











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.62 no.2 San José abr./jun. 2014
Inter-habitat variation in density and size composition of reef fishes from the Cuban Northwestern shelf
Variación entre hábitats de la densidad y composición por tallas en peces de arrecife de la región noroccidental de Cuba
Variación entre hábitats de la densidad y composición por tallas en peces de arrecife de la región noroccidental de Cuba
*Dirección para correspondencia:
Abstract
Movement and exchange of individuals among habitats is critical for the dynamics and success of reef fish populations. Size segregation among habitats could be taken as evidence for habitat connectivity, and this would be a first step to formulate hypotheses about ontogenetic inter-habitat migrations. The primary goal of our research was to find evidence of inter-habitat differences in size distributions and density of reef fish species that can be classified a priori as habitat-shifters in an extensive (~600km2) Caribbean shelf area in NW Cuba. We sampled the fish assemblage of selected species using visual census (stationary and transect methods) in 20 stations (sites) located in mangrove roots, patch reefs, inner zone of the crest and fore reef (12-16m depth). In each site, we performed ten censuses for every habitat type in June and September 2009. A total of 11 507 individuals of 34 species were counted in a total of 400 censuses. We found significant differences in densities and size compositions among reef and mangrove habitats, supporting the species-specific use of coastal habitats. Adults were found in all habitats. Reef habitats, mainly patch reefs, seem to be most important for juvenile fish of most species. Mangroves were especially important for two species of snappers (Lutjanus apodus and L. griseus), providing habitat for juveniles. These species also displayed well defined gradients in length composition across the shelf. Rev. Biol. Trop. 62 (2): 589-602. Epub 2014 June 01.
Key words: reef fishes, Caribbean, habitat, size composition, Cuba, ontogenetic changes.
Resumen
El movimiento e intercambio de individuos entre hábitats son aspectos críticos para la dinámica y el éxito de las poblaciones de peces arrecifales. La segregación de tallas entre hábitats puede ser considerada como evidencia para la conectividad de hábitats y esta podría ser el primer paso para la formulación de hipótesis sobre migraciones ontogenéticas entre hábitats. El objetivo principal de nuestra investigación fue encontrar evidencia de diferencias entre hábitats en la distribución de tallas y densidad de especies de peces de arrecifes, las cuales pueden ser clasificadas, a priori, como especies con cambios ontogénicos en el hábitat, en una área extensa (~600 km2) de plataforma en el Caribe. Se realizaron censos visuales (métodos estacionario y de recorrido) de especies de peces seleccionadas en 20 sitios localizados en raíces de mangle, arrecifes de parche, la zona interna de la cresta arrecifal y el arrecife frontal (12-16m de profundidad). Se hicieron diez censos en cada sitio de los cuatro tipos de hábitat y se repitió este esquema en junio y septiembre 2009. Fueron contados un total de 11 507 individuos de 34 especies en un total de 400 censos. Se encontraron diferencias significativas en la densidad y composición por tallas entre los hábitats de arrecife y manglar. Estas diferencias, indicaron un uso variable de los hábitats costeros dependiendo de la especie. Los adultos fueron encontrados en todos los hábitats. Los hábitats de arrecifes, principalmente los arrecifes de parche, parecen ser muy importantes para los juveniles de peces de la mayoría de las especies en nuestra área de estudio. Los manglares fueron especialmente importantes para dos especies (L. apodus y L. griseus), que a lo largo de la plataforma fueron las únicas que mostraron gradientes bien definidos en las tallas.
Palabras clave: peces de arrecife, Caribe, hábitat, composición por tallas, Cuba, cambios ontogenéticos.
Movement and exchange of individuals among habitats is critical for the dynamics and success of reef fish populations. Many fish species move among different coastal habitats created by mangroves, seagrass beds, and coral reefs during their life cycles (Grol, Nagelkerken, Rypel & Layman, 2011). Consequently, conservation efforts to protect species and bio- diversity are developing to provide protection of habitats including linking corridors for all life stages (Mumby et al., 2004; Sale et al. ,2005). Recent research indicates that more consideration should be given to the ecological processes that occur along nursery-reef boundaries that connect neighboring ecosystems (Nagelkerken, Grol & Mumby, 2012).
In a recent review, Adams et al. (2006) classified coral reef fishes based on their ontogenic migration patterns. The authors define three groups: Group A) habitat specialists that use the same habitat at all life stages, Group B) habitat generalists which are not site-attached and use a variety of habitats, and Group C) ontogenetic shifters. The latter species switch habitats during their life time, such as the transition from larval to juvenile to maturing adults. These species in particular depend on ontogenetic habitat change from the back- reef to the fore-reef (Adams & Ebersole, 2009).
The nursery function of back-reef habitats has been inferred by studying spatial and temporal patterns in the size distribution of juveniles and adults across such habitats (Mumby et al., 2004; verweij, Nagelkerken, Wartenbergh, Pen & van der velde, 2006; Dorenbosch, Verbeck, Nagelkerken & Van der Velde, 2007). Size segregation among habitats could be taken as evidence for habitat connectivity and this would be a first step to formulate hypotheses about ontogenetic inter-habitat migrations.
Most research on reef fish habitat segregation in the Caribbean has been done at relative small spatial scales (<10km2). Several studies are restricted to non-estuarine bays of small islands (Nagelkerken & van der velde, 2002; Beets, Haught & Schmitges, 2003). In one case, the study area was a portion of the shelf less than 10km2 (Aguilar-Perera & Appeldoorn, 2007). In other cases, several locations were sampled around a small island with a total surface of less than 200km2 (Adams & Ebersole, 2002; Grober-Dunsmore, Frazer, Lindberg & Beets, 2007). Mumby et al. (2004) and Chittaro, Usseglio & Sale (2005) studied larger spatial scales (10-100km2).
Fish assemblages of the NW Cuban shelf have been recently investigated and spatial variations of assemblage composition have been discussed at a region-wide scale (González-Sansón et al., 2009a; González-Sansón, Aguilar, Hernández, Cabrera & Curry, 2009b), and at smaller scales (Aguilar, González-Sansón, Munkittrick & MacLatchy, 2004; Aguilar, González-Sansón, Faloh & Curry, 2008; González-Sansón, Aguilar, Hernández & Cabrera, 2009c; González-Sansón & Aguilar, 2010).
The goals of these studies were to identify natural and anthropogenic factors that may influence fish assemblage structure. All of these and other studies across the Caribbean (Nagelkerken & van der velde, 2002; Beets et al., 2003; Aguilar-Perera & Appeldoorn, 2007) suggest a degree of habitat connectivity, but none was designed to specifically test the hypothesis of ontogenetic, habitat migrations, which probably occur across larger spatial scales of >100km2.
The primary goal of our research was to find evidence for habitat-related differences in size composition of a group of selected fish species considered as habitat shifters at the NW Cuban shelf. The coast in this region has a variable shelf width but a general profile from mangrove, seagrass to coral reef is found, which makes it an ideal area to study ontogenetic habitat shifts of reef fishes. We hypothesized that ontogenetic migrations occur, and then tested the predictions that inter-habitat differences in size distribution and density of reef fish species would be apparent. The specific predictions were: (1) fish density and size composition for selected species among habitats have significant differences; and (2) a gradient in length composition across the island shelf from mangrove to fore reef is found.
