











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.62 no.2 San José abr./jun. 2014
Effect of clonal reproduction on genetic structure in Pentaclethra macroloba (Fabaceae: Mimosoideae)
Efecto de la reproducción clonal en la estructura genética de Pentaclethra macroloba (Fabaceae: Mimosoideae)
Efecto de la reproducción clonal en la estructura genética de Pentaclethra macroloba (Fabaceae: Mimosoideae)
*Dirección para correspondencia:
Abstract
The existence of monodominant forests on well-drained soils in tropical regions has been widely reported. Such forests most likely result from a combination of both ecological and evolutionary factors. Under conditions of high seed and seedling mortality, vegetative reproduction could create a reproductive advantage leading to forest dominance, and profoundly affect the distribution of genetic variation in a clonal species. We investigated these effects in a low diversity forest site in Northeastern Costa Rica dominated by the species Pentaclethra macroloba, which sprouts from the root mass of fallen trees and from snapped trunks. We examined the population structure of juvenile P. macroloba growing in different soil types and across an elevational gradient. Using seven molecular markers, we genotyped 173 juvenile P. macroloba from 18 plots (six plots in seasonally inundated swamps, and 12 plots in upland non-swamp) spanning 50-300m in elevation at La Selva Biological Station and the adjacent Reserva Ecológica Bijagual in Northeastern Costa Rica. We answered two specific questions: (1) How extensive is clonal reproduction? and (2) what is the distribution of genetic diversity and structure?. We found that clonal reproduction occurred exclusively within inundated swamp areas. However, there was no significant difference between genetic diversity measures in swamp and non-swamp plots, which were both generally low when compared with other tropical forest species. Genetic structure was significant across all plots (FST=0.109). However, genetic structure among swamp plots (FST=0.128) was higher than among non-swamp upland plots (FST=0.093). Additionally, spatial autocorrelation among individuals within non-swamp upland plots was significant from the 25 to 100m spatial scale, but not within swamp plots. The degree of overall genetic structure we found in P. macroloba is high for a tropical forest tree. The incidence of clonal reproduction is a contributing factor in genetic differentiation, but the high structure among plots without clonal reproduction indicates that other factors contribute as well.
Key words: Costa Rica, genetic diversity, La Selva Biological Station, Reserva Ecológica Bijagual, sprouting, tropical forest.
Resumen
La existencia de bosques monodominantes sobre suelos bien drenados en regiones tropicales ha sido ampliamente reportada. Investigaciones recientes han sugerido que tales bosques son probablemente resultado de una combinación de factores ecológicos y evolutivos. Bajo condiciones de alta mortalidad de semillas y plántulas, la reproducción vegetativa podría crear una ventaja reproductiva llevando a la dominancia del bosque, pero también podría afectar profundamente la distribución de la variación genética en especies clonales. Investigamos estos efectos en un sitio de bosque con baja diversidad de especies en el Noreste de Costa Rica que es ampliamente dominado por la especie Pentaclethra macroloba, la cual retoña de la masa de raíces de árboles caídos y de troncos partidos. Examinamos la estructura poblacional de individuos juveniles de P. macroloba creciendo en diferentes tipos de suelo y a través de un gradiente de altitud. Utilizamos siete marcadores moleculares, genotipamos 173 Pentaclethra macroloba de 18 parcelas (seis en ciénagas y 12 en ambientes no cenagosos) ubicados en un gradiente de elevación entre 50-300m en las reservas adyacentes: Reserva Biológica Bijagual y Estación Biológica La Selva, en el centro de Costa Rica. Abordamos dos preguntas específicas: (1) ¿Qué tan extensa es la reproducción clonal? y (2) ¿Cuál es la distribución de diversidad y estructura genética? Encontramos que la reproducción clonal ocurrió exclusivamente dentro de áreas cenagosas inundadas. La estructura genética fue significativa en todas las parcelas (FST=0.109). Observamos una estructura genética más alta entre poblaciones juveniles dentro de las ciénagas (FST=0.128) comparada con poblaciones no cenagosas en parcelas a mayor altura (FST=0.093), con mayor autocorrelación espacial en sitios no cenagosos en el intervalo entre 25 y 100m. La presencia de reproducción clonal no afectó significativamente las medidas de diversidad entre las dos áreas, que fueron generalmente bajas comparadas con otras especies de bosque tropical. El alto grado de estructura genética en general es novedoso para un árbol de bosque tropical. La incidencia de reproducción clonal es un factor que contribuye en la diferenciación genética, pero la alta estructura en parcelas sin reproducción clonal indica que otros factores están contribuyendo también.
Palabras clave: Costa Rica; diversidad genética; Estación Biológica La Selva; Reserva Ecológica Bijagual; germinación; bosque tropical.
Our understanding of the ecology of tropical forests has been shaped by the paradigm of high species diversity and low density of individual tree species (Wright, 2002; Terborgh, 2012). However, the existence of monodominant tropical forests has increasingly drawn the attention of tropical forest biologists. Such forests are known to occur throughout the world’s tropical regions (Richards, 1952; Whitmore & Burnham, 1975; Connell & Lowman, 1989; Hart, 1990, ter Steege et al., 2013). Torti, Coley and Kursar (2001) reviewed the phenomenon, and concluded that monodominance in tropical forests is not the result of a single mechanism, but rather, of multiple factors operating both within and across sites. Peh, Lewis and Lloyd (2011) classified the various proposed mechanisms for tropical monodominance into those operating at the ecological and the evolutionary levels, and developed a probabilistic model to explain how monodominance can result from a combination of influences and life history characteristics.
