e111. [ Links ]
Graham, B. S., Grubbs, D., Holland, K., & Popp, B. (2007). A rapid ontogenetic shift in the diet of juvenile yellowfin tuna from Hawaii. Marine Biology, 150, 647-658. [ Links ]
Graham, B. S., Koch, P. L., Newsome, S. D., McMahon, K. W., & Aurioles, D. (2010). Using isoscapes to trace the movements and foraging behavior of top predators in oceanic ecosystems. In J. B. West, G. Bowen, T. Dawson, & K. Tu (Eds.), Isoscapes: understanding movement, pattern, and process on earth through isotope mapping. (pp. 299-318). New York: Springer-Verlag. [ Links ]
Hammerschlag, N., Martin, R. A., & Fallows, C. (2006). Effects of environmental conditions on predator-prey interactions between white sharks (Carcharodon carcharias) and Cape fur seals (Arctocephalus pusillus pusillus) at Seal Island, South Africa. Environmental Biology of Fishes, 76, 341-350. [ Links ]
Hobson, K. A., & Clark, R. G. (1992a). Assessing Avian Diets Using Stable Isotopes I: Turnover of 13C in Tissues. The Condor, 94, 181-188. [ Links ]
Hobson, K. A., Schell, D. M., Renouf, D., & Noseworthy, E. (1996). Stable carbon and nitrogen fractionation between diet and tissues of captive seals: Implications for dietary reconstructions involving marine mammals. Canadian Journal of Fisheries and Aquatic Sciences, 53, 528-533. [ Links ]
Hobson, K. A., & Sease, J. L. (1998). Stable isotope analyses of tooth annuli reveal temporal dietary records: an example using Steller sea lions. Marine Mammal Science, 14, 116-129. [ Links ]
Hoyos-Padilla, E. M. (2009). Patrones de movimiento del tiburón blanco (Carcharodon carcharias) en isla Guadalupe, México (Tesis de doctorado, CICIMAR, IPN). [ Links ]
Hussey, N. E., Dudley, S. F. J., McCarthy, I. D., Geremy, C., & Fisk, A. T. (2011). Stable isotope profiles of large marine predators: viable indicators of trophic position, diet, and movement in sharks. Canadian Journal of Fisheries and Aquatic Sciences, 68, 2029-2045. [ Links ]
Itano, D. G. (2000). The reproductive biology of yellowfin tuna (Thunnus albacares) in Hawaiian waters and the western tropic Pacific Ocean: project summary. SOEST 00-01 JIMAR Contribution 00-328. Pelagic Fisheries Research Program, JIMAR, University of
Hawaii. [ Links ]
Jackson, A. L., Inger, R., Bearhop, S., & Parnell, A. (2009). Erroneous behaviour of MixSIR, a recently published Bayesian isotope mixing model: a discussion of Moore & Semmens (2008). Ecology Letters, 12, E1-E5. [ Links ]
Jorgensen, S. J., Reeb, C. A., Chapple, T. K., Anderson, S., Perle, C., Van Sommeran, S. R., Fritz-Cope, C., Brown, A. C., Klimley, A. P., & Block, B. A. (2009). Philopatry and migration of Pacific white sharks. Proceedings of the Royal Society B: Biological Sciences, 277, 679-688. [ Links ]
Jorgensen, S. J., Chapple, T. K., Anderson, S., Hoyos, M., Reeb, C., & Block, B. (2012). Connectivity among White Shark Coastal Aggregation Areas in Northeastern Pacific. In M. L. Domeier (Ed.), Global Perspectives on the Biology and Life History of the White Shark (pp. 159-167). Florida: CRC Press. [ Links ]
Kennedy, B. P., Blum, J. D., Folt, C. L., & Nislow, K. H. (1997). Natural isotope markers in salmon. Nature, 387, 766-767. [ Links ]
Kerr, L. A., Andrews, A. H., Cailliet, G. M., Brown, T. A., & Coale, K. H. (2006). Investigations of Δ14C, δ13C, and δ15N in vertebrae of white shark (Carcharodon carcharias) from the eastern North Pacific Ocean. Environmental Biology of Fishes,77, 337-353. [ Links ]