Materials and methods
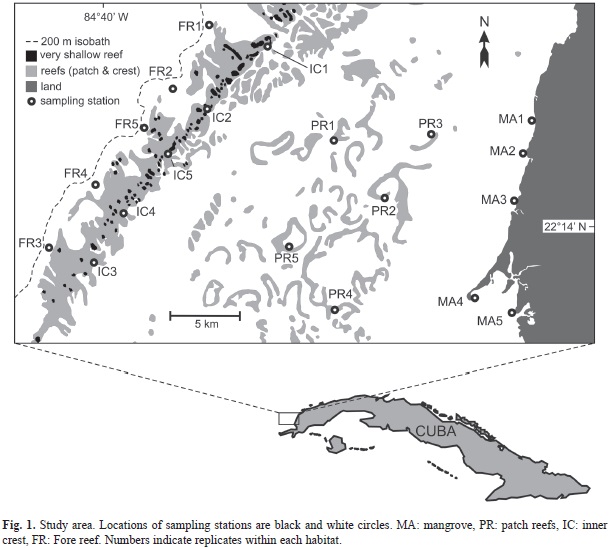
Study area: We included the spatial coverage assumed to be the normal range of reef fish species (e.g. Nemeth, Blondeau, Hezlieb & Kadison, 2007; Pina-Amargós et al., 2008), we studied an extensive area of the Caribbean shelf in Northwestern Cuba (Fig. 1). The shelf in the study area is broad with a distance from shore to the shelfedge (200m isobath) ranging from 22 to 36km. Total surface of the study area was 584km2. Mangroves were present along the shore occupying an almost continuous fringe of ~26km (excluding the coastal indentations). Red mangrove (Rhizophora mangle) was dominant and its roots provide a shallow (<1m deep) habitat for fishes. The width of the aquatic habitat associated with mangrove roots was narrow (<5m) due to the small tidal range (<0.5m) in the area (Claro, Lindeman& Parenti, 2002). There is a bank-barrier reef near the edge of the shelf. Corals occupied an almost continuous and shallow (≤5m deep) reef crest 21km long and 2-5km wide. Dominant coral species were Porites astreoides, Millepora complanata and Acropora palmata. The fore-reef was narrow ranging from 500-900m from the crest to the 50m isobath. Coral diversity was relatively high for a Caribbean reef and dominant species were Siderastrea spp., Porites astreoides, Montastrea spp. and Millepora alcicornis (González-Díaz, González-Sansón, Alvarez & Perera, 2010). There is a wide back-reef lagoon between the crest and the mangrove fringe. The depth of the lagoon varied from 6-8m near the crest and mangrove to 15-20m in the central portion. At this deeper zone, there were many patch reefs which rose from the bottom 14-18m and 6-9m from the surface. The remaining lagoons were covered by sand and extensive patches of turtle grass (Thalassia testudinum).
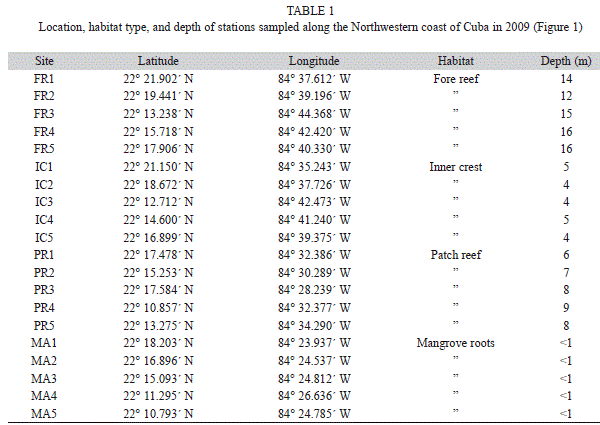
Sampling methods: We sampled four habitat types: mangrove (MA), patch reefs (PR), inner zone of the crest (IC) and fore-reef (FR). We located five sampling stations within each habitat type (Fig. 1, Table 1). Stationary visual censuses (Bohnsack & Bannerot, 1986) were conducted by two scuba divers at the PR, IC and FR habitats. Instead of 7m as recommended in the method, we used a radius of 5m (area of sampling unit ~79m2) to address any potential visibility variability among sites. At mangrove roots, we used 40m long transects (measured with a rope) for visual censuses by two snorkelers. The average distance of observation within the root habitat was 2m from the edge of the mangrove roots towards land, for a total area of 80m2, which was equivalent to that of the stationary visual censuses. Ten censuses (five per each diver) were done at each station for each habitat type. This sampling protocol was repeated twice over one week in June and September 2009.
For the census, we targeted a priori a selected group of reef fish species considered habitat-shifters (sensu Adams et al., 2006) by several authors (Cocheret de la Moriniere, Pollux, Nagelkerken & van der velde, 2002; Dorenbosch et al., 2007). These species were from the families: Serranidae (genera Cephalopholis, Epinephelus and Mycteroperca), Lutjanidae, Haemulidae, Chaetodontidae, Scaridae and Acanthuridae, and Lachnolaimus maximus (Labridae) and Sphyraena barracuda (Sphyraenidae). The individuals of Scarus iseri and S. taeniopterus were usually indistinguishable in the field and we hereafter refer to them as S. iseri/taeniopterus. Following the approach of Nagelkerken, van der velde, Gorissen, Meijer, van’t Hof & den Hartog (2000), we used 5cm bins to estimate fish body length to reduce differences in size estimation between observers. We used the criteria of Nagelkerken & van der velde (2002) to classify the sampled fishes as juvenile or adults.
For the inter-habitat comparisons in our study area, we did not transform the data to density in terms of number of individuals per unit area. Instead, we averaged the values of the ten censuses made at each station, and used this average as a density indicator expressed as the number of fish per 80m2 (Bohnsack & Bannerot, 1986). It is a relative estimate of abundance that allowed for comparisons within this study.