In this study, we investigated a possible mechanism for forest dominance by one of the best-studied tree species at one of the most well-studied tropical forest sites, La Selva Biological Station in Costa Rica. At La Selva, the forest is dominated by the tree species Pentaclethra macroloba (Willd.) Benth (Fabaceae: Mimosoideae). In the Atlantic lowlands of Northern Costa Rica, P. macroloba is a frequent canopy dominant in seasonally inundated swamps as well as drier uplands (Hartshorn, 1975; Hartshorn & Hammel, 1994). Although P. macroloba at La Selva comprises a smaller percent of the forest than the 40-60% expected for a forest dominant (Peh et al., 2011), it has been cited as exemplary of monodominance (Connell & Lowman, 1989). By any measure, P. macroloba is extremely important and abundant at La Selva, and is an important driver of forest processes. In previous survey, P. macroloba comprised nearly 40% of the aboveground biomass across multiple study plots at varying elevations within La Selva (Clark & Clark, 2000), and accounted for up to 13% of all stems, 13.9% of total leaf area index, and 35.3% of total basal area (Clark, Olivas, Oberbauer, Clark, & Ryan, 2008). The species has been the focus of multiple ecological studies focusing on physiology (e.g. Oberbauer & Strain 1985, 1986; Oberbauer, Strain, & Riechers, 1987); demography (Hartshorn, 1972, 1975); genetics (Hall, Chase, & Bawa, 1994); relationships with other organisms [seed predators (Guariguata, Adame, & Finegan, 2000; Hart-shorn, 1972; McKenna & McKenna, 2006); mycorrhizae (Janos, 1980); floral visitors (Perry, 1983)]; carbon dynamics (Russell, Raich, Arrieta, Valverde-Barrantes, & González, 2010); and role in forest structure (Clark & Clark, 2000; Hartshorn, 1975; Lieberman, Lieberman, Peralta, & Hartshorn, 1996). In spite of all this work, a basic question remains unanswered: why is the La Selva forest dominated by P. macroloba, and how does this species maintain its numerical dominance?
One clue to the uncommonly high frequency of P. macroloba may lie in unique aspects of its life history. Demographic studies by Harts-horn (1972, 1975), indicated that in areas of inundated soil (such as lower elevations within the La Selva forest), vegetative sprouting from fallen trees is common. Fallen trees may provide sprouts with a mature root system and a ready source of mobilizable nutrients stored in woody tissue or in rapid local decomposition of the trunk, thus shifting carbon allocation to leaf production and stem growth (Loehle, 1987). For trees with high mortality of juveniles (such as is the case for P. macroloba at La Selva (Clark & Clark, 2006)), an asexual reproductive strategy may allow faster growth to reach a higher light environment than would sexual reproduction. If clonal reproduction is the dominant reproductive mode in wetter areas, we would expect lowered genetic diversity within lowland sites and enhanced genetic differentiation among populations.
One published study has addressed the population genetics of P. macroloba (Hall et al., 1994). Using three allozyme markers, Hall et al. (1994) reported that in both adults and juvenile trees sampled at and near La Selva, P. macroloba had low genetic diversity and little genetic differentiation among sites. To our knowledge, there have been no recent studies of the genetics of P. macroloba using more sensitive genetic markers. The reported low level of genetic diversity of P. macroloba could be a result of the limitations of allozymes as genetic markers. Alternatively, low genetic diversity may result from the unique ecology of this species. Given the ambiguity of these results, we reexamined the genetic pattern of juvenile P. macroloba at La Selva and at the nearby Reserva Ecológica Bijagual. We address two specific questions: (1) what is the extent of clonal reproduction in P. macroloba in lowland swamp habitat of La Selva Biological Station and at higher elevations; and (2) in light of the clonal ability of P. macroloba, what is the genetic structure and distribution of genetic diversity in juvenile trees? If these populations have a high degree of clonal reproduction, we would expect low genetic diversity within plots and high genetic structure among plots.
Materials and Methods
Study species: Pentaclethra macroloba (Willd.) Kuntze is a tropical canopy tree occurring from Central to South America. The species is a member of the Mimosoideae subfamily of Fabaceae, with racemic flowers and dehiscent pods. It is an outcrossing species that is putatively pollinated by small insects, including bees and beetles (Perry, 1983). Few flowers actually develop into seed pods, thus where it occurs it is possible for asexual reproduction to dominate (Flores, 2004). Cloning occurs in this species through vegetative sprouting off of fallen trees in moist areas (Hartshorn, 1972). Seeds are dispersed short distances by explosive dehiscence, while rodents (Beard, 1946; Hartshorn, 1972; Smythe, 1970) and water (Hartshorn, 1972; Williamson & Costa, 2000) move seeds longer distances.
Study area: Our study was conducted at two adjacent reserves in Northeastern Costa Rica. La Selva Biological Station, of the Organization for Tropical Studies, is one of the largest and most important tropical biological research stations in the world. Encompassing a total of 1 536ha, the reserve is nestled between two major rivers, the Sarapiquí and Puerto Viejo. The neighboring site, Reserva Biológica Bijagual, is 290ha and located upstream from La Selva at the confluence of the Tirimbina and Bijagual rivers (tributaries of the Sarapiquí). This area receives a mean annual rain fall of 4 367mm. Alluvial floods and recent volcanic activity provide these reserves with varying types of nutrient rich soil, allowing for over 2 000 vascular flora species to flourish (McDade & Hartshorn, 1994). Elevation ranges from 50 to over 400m.
Sampling: In 2011, we sampled leaf tissue from 173 juvenile P. macroloba from 18plots within La Selva and Bijagual reserves. Circular plots were 10m in diameter and distributed evenly at 50, 150, and 300m elevation (Table 1). A particular plot was used if it had at least eight individual P. macroloba between 0.5 and 2m. These plots were spaced a minimum of 50m apart and were located within two habitat types. All plots at 50m elevation were located in semi-inundated swamp habitat (hereafter, swamp). The 150 and 300m elevation plots were located in drier upland primary forest habitat (hereafter, non -swamp) . Leaf tissue was dried and preserved in silica gel until DNA extraction in laboratory. Voucher specimens of Pentaclethra macroloba are deposited in the herbarium at La Selva (Herbarium Index LSCR); collection numbers 7850 (R. Aguilar), 697 (O. Vargas), 2110 (O. Vargas), and 1777 (P. Foster).
Marker development: For the entire project, we used a modified Cetyltrimethylammonium Bromide (CTAB) protocol compiled from Doyle & Doyle (1987) and Cullings (1992) to extract DNA from leaf tissue. We added a prewash step derived from Li, Yang, Chen, Zhang & Tang, (2007) to remove secondary compounds common in leaf tissue such as polysaccharides, tannins, and polyphenolics that are known to inhibit PCR reactions.
We used high-throughput sequencing to generate a microsatellite library on a Roche 454 Genome Sequencer FLX instrument. We used 500ng of DNA to prepare our library according to the GS FLX+ Series – XL+ protocol (Roche Applied Sciences, Indianapolis, USA). Library fragment size distribution, quality, and quantity were assessed on an Agilent 2100 Bioanalyzer. We then processed our library through the GS FLX emPCR protocol for small and then large volumes. Sequencing was performed on a GS FLX+ picotiterplate and yielded 106 855 sequences.