Kim, S. L., Tinker, T., Estes, J. A., & Koch, P. L. (2012a). Ontogenetic and Among-Individual Variation in Foraging Strategies of Northeast Pacific White Sharks Based on Stable Isotope Analysis. PLoS ONE, 7(9), e45068. [ Links ]
Kim, S. L., del Rio, C. M. N., Casper, D., & Koch, P. L. (2012b). Isotopic incorporation rates for shark tissues from a long-term captive feeding study. The Journal of Experimental Biology, 215, 2495-2500. [ Links ]
Lesage, V., Hammill, M. O., & Kovacs, K. M. (2001). Marine mammals and the community structure of the estuary and gulf of St Lawrence, Canada: evidence from stable isotope analysis. Marine Ecology Progress Series, 210, 203-221. [ Links ]
Long, J. D., & Jones, R. (1996). White Shark Predation and Scavening on Cetaceans in the Eastern North Pacific Ocean. In A. P. Klimley & D. G. Ainley (Eds.), Great white sharks: the biology of Carcharodon carcharias (pp. 161-177). San Diego: Academic Press. [ Links ]
Malpica-Cruz, L., Herzka, S. Z., Sosa-Nishizaki, O., Lazo, J. P., & Trudel, M. (2012). Tissue-specific isotope trophic discrimination factors and turnover rates in a marine elasmobranch: empirical and modeling results. Canadian Journal of Fisheries and Aquatic Sciences, 69, 551-564. [ Links ]
McClellan, C. M., Braun-McNeill, J., Avens, L., Wallace, B. P., & Read, A. J. (2010). Stable isotopes confirm a foraging dichotomy in juvenile loggerhead sea turtles. Journal of Experimental Marine Biology and Ecology, 387, 44-51. [ Links ]
Morales-Bojórquez, E., Hernández-Herrera, A., Nevárez-Martínez, M. O., & Díaz-Uribe, J. G. (2012). Population size of the jumbo squid Dosidicus gigas in the central Gulf of California, Mexico, based on markrecapture data. Aquatic Biology, 15, 27-34. [ Links ]
Morey, G., Martínez, M., Massutí, E., & Moranta, J. (2003). The Occurrence of White Sharks, Carcharodon carcharias, Around the Balearic Islands (Western Mediterranean Sea). Environmental Biology of Fishes, 68, 425-432. [ Links ]
Motta, Ρ. J. (1977). Anatomy and functional morphology of dermal collagen fibres in shark. Copeia, 3, 454-464. [ Links ]
Moore, J. W., & Semmens, B. X. (2008). Incorporating uncertainty and prior information into stable isotope mixing models. Ecology Letters, 11, 470-480. [ Links ]
Newsome, S. D., Koch, P. L., Etnier, M. A., & Aurioles-Gamboa, D. (2006). Using carbon and nitrogen isotope values to investigate maternal strategies in northeast Pacific otariids. Marine Mammal Science, 22, 556-572. [ Links ]
Okamoto, H., & Bayliff, W. H. (2003). A review of the Japanese longline fishery for tunas and billfishes in the eastern Pacific Ocean, 1993-1997. Inter-american tropical tuna commission bulletin, 22, 221-431. [ Links ]
Olson, R. J., Popp, B. N., Graham, B. S., Lopez-Ibarra, G. A., Galván-Magaña, F., Lennert-Cody, C. E., Bocanegra-Castillo, N., Wallsgrove, N. J., Gier, E., Alatorre-Ramirez, V., Balance, L. T., & Fry, B. (2010). Food-web inferences of stable isotope spatial patterns in copepods and yellowfin tuna in the pelagic eastern Pacific Ocean. Progress in Oceanography, 86, 124-138. [ Links ]
Parnell, A. C., Inger, R., Bearhop, S., & Jackson, A. L. (2010). Source partitioning using stable isotopes: coping with too much variation. PLoS ONE, 5, e9672. [ Links ]
Phillips, D. L., & Gregg, J. W. (2003). Source partitioning using stable isotopes: coping with too many sources. Oecologia, 136, 261-269. [ Links ]