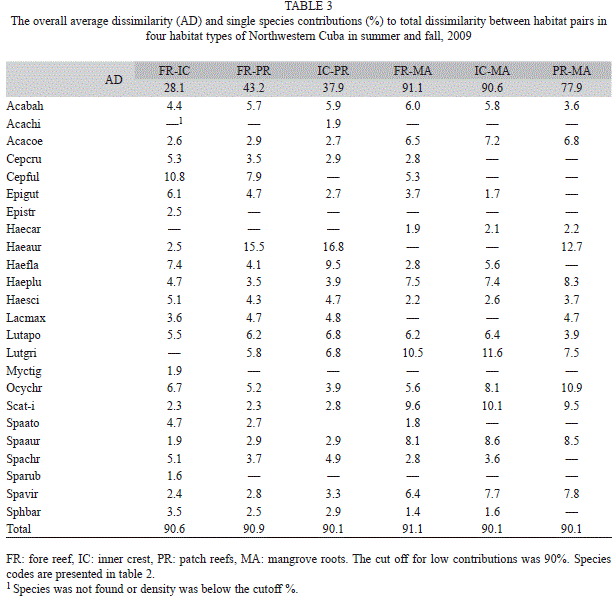
A hierarchical agglomerative cluster analysis was performed using the Bray-Curtis index as the similarity measure and clustering based on group average (UPGMA). SIMPROF test (Clarke, Somerfield & Gorley, 2008) was used for identifying significant clusters. Nonparametric multidimensional scaling (MDS) was employed for ordination of samples based in the same distance matrix as the cluster analysis. The combination of clustering and ordination analyses has been described by Clarke & Gorley (2006) as the most effective way to check the adequacy and mutual consistency of both representations. Contribution of each species to total dissimilarity between pairs of habitats was calculated using the SIMPER routine. Only species adding to at least 90% dissimilarity were retained for comparative analyses. A two-way crossed ANOSIM based on a priori classification of samples (habitats x date) using sites inside habitats as replicates was performed on Bray-Curtis dissimilarities calculated between pairs of species. All analyses were performed on log-transformed number of individuals per count using PRIMER 6.0 (Clarke & Gorley, 2006).
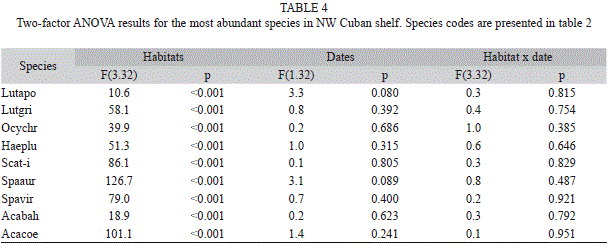
A two-way fixed effects factorial design ANOvA, using date of sampling and habitat as factors, was performed on the average number/80m2 (fourth-root transformed) for each of the most abundant species separately. A Student-Newman-Keuls post-hoc test was used for the pairwise comparisons of averages. Correlation between sampling dates for the percentage of juvenile fish was tested using the Spearman’s rank correlation coefficient. Significant correlation indicates that date of sampling had no effect on juvenile abundance. A Kruskal-Wallis test was used to test the significance of observed differences among habitats for the median juvenile percentage. A Post-hoc pairwise test using z-transformation was applied to discriminate differences between pairs of habitats. All tests were performed with a level of significance equal to 0.05 using STATISTICA 7.0 (Statsoft, 2006).
Results
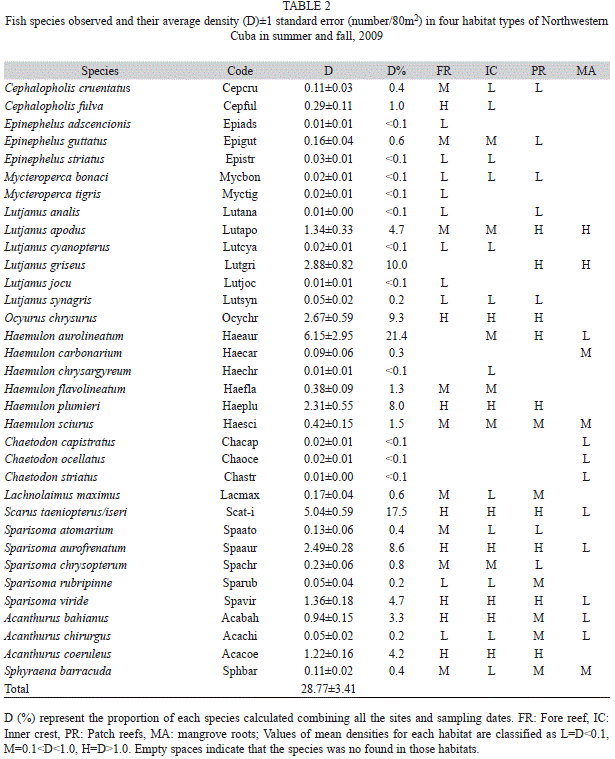
A total of 11 507 individuals of 34 species were counted in 400 censuses (Table 2). The ten most abundant species were Haemulon aurolineatum, S. iseri/taeniopterus, L. griseus, Ocyurus chrysurus, Sparisoma aurofrenatum, Haemulon plumieri, Sparisoma viride, L. apodus, Acanthurus coeruleus, and A. bahianus. These species accounted for >90% of individuals per census. Nine species occurred in at least two habitats (with averages >1/80m2). H. aurolineatum was present in large numbers at a few sites of the PR habitat resulting in a high aggregation (61.5±29.5). The remaining species had lower densities and thus we could not perform reliable single-species quantitative analyses. We used the total assemblage data for exploratory multivariate analyses and analyses by species restricted to the nine species present in at least two habitats.
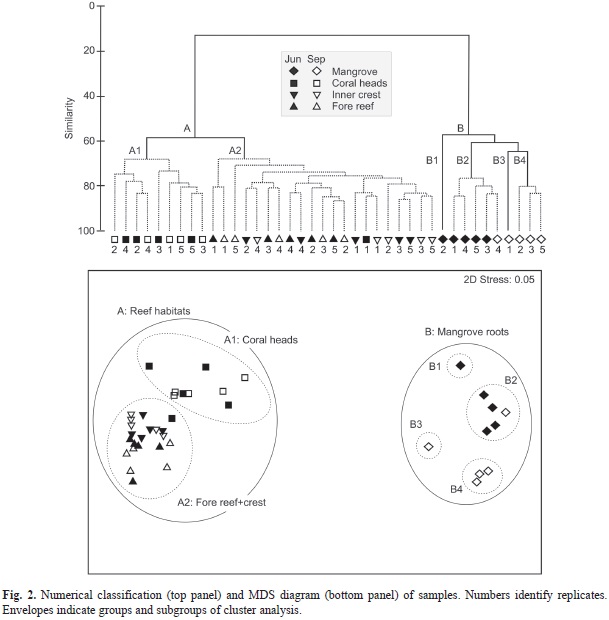
The assemblage composition varied notably among habitats (ANOSIM´s global R=0.752, p<0.001, 999 permutations), but was similar among sampling periods per habitat (global R=0.08, p=0.148, 999 permutations). All pairwise comparisons between habitats were also significant (p<0.005). The numerical classification and multidimensional scaling yielded diagrams those were highly consistent with ANOSIM results. Samples formed two well separated groups (SIMPROF Pi=16.36, p=0.001; Fig. 2). One group (A) included all reef samples and the other group (B) included all mangrove samples. Within the reef habitats, two subgroups were significantly separated (Pi=3.69, p=0.001): the A1 including patch reefs and the A2 fore-reef plus inner crest habitats. Mangrove stations showed a greater heterogeneity and split into four subgroups (B1-B2, B3, B4: Pi=2.83, p=0.03; B2-B3, B4: Pi=2.82, p=0.04 and B3-B4: Pi=3.96, p=0.024). Group B2 included mostly June, and group B4 included mostly September stations. Group B1 and group B3 included only one station in June and September, respectively.