Potential microsatellite loci were identified and screened. Of the 106 855 sequences scanned, we found multiple motifs with usable flanking primers using MsatCOMMANDER (Faircloth, 2008). This included 297 tetramer repeats, 1 650 trimer repeats, and 1 832 dimer repeats. We tested 48 markers (18 di, 18 tri, and 12 tetra) for PCR suitability and variation in 14 individuals distributed across the study range. All forward primers included an M13F(-20) sequence (GTAAAACGACGGCCAG) on the 5’ end to attach a fluorescent label and allow genotyping by size (Boutin-Ganache, Raposo, Raymond, & Deschepper, 2001). Fragment analysis was done at the UCLA Genotyping and Sequencing Core and genotype calls were done in the laboratory using GeneMapper® (Applied Biosystems). We eliminated monomorphic markers. We used Microchecker (Van Oosterhout, Hutchinson, Wills, & Shipley, 2004) to check for effect of stutter, allelic drop-out, and null alleles within the larger dataset (including unique genotypes and one representative from each clone set (n=162)). To ensure independence of loci, we tested for linkage disequilibrium using GenePop (Raymond & Rousset, 1995).
Genetic analysis: To identify clones, we matched multilocus genotypes using GenAlEx (Peakall & Smouse, 2006). Only individuals with identical genotypes that occurred within the same plot were considered clones in the final analysis. Identical genotypes across different plots were not considered clones because it was unlikely that they represented true clones given the distance between plots. This method provided a conservative estimate of clonal reproduction in our study area. We compare the proportion of clonal individuals between swamp and non-swamp areas using a non-parametric Wilcox test.
For all subsequent analyses, we examined the data in three ways: including all individuals (n=173), excluding clones (n=154), and including all unique genotypes and one representative from each clone set (n=162), referred to as “genets” Examining genotypes of all individuals identifies the genetic structure and distribution of genetic diversity in juvenile trees. Comparing these three sample sets identifies the effect of cloning on these patterns.
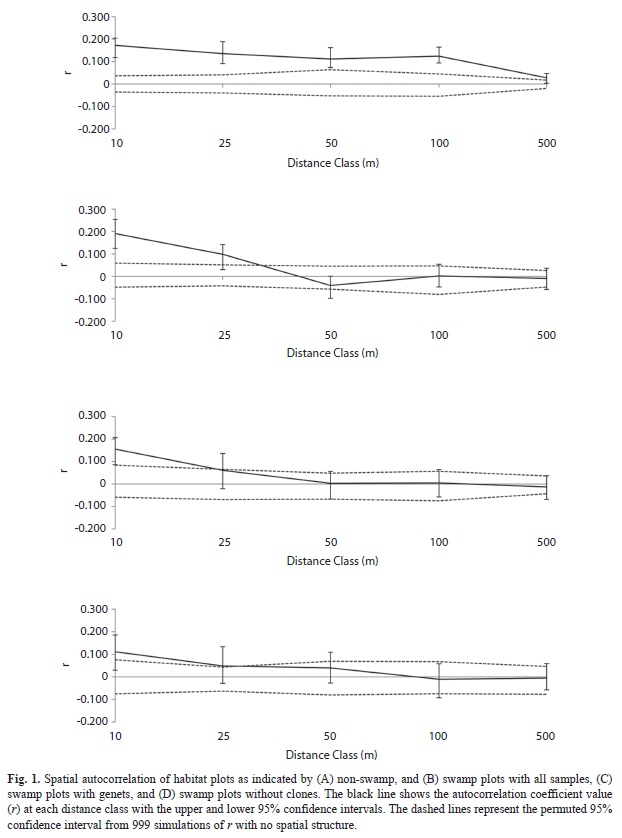
We identified the distribution of genetic diversity and structure using three methods implemented in GenAlEx (Peakall & Smouse, 2006). First, we measured genetic diversity at each site as the average number of observed alleles per locus (Na), average number of effective alleles per locus (Ne), observed heterozygous-ity (HO), and expected heterozygous-ity (HE). To identify a relationship between genetic diversity and elevation we ran a linear regression for each of these measures. Additionally, we ran t-tests to identify a difference in each genetic diversity measure between the two habitat types. Second, we examined the degree of genetic structure among plots using an AMOVA analysis reporting FST, FIS, and FIT values. From this analysis we were able to determine the degree of genetic separation between plots and estimate the average number of migrants between groups within a generation (Nm). Third, we used spatial autocorrelation to test for fine-scale genetic structure. For each pair of individuals, a correlation coefficient was calculated (r), which provides a measure of genetic similarity. The r values were grouped into five distance classes based on the distance between the individuals compared: 0-10m, 10-25m, 25-50m, 50-100m, and 100-500m. GenAlEx provides two measures of significant departure from the null hypothesis that r=0. The first method uses bootstrapping to create confidence intervals around the calculated r values for each distance class by sub-sampling from within the pairwise comparisons. If this bootstrapped confidence interval does not include a zero r-value, then there is a significant deviation from zero within the distance class. The second stronger measure uses a permutation method to create a distribution based on the assumption of no spatial structure, and the associated confidence interval. If the actual r value for the group is outside of this confidence interval, then there is a significant deviation from zero within the distance class.
Results
Of the 48 loci tested in 14 individuals, six were indiscernible or did not amplify, 35 were monomorphic, and seven were found to be variable for Pentaclethra macroloba (Table 2). None of our seven variable markers showed evidence for stuttering or allele dropout. Two markers, PMA11 and PMA38, showed significant evidence for possible null alleles. The size, shape and behavior of allelic peaks in these loci were distinct and consistent across identified clones, assuring us that the effect of null alleles on identifying genetic identity was negligible. After Bonferroni correction, we found no indication for linkage disequilibrium.
Clonal reproduction occurred only in swamp areas. Of those samples collected in swamps, 33% (19 individuals) had identical genotypes with at least one other individual within the same plot. Some genotypes occurred three times within a plot. Only one swamp plot had no duplicate genotypes (Table 1). The percent clonal reproduction in swamp plots was significantly greater than in non-swamp plots (Wilcox, p<0.001). We only observed one instance of identical genotypes between plots. These plots, 150-2 and 150-4, lay 100m apart. Thus, it is not clear if they represent true clones.
Overall, P. macroloba showed no clear pattern explaining the level of genetic diversity at a plot regardless of sample set. We found no significant relationship between the four diversity measures and elevation (linear regression, p>0.05) or difference between habitat types (Table 1, t-test, p>0.05). Average allelic diversity was relatively low, with most loci having only two to three observed alleles across the range (Table 1).