Porras-Peters, H., Aurioles-Gamboa, D., Cruz-Escalona, V. H., & Koch, P. L. (2008). Trophic level and overlap of sea lions (Zalophus californianus) in the Gulf of California, Mexico. Marine Mammal Science, 24, 554-576. [ Links ]
Post, D., Layman, C., Arrington, D., Takimoto, G., Quattrochi, J., & Montaña, C. (2007). Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia, 152, 179-189. [ Links ]
Ruiz-Cooley, R. I., Gendron, D., Aguiñiga, S., Mesnick, S., & Carriquiry, J. D. (2004). Trophic relationships between sperm whales and jumbo squid using stable isotopes of C and N. Marine Ecology Progress Series, 277, 275-283. [ Links ]
Schell, D. M., Barnett, B. A., & Vinette, K. A. (1998). Carbon and nitrogen isotope ratios in zooplankton of the Bering, Chukchi and Beaufort seas. Marine Ecology Progress Series, 162, 11-23. [ Links ]
Smale, M. J., & Cliff, G. (2012). White sharks and Cephalopod Prey. Indicators of Habitat use? In M.L Domeier (Ed.), Global Perspectives on the Biology and Life History of the White Shark (pp. 51-57). Florida: CRC Press. [ Links ]
Tieszen, L. L., Boutton, T. W., Tesdahl, K. G., & Slade, N. A. (1983). Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet. Oecologia, 57, 32-37. [ Links ]
Tricas, T. C., & McCosker, J. E. (1984). Predatory behavior of the white shark, Carcharodon carcharias, with notes on its biology. Proceedings of California Academy Science, 43, 221-238. [ Links ]
Wcisel, M., Chivelle, W., & Gottfried, M. D. (2010). A potential predation attempt by a great white shark on an Indo-Pacific humpback dolphin. South African Journal of Wildlife Research 40, 184-187. [ Links ]
Weng, K., Boustany, A., Pyle, P., Anderson, S., Brown, A., & Block, B. (2007). Migration and habitat of white sharks (Carcharodon carcharias) in the eastern Pacific Ocean. Marine Biology 152, 877-894. [ Links ]
West, A. G., Goldsmith, G., Brooks, P., & Dawson, T. (2010). Discrepancies between isotope ratio infrared spectroscopy and isotope ratio mass spectrometry for the stable isotope analysis of plant and soil waters. Rapid Communications in Mass Spectrometry 24, 1948-1954. [ Links ]
Winn, H. E., Bishoff, W. L., & Tarusky, A. G. (1973). Cytological sexing of Cetacea. Marine Biology 23: 343-346. [ Links ]
*Correspondencia:
Mario Jaime-Rivera. Centro de Investigaciones Biológicas del Noroeste, Mar Bermejo No. 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, México; carcharodonmaritus@yahoo.com.mx
Javier Caraveo-Patiño. Centro de Investigaciones Biológicas del Noroeste, Mar Bermejo No. 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, México; jcaraveo04@cibnor.mx
Mauricio Hoyos-Padilla. Pelagios-Kakunjá, Sinaloa 1540, La Paz, B.S.C. México; amuakua@gmail.com
Felipe Galván-Magaña. Centro Interdisciplinario de Ciencias Marinas, Av. Instituto Politécnico Nacional s/n Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, Mexico; galvan.felipe@gmail.com
1. Centro de Investigaciones Biológicas del Noroeste, Mar Bermejo No. 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, México; carcharodonmaritus@yahoo.com.mx
2. Pelagios-Kakunjá, Sinaloa 1540, La Paz, B.S.C. México; amuakua@gmail.com
3. Centro Interdisciplinario de Ciencias Marinas, Av. Instituto Politécnico Nacional s/n Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, Mexico; galvan.felipe@gmail.com
Received 11-II-2013. Corrected 20-X-2013. Accepted 26-XI-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}