The nine species used for two factorial analyses made an important contribution to the dissimilarity among habitats (Table 3). Other species were also important because they were abundant in one habitat (H. aurolineatum) or were observed in just one habitat (Cephalopholis fulva in FR). Some rare species also made smaller contributions (>4%) to total dissimilarities (e.g., Cephalopholis cruentata, Epinephelus guttatus, Haemulon flavolineatum, and Sparisoma chrysopterum) in reef habitats but not mangrove roots (<1/80m2). Haemulon sciurus was present in all habitats with a low average density (<1/80m2) but densities in patch reefs and mangroves were more than twice the inner crest and fore reef habitats.
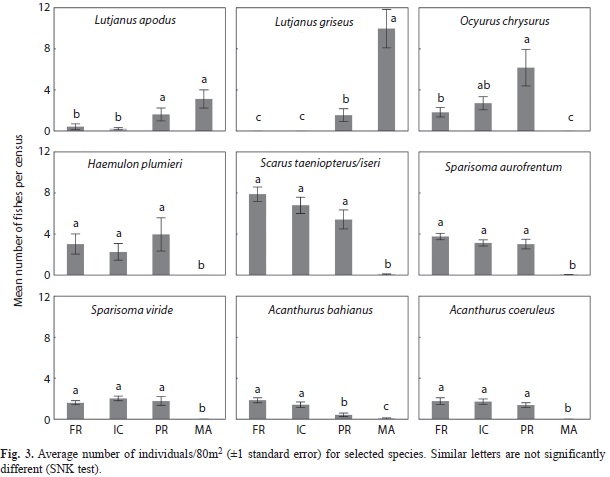
No significant differences in relative abundance were found between sampling periods for any species. Significant differences in fish densities were found between habitats in all cases (Table 4; Fig. 3). L. apodus showed significantly higher density at mangrove and patch reefs. Density of L. griseus in mangrove roots was the highest, followed by a lower value in patch reefs, but this species was not recorded in other habitats. O. chrysurus was not recorded in mangrove habitats and had higher densities in patch reefs with a progressive decrease towards the fore reef. H. plumieri, S. taeniopterus/iseri, S. aurofrenatum, S. viride and A. coeruleus showed no significant differences in density among reef habitats, and were mostly absent in mangrove roots. Density of A. bahianus was higher in fore reef and inner crest habitats, lower in patch reefs, and observed the lowest values in mangrove roots.
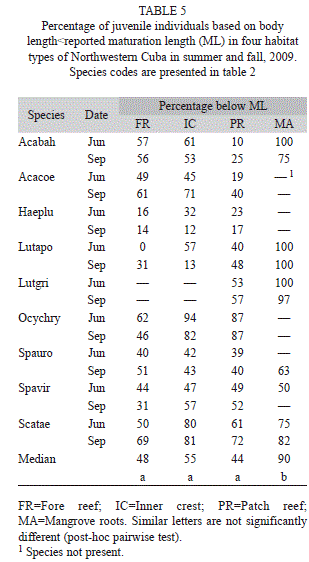
Juvenile proportions changed among habitats and species, but showed less variability between sampling dates (Table 5). Based in our results, we grouped juvenile species in three categories: a) species that were not present in mangrove roots; b) species present in all sampled habitats; and c) species that were more abundant in mangrove roots. For the three species that were not present in mangrove roots habitats, no clear trend in size composition was found. For A. coeruleus, the percentage of juvenile individuals was high (45-71%) in fore reef and inner crest habitats; while O. chrysurus was found in most habitats (62-94%). H. plumieri were mostly adults (68-88%) in all habitats.
Additionally, four species (S. taeniopterus/ iseri, S. viride, S. aurofrenatum and Acanthurus bahianus) were present in all habitats but were scarce in mangrove roots. No clear trend was evident in three species of parrotfishes (S. taeniopterus/iseri, S. viride and S. aurofrenatum) and most of these species were juveniles (40-82%). A. bahianus individuals were larger in patch reefs with a low representation of juvenile fish (10-25%). In the rest of the habitats, however, the percentages of juveniles for this species were greater (53-100%).
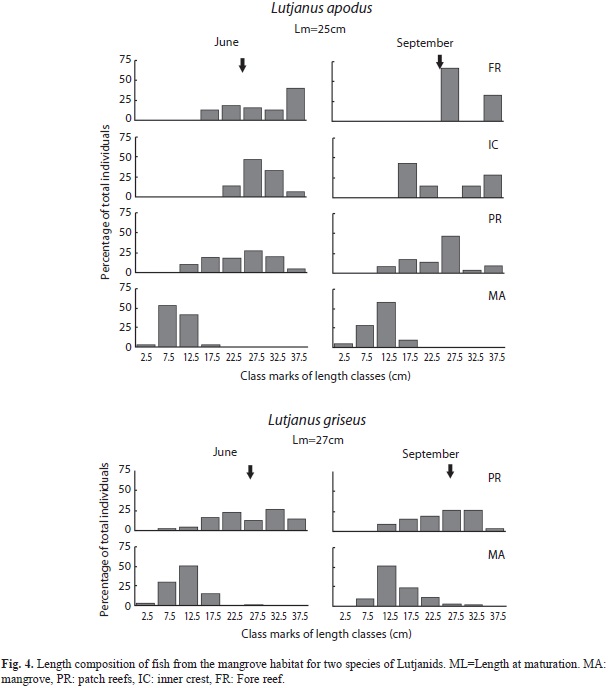
L. apodus and L. griseus were the only species that were abundant in mangrove root habitats as juveniles, yet they were larger in body size at reef habitats. For this reason, we presented a detailed analysis of size distributions by habitat for these two species (Fig. 4). Juveniles of L. apodus were present in all habitats and their proportion decreased from mangrove to fore reef habitats (mangrove, 100%; patch, 40-48%; inner crest, 13-57%; fore reef, 0-31%). Individuals of L. griseus were almost only juveniles in mangrove root habitats (97- 100%), while juvenile fish decreased notably in patch reefs (53-57%). This species was not observed in inner crest and fore reef stations.
The estimated percentage of juvenile fish per species per habitat showed a significant correlation between sampling dates (rs=0.787, p<0.001, n=30). Kruskal-Wallis test pooling Species all nine species data yielded a significant value indicating differences among habitats (H=18.04, p=0004, n=60). After a post-hoc pairwise comparison of the medians, mangrove habitat values were significantly higher compared to other habitats (not significantly different, Table 5).
Discussion
The presence of adult fish in all sampled habitats was consistent with Sheaves (2005) and Chittaro et al. (2005) conclusions that the term ‘nursery area’ applied for mangroves and seagrass habitats should be used with discretion. Based in our results, we consider the term ‘adult habitat’ applied for coral reefs should be used also with discretion. As Gillanders, Able, Brown, Eggleston & Sheridan (2003) discuss, it is important to measure the life history and specific movements of individuals, if we were to better understand the location and function of nursery habitats for marine organisms. Supporting former ideas, we found adults of scarid species in mangrove roots, and juvenile of all most abundant fish species in reef habitats.