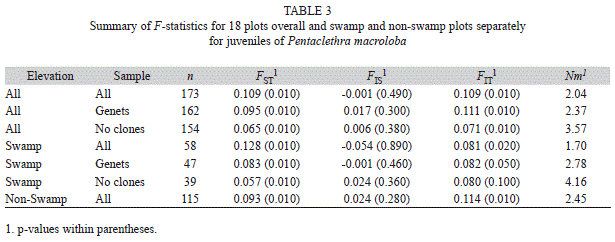
We found significant genetic structure among plots (Table 3). When we examined genetic structure separately for swamp and non-swamp plots using FST, swamp areas had greater genetic differentiation between plots when including all individuals, but was roughly equivalent or lower when looking at genets or excluding clones respectively (Table 3). Inbreeding values in swamp and non-swamp area did not differ significantly.
Fine-scale genetic structure differed between swamp and upland plots (Fig. 1). There was significant autocorrelation for the swamp plots at the two lowest distance classes: 10 and 25m, when examining all individuals (Fig. 1b), and only at 10m when examining genets or excluding all clones (Fig 1c & Fig1d). In contrast, individuals in the non-swamp regions showed significant autocorrelation at four distance classes: 10, 25, 50 and 100m (Fig. 1a). At the 50 and 100m distance class, the bootstrapped confidence intervals for r between swamp and non-swamp plots are non-overlapping, indicating a significantly greater spatial autocorrelation for non-swamps at that range.
Discussion
Our data provide strong evidence that P. macroloba reproduces clonally in swamps and not at higher elevations, as proposed by Hartshorn (1972) (Wilcox, p<0.001). Within the swamp, 33% of juveniles sampled were identified as clones. Previous work on other tropical trees with clonal sprouting ability has shown a similar total proportion of vegetative sprouts to seedlings (Putz & Brokaw, 1989) as well as similar habitat influence on sprouting ability (Lasso, Engelbrecht, & Dalling, 2009; Salk & McMahon, 2011). In contrast, of the 115 juveniles surveyed at higher elevations, no identical genotypes were found within the same plot. Thus, it appears that cloning is restricted to seasonally inundated habitats.
The incidence of asexual reproduction in P. macroloba strongly affects the distribution of genetic variation in swamps. Both swamp and higher forest areas exhibited significant genetic structure. Swamp plots showed a higher degree of differentiation when all individuals were included. This difference disappears when looking only at genets or without clones, indicating that larger structure values in swamp plots are driven by repeated genotypes, and have less to do with historic movement patterns or influence of cloning on adult populations. In contrast, when looking at the fine-scale genetic pattern in the analysis of spatial autocorrelation, we see that the extent and spatial scale of auto-correlation is greater for the juveniles in swamp plots than in the non-swamp plots regardless of including or excluding clones. The difference in autocorrelation and FST results is likely due to spatial scale of analysis. We see a difference in fine scale spatial structure between swamp and non-swamp sites from 25 to 100m, but not at greater distances. The FST values are calculated between plots, and the average interplot distance is 5km, explaining why we see equivalent FST values when comparing genets in the two habitats. If we assume that seed dispersal and selective forces are similar in swamp and non-swamp plots, then the difference in fine scale genetic structure between these two zones is likely due to higher clonal reproduction in swamps leading to higher genetic similarity locally, and a smaller proportion of dispersed individuals making up the adults and juveniles. These results illustrate the spatial consequences of vegetative reproduction compared to simple seed dispersal for genetic structure. However, this effect did not translate to a significant difference in the amount of genetic diversity in the two habitats. No genetic diversity measure showed a significant difference between the two habitats, or trend along elevation. Thus, although clonal reproduction may result in greater genetic dissimilarity between plots, it has not yet led to a decreased amount of genetic diversity in this area.
Our findings for P. macroloba contrast with those of other Costa Rican forest trees examined within La Selva. For example, Hanson Brunsfeld, Finegan and Waits (2008) found lower genetic structure among plots in the canopy tree species, Dipteryx panamensis (Fabaceae) (GST=0.050), than we found in P. macroloba (FST=0.109). Schierenbeck Brunsfeld, Finegan and Waits (1997) shows a similar disparity for the trees Dendropanax arboreus (Araliaceae) (ΦPT=0.032), Inga thibaudiana (Fabaceae: Mimosoideae) (ΦPT=0.009), and Protium glabrum (Burseraceae) (ΦPT=-0.003). The values we observed are not unheard of for tropical forest trees (Arias et al., 2012; Jones, Cerón-Souza, Hardesty, & Dick, 2013), but it is counterintuitive that P. macroloba, an abundant canopy dominant, would have such high genetic structure unless one considers its facultative asexual reproduction as a factor (Loveless & Hamrick, 1984). Nonetheless, the observation of high structure among upland non-clonal plots cannot be explained by our study.
Findings reported here are inconsistent with a previous population genetic study of P. macroloba diversity at La Selva. Hall et al. (1994) examined population differentiation of seedling and adults within La Selva and across much of Costa Rica. Using three allozy-me markers, they found lower heterozygos-ity (Ho=0.04) compared to ours (Ho=0.49) and a non-significant population differentiation in La Selva of seedlings (1.9%) and adults (3.8%). In contrast, we observed 10.9% differentiation among plots. We attribute most of this disparity to the difference between marker type and number. Microsatellite markers are more likely to detect genetic variation than allozymes, especially with only a few markers. Nonetheless, we found some evidence for low genetic diversity in P. macroloba. Of the 42 operable loci that we screened for microsatellite development, only 14% showed evidence of polymorphism. A quick survey of the most recent published articles using this same methodology for microsatellite development in plants shows on average 50% of loci tested were polymorphic, with a range of 38-72% (Fatemi, Haddadchi, & Gross, 2012; Nakahama et al., 2012; Sawicki et al., 2012; Sherman, Stanley, Keough, Gardner, & Macreadie, 2012). Our low number of polymorphic alleles could be due to differences in geographic range of these studies as well as life history characteristics of the plants involved. However, within the loci that we developed, we also see a low level of average allelic diversity (Na=2.47, Ne=1.98), which also indicates lower overall diversity of P. macroloba. While the Hall et al. (1994) findings underestimated genetic structure, their evidence of low genetic diversity is supported by our findings.
The potential for vegetative reproduction in Pentaclethra macroloba does not singularly explain the spatial genetic pattern we observed or this species’ dominance at La Selva. First, asexual reproduction was not dominant, with only 33% of juveniles in swamps and none outside having been produced clonally. Second, significant structure values were seen between juvenile plots in the non-swamp uplands where we saw no incidence of clonal reproduction. Third, there was no significant difference in inbreeding or genetic diversity values between the two habitats or across elevations. Low FIS values, as we observed, are generally expected in a facultatively asexual species with outcrossing due to fixation of heterozygotes (Navascues, Stoeckel, & Mariette, 2010; Stoeckel et al., 2006). If asexual contribution to the adult populations was significant, we would expect to see a reduced amount of variation centered in the swamp, where rare variant alleles would be lost through an overabundance of identical genotypes. This low diversity in the swamps is not what we observed.