Our data did not allow a quantitative approach to define nursery areas (sensu Beck et al., 2001) or essential juvenile habitats (Dahlgren et al., 2006; Layman et al, 2006). Instead, we discuss juvenile habitats as a qualitative labeling for those places where a relative high percent (>40%) of individuals were classified as juveniles. Generally, juveniles were relatively abundant in reef habitats and most abundant in mangrove roots. Nagelkerken et al. (2000) showed that fish had a preference for specific nursery habitats, but that most species used multiple nursery habitats simultaneously. The value of nursery habitats is species specific and a combination of habitats is used by high densities of juveniles (Aguilar-Perera, 2004).
Our results for A. bahianus and A. coeruleus corroborate earlier findings. Nagelkerken et al. (2000) found that juveniles of Acanthuridae were restricted to the shallow water biotopes (mangrove, notches in fossil reef rock, seagrass bed and fossil reef boulders), whereas adults were found on the reef. In contrast, Cocheret de la Moriniere et al. (2002) reported that A. bahianus was not observed in mangroves. Aguilar-Perera & Appeldorn (2007) found a similar trend for A. coeruleus and A. bahianus which were almost absent from mangroves in their study.
Habitat and size distribution patterns of Lutjanidae in our study agree with those reported by other authors. Nagelkerken et al. (2000) found that L. apodus and L. griseus were more abundant in mangrove habitats and that L. apodus showed an ontogenetic shift to deeper coral reefs. Cocheret de la Moriniere et al. (2002), Chittaro et al. (2005), and Aguilar-Perera & Appeldoorn (2007) found that L. apodus and L. griseus showed strong preferences for mangroves over seagrass beds at all size-classes.
Juvenile yellowtail snapper (O. chrysurus) prefer seagrass over mangrove roots and other habitats (Nagelkerken et al., 2000; Aguilar-Perera & Appeldoorn, 2007), but they have been recorded in both habitats (Cocheret de la Moriniere et al., 2002). Huijbers, Nagelkerken, Debrot & Jongejans (2013) found that the contribution of the coral reef as a nursery habitat for O. chrysurus was minimal. We found relatively high abundances of juveniles of this species in all reef habitats except mangrove roots.
Mumby et al. (2004) reported that juveniles of the white grunt, H. plumieri, were common in mangrove habitats. Appeldoorn, Recksiek, Hill, Pagan & Dennis (1997) presented length-frequency distributions that indicated an offshore ontogenetic migration. Along the Northwest coast of Cuba, we observed no white grunt in mangrove roots, and the length composition in reef habitats did not suggest an ontogenetic migration, because proportions of juveniles were similar in all reef habitats. Aguilar-Perera & Appeldoorn (2007) reported that mean densities of juveniles were significantly higher in seagrass relative to mangroves and coral reefs.
Our results for species in the family Scaridae indicated that these species were found at all stages in all reef habitats. Aguilar-Perera & Appeldoorn (2007) found significant differences among habitats for S. aurofrenatum and S. viride juveniles, but not for S. iseri juveniles. The preference of S. aurofrenatum and S. iseri for reef habitats has also been reported by others (Adams & Ebersole, 2002; Chittaro et al., 2005).
In summary, we found significant differences in densities and size compositions of fishes among reef and mangrove habitats of Northwestern Cuba. These suggest a species-specific, differential use of coastal habitats during ontogeny. Adults were found in all habitats. Reef habitats, mainly patch reefs, seem to be very important for juvenile fish of most species in our study area. These results differ notably from the main trend found in the Caribbean, where studies report that most or all nursery fish species studied herein had the highest juvenile densities in mangrove areas only (Nagelkerken, 2009). In our study area, mangroves were especially important for the two snapper species (L. apodus and L. griseus) which were also the only species that showed well defined gradients in length composition from mangrove to the fore reef.
Acknowledgments
A part of this research was made possible with funding from The Bay and Paul Foundations and The Marine ventures Foundation. Special thanks to D. Guggenheim and F. Bretos for their support and friendship. We thank José Miguel Díaz Escrich, Commodore of the Hemingway International Yacht Club of Cuba, for his support during the organization of research trips. We thank also I. Rodriguez and L. V. Garcia as well as V. de la Guardia, skipper of the boat “MarinaBella” for their assistance in the field.
References
Adams, A. J., & Ebersole, J. P. (2002). Use of back-reef and lagoon habitats by coral reef fishes. Marine Ecology Progress Series, 228, 213-226. [ Links ]
Adams, A. J. & Ebersole, J .P. (2009). Mechanisms Affecting Recruitment Patterns of Fish and Decapods in Tropical Coastal Ecosystems. In I. Nagelkerken (Ed.), Ecological Connectivity among Tropical Coastal Ecosystems (pp.185-228). Netherlands: Springer. [ Links ]
Adams, A. J., Dahlgren, C. P., Kellison, G. T., Kendall, M. S., Layman, C. A., Ley, J. A., Nagelkerken, I., & Serafy, J. E. (2006). Nursery function of tropical back-reef systems. Marine Ecology Progress Series, 318, 287-301. [ Links ]
Aguilar, C., González-Sansón, G., Munkittrick, K. R., & MacLatchy, D. L. (2004). Fish assemblages on fringe coral reefs of the northern coast of Cuba near Havana Harbor. Ecotoxicology and Environmental Safety, 58, 126-138. [ Links ]
Aguilar, C., González-Sansón, G., Faloh, I., & Curry, R. A. (2008). Spatial variation in stable isotopes (δ13C and δ15N) in marine fishes along the coast of Havana City: Evidence of human impacts from harbor and river waters. Journal of Coastal Research, 24(5), 1281-1288. [ Links ]
Aguilar-Perera, J. A. (2004). Coastal habitat connectivity of reef fishes from southwestern Puerto Rico. (Doctoral dissertation). Retrieved from http://www. aoml.noaa.gov/general/lib/CREWS/Cleo/PuertoRi-co/prpdfs/aguilar-coastal.pdf. [ Links ]
Aguilar-Perera, J. A. & Appeldoorn, R. S. (2007). Variation in juvenile fish density along the mangrove–seagrass–coral reef continuum in SW Puerto Rico. Marine Ecology Progress Series, 348,139-148. [ Links ]
Appeldoorn, R. S., Recksiek, C. W., Hill, R. L., Pagan, F. E., & Dennis, G. D. (1997). Marine protected areas and reef fish movements: the role of habitat in con- trolling ontogenetic migration. Proceedings of the 8th International Coral Reef Symposium, 2, 1917-1922. [ Links ]
Beck, M. W., Heck Jr., K. L., Able, K. W., Childers, D. L., Eggleston, D. B., Gillanders, B. M., Halpern, B., Hays, C. G., Hoshino, K., Minello, T. J., Orth, R. J., Sheridan, P. F., & Weinstein, M. P. (2001). The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. BioScience, 51(8), 633-641. [ Links ]
Beets, J., Muehistein, L., Haught, K., & Schmitges, H. (2003). Habitat connectivity in coastal environments: patterns and movements of Caribbean coral reef fishes with emphasis on bluestriped grunt, Haemulon sciurus. Gulf and Caribbean Research, 14(2), 29-42. [ Links ]
Bohnsack, J. A. & Bannerot, S. P. (1986). A stationary visual census technique for quantitatively assessing community structure of coral reef fishes. NOAA Technical Report NMFS (41). Miami, FL.: National Oceanic and Atmospheric Administration, Southeast Science Fisheries Center. [ Links ]
Chittaro, P. M., Usseglio, P., & Sale, P. F. (2005). Variation in fish density, assemblage composition and relative rates of predation among mangroves, seagrass and coral reef habitats. Environmental Biology of Fishes, 72(2), 175-187. [ Links ]
Clarke, K. R. & Gorley, R. N. (2006). PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth. [ Links ]
Clarke, K. R., Somerfield, P. J., & Gorley, R. N. (2008). Testing of null hypotheses in exploratory community analyses: similarity profiles and biota-environment linkage. Journal of Experimental Marine Biology and Ecology, 366, 56-69. [ Links ]
Claro, R., Lindeman, K. C., & Parenti, L. R.(2002). Ecology of the marine fishes of Cuba. Washington: Smithsonian Institution Scholarly Press. [ Links ]
Cocheret de la Morinière, E., Pollux, B. J. A., Nagelkerken, I., & Van der Velde, G. (2002). Post-settlement life cycle migration patterns and habitat preference of coral reef fish that use seagrass and mangrove habitats as nurseries. Estuarine, Coastal and Shelf Science, 55, 309-321. [ Links ]
Dahlgren, C. P., Kellison, G. T., Adams, A. J., Gillanders, B. M., Kendall, M. S., Layman, C. A., Ley, J. A., Nagelkerken, I., & Serafy, J. E. (2006). Marine nurseries and effective juvenile habitats: concepts and applications. Marine Ecology Progress Series 312, 291-295. [ Links ]
Dorenbosch, M., Verberk, W. C. E. P., Nagelkerken, I., & Van der Velde, G. (2007). Influence of habitat configuration on connectivity between fish assemblages of Caribbean seagrass beds, mangroves and coral reefs. Marine Ecology Progress Series, 334, 103-116. [ Links ]
Gillanders, B. W., Able, K. W., Brown, J. A., Eggleston, D. B., & Sheridan, P. F. (2003). Evidence of connectivity between juvenile and adult habitats for mobile marine fauna: an important component of nurseries. Marine Ecology Progress Series, 247, 281-295. [ Links ]
González-Díaz, P., González-Sansón, G., Alvarez, S., & Perera, O. (2010). High spatial variability of coral, sponges and gorgonian assemblages in a well preserved reef. Revista de Biología Tropical, 58(2), 621-634. [ Links ]
González-Sansón, G. & Aguilar, C. (2010). Reef fish diversity components as indicators of cumulative effects in a highly impacted fringe reef. Ecological Indicators, 10(3), 766-772. [ Links ]
González-Sansón, G., Aguilar, C., Hernández, I., Cabrera, Y., Suarez-Montes, N, Bretos, F., & Guggenheim, D. (2009a). Natural and human-induced variability in the composition of fish assemblages in the Northwestern Cuban shelf. Revista de Biología Tropical, 57(3), 721-740. [ Links ]
González-Sansón, G., Aguilar, C., Hernández, I., Cabrera, Y., & Curry, A. (2009b). The influence of habitat and fishing on reef fish assemblages in Cuba. Gulf and Caribbean Research, 21, 13-21. [ Links ]
González-Sansón, G., Aguilar, C., Hernández, I., & Cabrera, Y. (2009c). Effects of depth and bottom communities on the distribution of highly territorial reef fish in the Northwestern region of Cuba. Journal of Applied Ichthyology, 25(6), 652-660. [ Links ]
Grober-Dunsmore, R., Frazer, T. K., Lindberg, W. J., & Beets, J. (2007). Reef fish and habitat relationships in a Caribbean seascape: the importance of reef context. Coral Reefs, 26(1), 201-216. [ Links ]
Grol, M. G. G., Nagelkerken, I., Rypel, A. L., & Layman, C. A. (2011). Simple ecological trade-offs give rise to emergent cross-ecosystem distributions of a coral reef fish. Oecologia, 165, 79-88. [ Links ]
Huijbers, C. M., Nagelkerken, I., Debrot, A. O., & Jongejans, E. (2013) Geographic coupling of juvenile and adult habitat shapes spatial population dynamics of a coral reef fish. Ecology, 94, 1859-1870. [ Links ]
Layman, C. A., Dahlgren, C. P., Kellison, G. T., Adams, A. J., Gillanders, B. M., Kendall, M. S., Ley, J. A., Nagelkerken, I., & Serafy, J. E. (2006). Marine nurseries and effective juvenile habitats. Marine Ecology Progress Series, 318, 307-308. [ Links ]
Mumby, P. J., Edwards, A. J., Arias-González, J. E., Lindeman, K. C., Blackwell, P. G. Gall, A., Gorczynska, M. I., Harborne, A. R., Pescod, C. L., Renken, H., Wabnitz, C. C. C., & Llewellyn, G. (2004). Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature, 427, 533-536. [ Links ]
Nagelkerken, I. (2009). Evaluation of nursery function of mangroves and seagrass beds for tropical decapods and reef fishes: patterns and underlying mechanisms. In I. Nagelkerken (Ed.), Ecological connectivity among tropical coastal ecosystems (p. 357-399). Dordrecht: Springer. [ Links ]
Nagelkerken, I., Grol, M. G. G., & Mumby, P. J. (2012). Effects of Marine Reserves versus Nursery Habitat Availability on Structure of Reef Fish Communities. PLoS ONE 7(6), e36906. [ Links ]
Nagelkerken, I., van der Velde, G., Gorissen, M. W., Meijer, G. J., Van’t Hof, T., & den Hartog, C. (2000). Importance of mangroves, seagrass beds and the shallow coral reef as a nursery for important coral reef fishes, using a visual census technique. Estuarine, Coastal and Shelf Science, 51(1), 31-44. [ Links ]
Nagelkerken, I. & van der Velde, G. (2002). Do non-estuarine mangroves harbor higher densities of juvenile fish than adjacent shallow-water and coral reef habitats in Curacao (Netherlands Antilles)? Marine Ecology Progress Series, 245, 191-204. [ Links ]
Nemeth, R. S., Blondeau, J., Herzlieb, S., & Kadison, E. (2007). Spatial and temporal patterns of movement and migration at spawning aggregations of red hind, Epinephelus guttatus, in the U.S. Virgin Islands. Environmental Biology of Fishes, 78, 365-381. [ Links ]