In conclusion, our findings indicate that clonal reproduction in P. macroloba has increased genetic differentiation in populations at La Selva. However, sexual reproduction is still the dominant means of reproduction, even in swamp environments. Therefore, some other factors, such as low seed dispersal or demographic history, might account for these paradoxical patterns of genetic diversity in P. macroloba at La Selva.
Acknowledgments
This study was funded by NSF OISE-0854524 to HEB; by funding administered by Occidental College (Howard Hughes Medical Institute to HZ; ASP to HZ and ID), and by the Occidental College Biology Department (Hayes Fund). We thank the staffs of the Organization for Tropical Studies La Selva Biological Station (especially O. Vargas) and the Reserva Biológica Bijagual for intellectual and logistical support. Conversations with D.B. Clark, P. Foster, G.L. Hartshorn, J. Hamrick, G. B. North, and C. Prager were particularly helpful in generating the framework and sampling design. We thank J. Pollinger and B. Faircloth for guidance during molecular marker development, S. Nigenda Morales and A. Garmendia for abstract translation, and P. Thompson, S. Steele, and two anonymous reviewers whose comments greatly improved the manuscript.
References
Arias, D. M., Albarrán-Lara, A. L., González-Rodríguez, A., Peñaloza-Ramírez, J., Dorado, O., & Leyva, E. (2012). Genetic diversity and structure of wild populations of the tropical dry forest tree Jacaratia mexicana (Brassicales: Caricaceae) at a local scale in Mexico. Revista de Biología Tropical, 60(1), 1-10. [ Links ]
Beard, J. S. (1946). The Mora Forests of Trinidad, British WestIndies. Journal of Ecology, 33(2), 173-192. [ Links ]
Boutin-Ganache, I., Raposo, M., Raymond, M., & Deschepper, C. F. (2001). M13-tailed primers improve the readability and usability of microsatellite analyses performed with two different allele-sizing methods. Biotechniques, 31(1), 24-28. [ Links ]
Clark, D. B., & Clark, D. A. (2000). Landscape-scale variation in forest structure and biomass in a tropical rain forest. Forest Ecology and Management, 137(1-3), 185-198. [ Links ]
Clark, D. B, Olivas, P. C., Oberbauer, S. F., Clark, D. A, & Ryan, M. G. (2008). First direct landscape-scale measurement of tropical rain forest Leaf Area Index, a key driver of global primary productivity. Ecology Letters, 11(2), 163-172. [ Links ]
Clark, D. B., & Clark, D. A. (2006). Tree growth, mortaity, physical condition, and microsite in an old-growth lowland tropical rain forest. Ecology, 87(8), 2132. [ Links ]
Connell, J., & Lowman, M. (1989). Low-diversity tropical rain forests: some possible mechanisms for their existence. The American Naturalist, 134(1), 88-119. [ Links ]
Cullings, K. W. (1992). Design and testing of a plant-specific PCR primer for ecological and evolutionary studies. Molecular Ecology, 1(4), 233-240. [ Links ]
Doyle, J. J., & Doyle, J. L. (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin, 19, 11-15. [ Links ]
Faircloth, B. C. (2008). MSATCOMMANDER: detection of microsatellite repeat arrays and automated, locus-specific primer design. Molecular Ecology Resources, 8(1), 92-94. [ Links ]
Fatemi, M., Haddadchi, A., & Gross, C. L. (2012). The discovery of microsatellite markers for Hardenbergia violacea (Fabaceae), using next-generation sequencing. Conservation Genetics Resources, 4(4), 1063-1065. [ Links ]
Flores, E. M. (2004). Pentaclethra macroloba. Academia Nacional de Ciencies de Costa Rica. [ Links ]
Guariguata, M. R., Adame, J. J. R., & Finegan, B. (2000). Seed removal and fate in two selectively logged lowland forests with constrasting protection levels. Conservation Biology, 14(4), 1046-1054. [ Links ]
Hall, P., Chase, M. R., & Bawa, K. S. (1994). Low genetic-variation but high population differentiation in a common tropical forest tree species. Conservation Biology, 8(2), 471-482. [ Links ]
Hanson, T. R., Brunsfeld, S. J., Finegan, B., & Waits, L. P. (2008). Pollen dispersal and genetic structure of the tropical tree Dipteryx panamensis in a fragmented Costa Rican landscape. Molecular Ecology, 17(8), 2060-2073. [ Links ]
Hart, T. B. (1990). Monospecific dominance in tropical rain forests. Trends in Ecology & Evolution, 5(1), 6-11. [ Links ]
Hartshorn, G. S. (1972). The ecological life history and population dynamics of Pentaclethra macroloba, a wet forest dominant, and Stryphnodendron excelsum, and occasional associate. University of Washington. [ Links ]
Hartshorn, G. S. (1975). A matrix model of tree population dynamics. In F. B. Golley & E. Medina (Eds.), Tropical ecological systems (pp. 41-51). New York: Springer-Verlag. [ Links ]
Hartshorn, G. S., & Hammel, B. E. (1994). Vegetation types and floristic patterns. In La Selva: Ecology and natural history of a neotropical rain forest (pp. 73-89). Chicago, Illinois: University of Chicago Press. [ Links ]
Janos, D. (1980). Vesicular-arbuscular mycorrhizae affect lowland tropical rain forest plant growth. Ecology, 61(1), 151-162. [ Links ]
Jones, F. A., Cerón-Souza, I., Hardesty, B. D., & Dick, C. W. (2013). Genetic evidence of Quaternary demographic changes in four rain forest tree species sampled across the Isthmus of Panama. Journal of Biogeography, 40(4), 720-731. doi:10.1111/jbi.12037 [ Links ]
Lasso, E., Engelbrecht, B. M. J., & Dalling, J. W. (2009). When sex is not enough: ecological correlates of resprouting capacity in congeneric tropical forest shrubs. Oecologia, 161(1), 43-56. [ Links ]
Li, J. T., Yang, J., Chen, D. C., Zhang, X. L., & Tang, Z. S. (2007). An optimized mini-preparation method to obtain high-quality genomic DNA from mature leaves of sunflower. Genetics and Molecular Research, 6(4), 1064-1071. [ Links ]