Pina-Amargós, F., González-Sansón, G., Jiménez del Castillo, A., Zayas, A., Martín, F., & Acosta de la Red, W. (2008). Movement of adult fish in the Jardines de la Reina Marine Reserve, Cuba. In R. Grober-Dunsmore & B. D. Keller (Eds.), Caribbean connectivity: Implications for marine protected area management (pp.121-138). Proceedings of a Special Symposium, 9-11 November 2006, 59th Annual Meeting of the Gulf and Caribbean Fisheries Institute, Belize City, Belize. Marine Sanctuaries Conservation Series ONMS-08-07. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, Office of National Marine Sanctuaries, Silver Spring, MD. [ Links ]
Sale, P. F., Cowen, R. K., Danilowicz, B. S., Jones, G. P., Kritzer, J. P., Lindeman, K. C., Planes, S., Polunin, N. V. C., Russ, G. R., Sadovy, Y. J., & Steneck, R. S. (2005). Critical science gaps impede use of no-take fishery reserves. Trends in Ecology & Evolution, 20 (2), 74-80. [ Links ]
Sheaves, M. (2005). Nature and consequences of biological connectivity in mangrove systems. Marine Ecology Progress Series, 302, 293-305. [ Links ]
StatSoft (2006). STATISTICA (data analysis software system) version 7.1. www.statsoft.com. [ Links ]
Verweij, M. C., Nagelkerken, I., Wartenbergh, S. L. J., Pen, I. R., & van der Velde, G. (2006). Caribbean mangroves and seagrass beds as daytime feeding habitats for juvenile French grunts, Haemulon flavolineatum. Marine Biology, 149, 1291-1299. [ Links ]
Adams, A. J. & Ebersole, J .P. (2009). Mechanisms Affecting Recruitment Patterns of Fish and Decapods in Tropical Coastal Ecosystems. In I. Nagelkerken (Ed.), Ecological Connectivity among Tropical Coastal Ecosystems (pp.185-228). Netherlands: Springer. [ Links ]
Adams, A. J., Dahlgren, C. P., Kellison, G. T., Kendall, M. S., Layman, C. A., Ley, J. A., Nagelkerken, I., & Serafy, J. E. (2006). Nursery function of tropical back-reef systems. Marine Ecology Progress Series, 318, 287-301. [ Links ]
Aguilar, C., González-Sansón, G., Munkittrick, K. R., & MacLatchy, D. L. (2004). Fish assemblages on fringe coral reefs of the northern coast of Cuba near Havana Harbor. Ecotoxicology and Environmental Safety, 58, 126-138. [ Links ]
Aguilar, C., González-Sansón, G., Faloh, I., & Curry, R. A. (2008). Spatial variation in stable isotopes (δ13C and δ15N) in marine fishes along the coast of Havana City: Evidence of human impacts from harbor and river waters. Journal of Coastal Research, 24(5), 1281-1288. [ Links ]
Aguilar-Perera, J. A. (2004). Coastal habitat connectivity of reef fishes from southwestern Puerto Rico. (Doctoral dissertation). Retrieved from http://www. aoml.noaa.gov/general/lib/CREWS/Cleo/PuertoRi-co/prpdfs/aguilar-coastal.pdf. [ Links ]
Aguilar-Perera, J. A. & Appeldoorn, R. S. (2007). Variation in juvenile fish density along the mangrove–seagrass–coral reef continuum in SW Puerto Rico. Marine Ecology Progress Series, 348,139-148. [ Links ]
Appeldoorn, R. S., Recksiek, C. W., Hill, R. L., Pagan, F. E., & Dennis, G. D. (1997). Marine protected areas and reef fish movements: the role of habitat in con- trolling ontogenetic migration. Proceedings of the 8th International Coral Reef Symposium, 2, 1917-1922. [ Links ]
Beck, M. W., Heck Jr., K. L., Able, K. W., Childers, D. L., Eggleston, D. B., Gillanders, B. M., Halpern, B., Hays, C. G., Hoshino, K., Minello, T. J., Orth, R. J., Sheridan, P. F., & Weinstein, M. P. (2001). The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. BioScience, 51(8), 633-641. [ Links ]
Beets, J., Muehistein, L., Haught, K., & Schmitges, H. (2003). Habitat connectivity in coastal environments: patterns and movements of Caribbean coral reef fishes with emphasis on bluestriped grunt, Haemulon sciurus. Gulf and Caribbean Research, 14(2), 29-42. [ Links ]
Bohnsack, J. A. & Bannerot, S. P. (1986). A stationary visual census technique for quantitatively assessing community structure of coral reef fishes. NOAA Technical Report NMFS (41). Miami, FL.: National Oceanic and Atmospheric Administration, Southeast Science Fisheries Center. [ Links ]
Chittaro, P. M., Usseglio, P., & Sale, P. F. (2005). Variation in fish density, assemblage composition and relative rates of predation among mangroves, seagrass and coral reef habitats. Environmental Biology of Fishes, 72(2), 175-187. [ Links ]
Clarke, K. R. & Gorley, R. N. (2006). PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth. [ Links ]
Clarke, K. R., Somerfield, P. J., & Gorley, R. N. (2008). Testing of null hypotheses in exploratory community analyses: similarity profiles and biota-environment linkage. Journal of Experimental Marine Biology and Ecology, 366, 56-69. [ Links ]
Claro, R., Lindeman, K. C., & Parenti, L. R.(2002). Ecology of the marine fishes of Cuba. Washington: Smithsonian Institution Scholarly Press. [ Links ]
Cocheret de la Morinière, E., Pollux, B. J. A., Nagelkerken, I., & Van der Velde, G. (2002). Post-settlement life cycle migration patterns and habitat preference of coral reef fish that use seagrass and mangrove habitats as nurseries. Estuarine, Coastal and Shelf Science, 55, 309-321. [ Links ]
Dahlgren, C. P., Kellison, G. T., Adams, A. J., Gillanders, B. M., Kendall, M. S., Layman, C. A., Ley, J. A., Nagelkerken, I., & Serafy, J. E. (2006). Marine nurseries and effective juvenile habitats: concepts and applications. Marine Ecology Progress Series 312, 291-295. [ Links ]
Dorenbosch, M., Verberk, W. C. E. P., Nagelkerken, I., & Van der Velde, G. (2007). Influence of habitat configuration on connectivity between fish assemblages of Caribbean seagrass beds, mangroves and coral reefs. Marine Ecology Progress Series, 334, 103-116. [ Links ]
Gillanders, B. W., Able, K. W., Brown, J. A., Eggleston, D. B., & Sheridan, P. F. (2003). Evidence of connectivity between juvenile and adult habitats for mobile marine fauna: an important component of nurseries. Marine Ecology Progress Series, 247, 281-295. [ Links ]
González-Díaz, P., González-Sansón, G., Alvarez, S., & Perera, O. (2010). High spatial variability of coral, sponges and gorgonian assemblages in a well preserved reef. Revista de Biología Tropical, 58(2), 621-634. [ Links ]
González-Sansón, G. & Aguilar, C. (2010). Reef fish diversity components as indicators of cumulative effects in a highly impacted fringe reef. Ecological Indicators, 10(3), 766-772. [ Links ]