Lieberman, D., Lieberman, M., Peralta, R., & Hartshorn, G. (1996). Tropical forest structure and composition on a large-scale altitudinal gradient in Costa Rica. Journal of Ecology, 84(2), 137-152. [ Links ]
Loehle, C. (1987). Partitioning of reproductive effort in clonal plants: a benefit-cost model. Oikos, 49, 199-208. [ Links ]
Loveless, M. D., & Hamrick, J. L. (1984). Ecological determinants of genetic-structure in plant-populations. Annual Review of Ecology and Systematics, 15, 65-95. [ Links ]
McDade, L. A., & Hartshorn, G. S. (1994). La Selva biological station. In L.A. McDade, K. S. Bawa, H. A. Hespenheide, & G. S. Hartshorn (Eds.), La Selva: ecology and natural history of a neotropical rain forest (pp. 6-14). Chicago, Illinois: University of Chicago Press. [ Links ]
McKenna, D. D., & McKenna, K. M. (2006). Sesiid moths reduce germination, seedling growth, and survivorship in Pentaclethra macroloba (Mimosoideae), a locally dominant lowland neotropical tree. Biotropica, 38(4), 508-513. [ Links ]
Nakahama, N., Kaneko, S., Hayano, A., Isagi, Y., Inoue-Murayama, M., & Tominaga, T. (2012). Development of microsatellite markers for the endangered grassland species Vincetoxicum pycnostelma (Apocynaceae) by using next-generation sequencing technology. Conservation Genetics Resources, 4(3), 669-671. [ Links ]
Navascues, M., Stoeckel, S., & Mariette, S. (2010). Genetic diversity and fitness in small populations of partially asexual, self-incompatible plants. Heredity, 104(5), 482-492. [ Links ]
Oberbauer, S., & Strain, B. (1985). Effects of light regime on the growth and physiology of Pentaclethra macroloba (Mimosaceae) in Costa Rica. Journal of Tropical Ecology, 1(4), 303-320. [ Links ]
Oberbauer, S., & Strain, B. (1986). Effects of canopy position and irradiance on the leaf physiology and morphology of Pentaclethra macroloba (Mimosaceae). American Journal of Botany, 73(3), 409-416. [ Links ]
Oberbauer, S., Strain, B., & Riechers, G. (1987). Field water relations of a wet-tropical forest tree species, Pentaclethra macroloba (Mimosaceae). Oecologia, 71, 369-374. [ Links ]
Peakall, R., & Smouse, P. E. (2006). GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes, 6(1), 288-295. [ Links ]
Peh, K. S. H., Lewis, S. L., & Lloyd, J. (2011). Mechanisms of monodominance in diverse tropical tree-dominated systems. Journal of Ecology, 99(4), 891-898. [ Links ]
Perry, D. R. (1983). Access methods, observations, pollination biology, bee foraging behavior, and bee community structure within a neotropical wet forest canopy. Los Angeles: University of California, [ Links ]
Putz, F., & Brokaw, N. (1989). Sprouting of broken trees on Barro Colorado Island, Panama. Ecology, 70(2), 508-512. [ Links ]
Raymond, M., & Rousset, F. (1995). Genepop (Version-1.2): population-genetics software for exact tests and ecumenicism. Journal of Heredity, 86(3), 248-249. [ Links ]
Richards, P. W. (1952). The tropical rain forest, an ecological study. Cambridge University Press, Cambridge. [ Links ]
Russell, A. E., Raich, J. W., Arrieta, R. B., Valverde-Barrantes, O., & González, E. (2010). Impacts of individual tree species on carbon dynamics in a moist tropical forest environment. Ecological Applications, 20(4), 1087-1100. [ Links ]
Salk, C. F., & McMahon, S. M. (2011). Ecological and environmental factors constrain sprouting ability in tropical trees. Oecologia, 166(2), 485-492. [ Links ]
Sawicki, J., Kwasniewski, M., Szczecinska, M., Chwialkowska, K., Milewicz, M., & Plasek, V. (2012). Isolation and Characterization of Simple Sequence Repeats (SSR) Markers from the Moss Genus Ortho-trichum Using a Small Throughput Pyrosequencing Machine. International Journal of Molecular Sciences, 13(6), 7586-7593. [ Links ]
Schierenbeck, K. A., Brunsfeld, S. J., Finegan, B., & Waits, L. M. (1997). Population structure and genetic diversity in four tropical tree species in Costa Rica. Molecular Ecology, 6(2), 137-144. [ Links ]
Sherman, C. D. H., Stanley, A. M., Keough, M. J., Gardner, M. G., & Macreadie, P. I. (2012). Development of twenty-three novel microsatellite markers for the seagrass, Zostera muelleri from Australia. Conservation Genetics Resources, 4(3), 689-693. [ Links ]
Smythe, N. (1970). Relationships between fruiting seasons and seed dispersal methods in a neotropical forest. The American Naturalist, 104(935), 25-35. [ Links ]
Stoeckel, S., Grange, J., Fernández-Manjarres, J. F., Bilger, I., Frascaria-Lacoste, N., & Mariette, S. (2006). Hete-rozygote excess in a self-incompatible and partially clonal forest tree species -Prunus avium L. Molecular Ecology, 15(8), 2109-2118. [ Links ]
Terborgh, J. (2012). Enemies maintain hyperdiverse tropical forests. The American Naturalist, 179(3), 303-314. [ Links ]
ter Steege, H., Pitman, N. C., Sabatier, D., Baraloto, C., Salomão, R. P., Guevara, J. E., & Fine, P. V. (2013). Hyperdominance in the Amazonian Tree Flora. Science, 342(6156), 1243092. doi:10.1126/ science.1243092 [ Links ]
Torti, S. D., Coley, P. D., & Kursar, T. A. (2001). Causes and consequences of monodominance in tropical lowland forests. The American Naturalist, 157(2), 141-153. [ Links ]
Van Oosterhout, C., Hutchinson, W. F., Wills, D. P. M., & Shipley, P. (2004). Micro-Checker: Software for Identifying and Correcting Genotyping Errors in Microsatellite Data. Molecular Ecology Notes, 4(3), 535-538. [ Links ]
Whitmore, T. C., & Burnham, C. P. (1975). Tropical rain forests of the Far East (p. 295). Clarendon, Oxford. [ Links ]
Williamson, G. B., & Costa, F. (2000). Dispersal of amazonian trees: Hydrochory in Pentaclethra macroloba. Biotropica, 32(3), 548-552. [ Links ]
Wright, S. J. (2002). Plant diversity in tropical forests: a review of mechanisms of species coexistence. Oecologia, 130(1), 1-14. [ Links ]