González-Sansón, G., Aguilar, C., Hernández, I., Cabrera, Y., Suarez-Montes, N, Bretos, F., & Guggenheim, D. (2009a). Natural and human-induced variability in the composition of fish assemblages in the Northwestern Cuban shelf. Revista de Biología Tropical, 57(3), 721-740. [ Links ]
González-Sansón, G., Aguilar, C., Hernández, I., Cabrera, Y., & Curry, A. (2009b). The influence of habitat and fishing on reef fish assemblages in Cuba. Gulf and Caribbean Research, 21, 13-21. [ Links ]
González-Sansón, G., Aguilar, C., Hernández, I., & Cabrera, Y. (2009c). Effects of depth and bottom communities on the distribution of highly territorial reef fish in the Northwestern region of Cuba. Journal of Applied Ichthyology, 25(6), 652-660. [ Links ]
Grober-Dunsmore, R., Frazer, T. K., Lindberg, W. J., & Beets, J. (2007). Reef fish and habitat relationships in a Caribbean seascape: the importance of reef context. Coral Reefs, 26(1), 201-216. [ Links ]
Grol, M. G. G., Nagelkerken, I., Rypel, A. L., & Layman, C. A. (2011). Simple ecological trade-offs give rise to emergent cross-ecosystem distributions of a coral reef fish. Oecologia, 165, 79-88. [ Links ]
Huijbers, C. M., Nagelkerken, I., Debrot, A. O., & Jongejans, E. (2013) Geographic coupling of juvenile and adult habitat shapes spatial population dynamics of a coral reef fish. Ecology, 94, 1859-1870. [ Links ]
Layman, C. A., Dahlgren, C. P., Kellison, G. T., Adams, A. J., Gillanders, B. M., Kendall, M. S., Ley, J. A., Nagelkerken, I., & Serafy, J. E. (2006). Marine nurseries and effective juvenile habitats. Marine Ecology Progress Series, 318, 307-308. [ Links ]
Mumby, P. J., Edwards, A. J., Arias-González, J. E., Lindeman, K. C., Blackwell, P. G. Gall, A., Gorczynska, M. I., Harborne, A. R., Pescod, C. L., Renken, H., Wabnitz, C. C. C., & Llewellyn, G. (2004). Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature, 427, 533-536. [ Links ]
Nagelkerken, I. (2009). Evaluation of nursery function of mangroves and seagrass beds for tropical decapods and reef fishes: patterns and underlying mechanisms. In I. Nagelkerken (Ed.), Ecological connectivity among tropical coastal ecosystems (p. 357-399). Dordrecht: Springer. [ Links ]
Nagelkerken, I., Grol, M. G. G., & Mumby, P. J. (2012). Effects of Marine Reserves versus Nursery Habitat Availability on Structure of Reef Fish Communities. PLoS ONE 7(6), e36906. [ Links ]
Nagelkerken, I., van der Velde, G., Gorissen, M. W., Meijer, G. J., Van’t Hof, T., & den Hartog, C. (2000). Importance of mangroves, seagrass beds and the shallow coral reef as a nursery for important coral reef fishes, using a visual census technique. Estuarine, Coastal and Shelf Science, 51(1), 31-44. [ Links ]
Nagelkerken, I. & van der Velde, G. (2002). Do non-estuarine mangroves harbor higher densities of juvenile fish than adjacent shallow-water and coral reef habitats in Curacao (Netherlands Antilles)? Marine Ecology Progress Series, 245, 191-204. [ Links ]
Nemeth, R. S., Blondeau, J., Herzlieb, S., & Kadison, E. (2007). Spatial and temporal patterns of movement and migration at spawning aggregations of red hind, Epinephelus guttatus, in the U.S. Virgin Islands. Environmental Biology of Fishes, 78, 365-381. [ Links ]
Pina-Amargós, F., González-Sansón, G., Jiménez del Castillo, A., Zayas, A., Martín, F., & Acosta de la Red, W. (2008). Movement of adult fish in the Jardines de la Reina Marine Reserve, Cuba. In R. Grober-Dunsmore & B. D. Keller (Eds.), Caribbean connectivity: Implications for marine protected area management (pp.121-138). Proceedings of a Special Symposium, 9-11 November 2006, 59th Annual Meeting of the Gulf and Caribbean Fisheries Institute, Belize City, Belize. Marine Sanctuaries Conservation Series ONMS-08-07. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, Office of National Marine Sanctuaries, Silver Spring, MD. [ Links ]
Sale, P. F., Cowen, R. K., Danilowicz, B. S., Jones, G. P., Kritzer, J. P., Lindeman, K. C., Planes, S., Polunin, N. V. C., Russ, G. R., Sadovy, Y. J., & Steneck, R. S. (2005). Critical science gaps impede use of no-take fishery reserves. Trends in Ecology & Evolution, 20 (2), 74-80. [ Links ]
Sheaves, M. (2005). Nature and consequences of biological connectivity in mangrove systems. Marine Ecology Progress Series, 302, 293-305. [ Links ]
StatSoft (2006). STATISTICA (data analysis software system) version 7.1. www.statsoft.com. [ Links ]
Verweij, M. C., Nagelkerken, I., Wartenbergh, S. L. J., Pen, I. R., & van der Velde, G. (2006). Caribbean mangroves and seagrass beds as daytime feeding habitats for juvenile French grunts, Haemulon flavolineatum. Marine Biology, 149, 1291-1299. [ Links ]
*Correspondencia a:
Consuelo Aguilar: Centro de Investigaciones Marinas, Universidad de la Habana, Cuba. Primera No.114, Playa, La Habana. Departamento de Estudios para el Desarrollo Sustentable de Zonas Costeras, Universidad de Guadalajara, Gómez Farías 82, C.P. 48980, San Patricio-Melaque, Cihuatlán, Jalisco, México; coquiaguilar06@yahoo.es. Correspondence
Gaspar González-Sansón: Departamento de Estudios para el Desarrollo Sustentable de Zonas Costeras, Universidad de Guadalajara, México; gaspargonzalez2001@yahoo.es
Yureidy Cabrera:Centro de Investigaciones Marinas, Universidad de la Habana, Cuba. Actualmente en Dirección de Regulaciones Pesqueras y Ciencia, Ministerio de la Industria Alimentaria; yurecab06@yahoo.com
Alexei Ruiz: Centro de Investigaciones Marinas, Universidad de la Habana, Cuba; alexeis@cim.uh.cu
R. Allen Curry: Canadian Rivers Institute, University of New Brunswick, PO Box 4400, Fredericton, NB. E3B 5A3, Canada; racurry@unb.ca
1.Centro de Investigaciones Marinas, Universidad de la Habana, Cuba. Primera No.114, Playa, La Habana. Departamento de Estudios para el Desarrollo Sustentable de Zonas Costeras, Universidad de Guadalajara, Gómez Farías 82, C.P. 48980, San Patricio-Melaque, Cihuatlán, Jalisco, México; coquiaguilar06@yahoo.es. Correspondence
2.Departamento de Estudios para el Desarrollo Sustentable de Zonas Costeras, Universidad de Guadalajara, México; gaspargonzalez2001@yahoo.es
3.Centro de Investigaciones Marinas, Universidad de la Habana, Cuba. Actualmente en Dirección de Regulaciones Pesqueras y Ciencia, Ministerio de la Industria Alimentaria; yurecab06@yahoo.com
4. Centro de Investigaciones Marinas, Universidad de la Habana, Cuba; alexeis@cim.uh.cu
5.Canadian Rivers Institute, University of New Brunswick, PO Box 4400, Fredericton, NB. E3B 5A3, Canada; racurry@unb.ca
Received 22-VII-2013. Corrected 18-X-2013. Accepted 20-XI-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}