Beard, J. S. (1946). The Mora Forests of Trinidad, British WestIndies. Journal of Ecology, 33(2), 173-192. [ Links ]
Boutin-Ganache, I., Raposo, M., Raymond, M., & Deschepper, C. F. (2001). M13-tailed primers improve the readability and usability of microsatellite analyses performed with two different allele-sizing methods. Biotechniques, 31(1), 24-28. [ Links ]
Clark, D. B., & Clark, D. A. (2000). Landscape-scale variation in forest structure and biomass in a tropical rain forest. Forest Ecology and Management, 137(1-3), 185-198. [ Links ]
Clark, D. B, Olivas, P. C., Oberbauer, S. F., Clark, D. A, & Ryan, M. G. (2008). First direct landscape-scale measurement of tropical rain forest Leaf Area Index, a key driver of global primary productivity. Ecology Letters, 11(2), 163-172. [ Links ]
Clark, D. B., & Clark, D. A. (2006). Tree growth, mortaity, physical condition, and microsite in an old-growth lowland tropical rain forest. Ecology, 87(8), 2132. [ Links ]
Connell, J., & Lowman, M. (1989). Low-diversity tropical rain forests: some possible mechanisms for their existence. The American Naturalist, 134(1), 88-119. [ Links ]
Cullings, K. W. (1992). Design and testing of a plant-specific PCR primer for ecological and evolutionary studies. Molecular Ecology, 1(4), 233-240. [ Links ]
Doyle, J. J., & Doyle, J. L. (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin, 19, 11-15. [ Links ]
Faircloth, B. C. (2008). MSATCOMMANDER: detection of microsatellite repeat arrays and automated, locus-specific primer design. Molecular Ecology Resources, 8(1), 92-94. [ Links ]
Fatemi, M., Haddadchi, A., & Gross, C. L. (2012). The discovery of microsatellite markers for Hardenbergia violacea (Fabaceae), using next-generation sequencing. Conservation Genetics Resources, 4(4), 1063-1065. [ Links ]
Flores, E. M. (2004). Pentaclethra macroloba. Academia Nacional de Ciencies de Costa Rica. [ Links ]
Guariguata, M. R., Adame, J. J. R., & Finegan, B. (2000). Seed removal and fate in two selectively logged lowland forests with constrasting protection levels. Conservation Biology, 14(4), 1046-1054. [ Links ]
Hall, P., Chase, M. R., & Bawa, K. S. (1994). Low genetic-variation but high population differentiation in a common tropical forest tree species. Conservation Biology, 8(2), 471-482. [ Links ]
Hanson, T. R., Brunsfeld, S. J., Finegan, B., & Waits, L. P. (2008). Pollen dispersal and genetic structure of the tropical tree Dipteryx panamensis in a fragmented Costa Rican landscape. Molecular Ecology, 17(8), 2060-2073. [ Links ]
Hart, T. B. (1990). Monospecific dominance in tropical rain forests. Trends in Ecology & Evolution, 5(1), 6-11. [ Links ]
Hartshorn, G. S. (1972). The ecological life history and population dynamics of Pentaclethra macroloba, a wet forest dominant, and Stryphnodendron excelsum, and occasional associate. University of Washington. [ Links ]
Hartshorn, G. S. (1975). A matrix model of tree population dynamics. In F. B. Golley & E. Medina (Eds.), Tropical ecological systems (pp. 41-51). New York: Springer-Verlag. [ Links ]
Hartshorn, G. S., & Hammel, B. E. (1994). Vegetation types and floristic patterns. In La Selva: Ecology and natural history of a neotropical rain forest (pp. 73-89). Chicago, Illinois: University of Chicago Press. [ Links ]
Janos, D. (1980). Vesicular-arbuscular mycorrhizae affect lowland tropical rain forest plant growth. Ecology, 61(1), 151-162. [ Links ]
Jones, F. A., Cerón-Souza, I., Hardesty, B. D., & Dick, C. W. (2013). Genetic evidence of Quaternary demographic changes in four rain forest tree species sampled across the Isthmus of Panama. Journal of Biogeography, 40(4), 720-731. doi:10.1111/jbi.12037 [ Links ]
Lasso, E., Engelbrecht, B. M. J., & Dalling, J. W. (2009). When sex is not enough: ecological correlates of resprouting capacity in congeneric tropical forest shrubs. Oecologia, 161(1), 43-56. [ Links ]
Li, J. T., Yang, J., Chen, D. C., Zhang, X. L., & Tang, Z. S. (2007). An optimized mini-preparation method to obtain high-quality genomic DNA from mature leaves of sunflower. Genetics and Molecular Research, 6(4), 1064-1071. [ Links ]
Lieberman, D., Lieberman, M., Peralta, R., & Hartshorn, G. (1996). Tropical forest structure and composition on a large-scale altitudinal gradient in Costa Rica. Journal of Ecology, 84(2), 137-152. [ Links ]
Loehle, C. (1987). Partitioning of reproductive effort in clonal plants: a benefit-cost model. Oikos, 49, 199-208. [ Links ]
Loveless, M. D., & Hamrick, J. L. (1984). Ecological determinants of genetic-structure in plant-populations. Annual Review of Ecology and Systematics, 15, 65-95. [ Links ]
McDade, L. A., & Hartshorn, G. S. (1994). La Selva biological station. In L.A. McDade, K. S. Bawa, H. A. Hespenheide, & G. S. Hartshorn (Eds.), La Selva: ecology and natural history of a neotropical rain forest (pp. 6-14). Chicago, Illinois: University of Chicago Press. [ Links ]
McKenna, D. D., & McKenna, K. M. (2006). Sesiid moths reduce germination, seedling growth, and survivorship in Pentaclethra macroloba (Mimosoideae), a locally dominant lowland neotropical tree. Biotropica, 38(4), 508-513. [ Links ]
Nakahama, N., Kaneko, S., Hayano, A., Isagi, Y., Inoue-Murayama, M., & Tominaga, T. (2012). Development of microsatellite markers for the endangered grassland species Vincetoxicum pycnostelma (Apocynaceae) by using next-generation sequencing technology. Conservation Genetics Resources, 4(3), 669-671. [ Links ]
Navascues, M., Stoeckel, S., & Mariette, S. (2010). Genetic diversity and fitness in small populations of partially asexual, self-incompatible plants. Heredity, 104(5), 482-492. [ Links ]
Oberbauer, S., & Strain, B. (1985). Effects of light regime on the growth and physiology of Pentaclethra macroloba (Mimosaceae) in Costa Rica. Journal of Tropical Ecology, 1(4), 303-320. [ Links ]
Oberbauer, S., & Strain, B. (1986). Effects of canopy position and irradiance on the leaf physiology and morphology of Pentaclethra macroloba (Mimosaceae). American Journal of Botany, 73(3), 409-416. [ Links ]
Oberbauer, S., Strain, B., & Riechers, G. (1987). Field water relations of a wet-tropical forest tree species, Pentaclethra macroloba (Mimosaceae). Oecologia, 71, 369-374. [ Links ]
Peakall, R., & Smouse, P. E. (2006). GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Molecular Ecology Notes, 6(1), 288-295. [ Links ]
Peh, K. S. H., Lewis, S. L., & Lloyd, J. (2011). Mechanisms of monodominance in diverse tropical tree-dominated systems. Journal of Ecology, 99(4), 891-898. [ Links ]
Perry, D. R. (1983). Access methods, observations, pollination biology, bee foraging behavior, and bee community structure within a neotropical wet forest canopy. Los Angeles: University of California, [ Links ]
Putz, F., & Brokaw, N. (1989). Sprouting of broken trees on Barro Colorado Island, Panama. Ecology, 70(2), 508-512. [ Links ]
Raymond, M., & Rousset, F. (1995). Genepop (Version-1.2): population-genetics software for exact tests and ecumenicism. Journal of Heredity, 86(3), 248-249. [ Links ]
Richards, P. W. (1952). The tropical rain forest, an ecological study. Cambridge University Press, Cambridge. [ Links ]
Russell, A. E., Raich, J. W., Arrieta, R. B., Valverde-Barrantes, O., & González, E. (2010). Impacts of individual tree species on carbon dynamics in a moist tropical forest environment. Ecological Applications, 20(4), 1087-1100. [ Links ]
Salk, C. F., & McMahon, S. M. (2011). Ecological and environmental factors constrain sprouting ability in tropical trees. Oecologia, 166(2), 485-492. [ Links ]
Sawicki, J., Kwasniewski, M., Szczecinska, M., Chwialkowska, K., Milewicz, M., & Plasek, V. (2012). Isolation and Characterization of Simple Sequence Repeats (SSR) Markers from the Moss Genus Ortho-trichum Using a Small Throughput Pyrosequencing Machine. International Journal of Molecular Sciences, 13(6), 7586-7593. [ Links ]
Schierenbeck, K. A., Brunsfeld, S. J., Finegan, B., & Waits, L. M. (1997). Population structure and genetic diversity in four tropical tree species in Costa Rica. Molecular Ecology, 6(2), 137-144. [ Links ]
Sherman, C. D. H., Stanley, A. M., Keough, M. J., Gardner, M. G., & Macreadie, P. I. (2012). Development of twenty-three novel microsatellite markers for the seagrass, Zostera muelleri from Australia. Conservation Genetics Resources, 4(3), 689-693. [ Links ]
Smythe, N. (1970). Relationships between fruiting seasons and seed dispersal methods in a neotropical forest. The American Naturalist, 104(935), 25-35. [ Links ]
Stoeckel, S., Grange, J., Fernández-Manjarres, J. F., Bilger, I., Frascaria-Lacoste, N., & Mariette, S. (2006). Hete-rozygote excess in a self-incompatible and partially clonal forest tree species -Prunus avium L. Molecular Ecology, 15(8), 2109-2118. [ Links ]
Terborgh, J. (2012). Enemies maintain hyperdiverse tropical forests. The American Naturalist, 179(3), 303-314. [ Links ]
ter Steege, H., Pitman, N. C., Sabatier, D., Baraloto, C., Salomão, R. P., Guevara, J. E., & Fine, P. V. (2013). Hyperdominance in the Amazonian Tree Flora. Science, 342(6156), 1243092. doi:10.1126/ science.1243092 [ Links ]
Torti, S. D., Coley, P. D., & Kursar, T. A. (2001). Causes and consequences of monodominance in tropical lowland forests. The American Naturalist, 157(2), 141-153. [ Links ]
Van Oosterhout, C., Hutchinson, W. F., Wills, D. P. M., & Shipley, P. (2004). Micro-Checker: Software for Identifying and Correcting Genotyping Errors in Microsatellite Data. Molecular Ecology Notes, 4(3), 535-538. [ Links ]
Whitmore, T. C., & Burnham, C. P. (1975). Tropical rain forests of the Far East (p. 295). Clarendon, Oxford. [ Links ]
Williamson, G. B., & Costa, F. (2000). Dispersal of amazonian trees: Hydrochory in Pentaclethra macroloba. Biotropica, 32(3), 548-552. [ Links ]
Wright, S. J. (2002). Plant diversity in tropical forests: a review of mechanisms of species coexistence. Oecologia, 130(1), 1-14. [ Links ]
*Correspondencia a:
Keith D. Gaddis. Department of Ecology and Evolutionary Biology, University of California, 612 Charles E. Young Drive East, Los Angeles, California 90095-7239 USA. Correo electrónico kgaddis@ucla.edu
Helen L. Zukin. Department of Biology, Occidental College, 1600 Campus Road, Los Angeles CA 90041 USA. Correo electrónico. Correo electrónico hlzukin@gmail.com
Inca A. Dieterich. Department of Biology, Occidental College, 1600 Campus Road, Los Angeles CA 90041 USA. Correo electrónico inca.dieterich@gmail.com
Elizabeth Braker. Department of Biology, Occidental College, 1600 Campus Road, Los Angeles CA 90041 USA. Correo electrónico bbraker@oxy.edu
Victoria L. Sork. Department of Ecology and Evolutionary Biology, University of California, 612 Charles E. Young Drive East, Los Angeles, California 90095-7239 USA. Institute of the Environment and Sustainability, University of California, 619 Charles E. Young Drive East, Los Angeles, California 90095-1496 USA. Correo electrónico vlsork@ucla.edu
1. Department of Ecology and Evolutionary Biology, University of California, 612 Charles E. Young Drive East, Los Angeles, California 90095-7239 USA. Correo electrónico kgaddis@ucla.edu
2. Department of Biology, Occidental College, 1600 Campus Road, Los Angeles CA 90041 USA. Correo electrónico bbraker@oxy.edu, hlzukin@gmail.com, inca.dieterich@gmail.com
3. Institute of the Environment and Sustainability, University of California, 619 Charles E. Young Drive East, Los Angeles, California 90095-1496 USA. Correo electrónico vlsork@ucla.edu
Received 20-viii-2013. Corrected 09-Xii-2013. Accepted 16-i-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}