











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.62 no.2 San José abr./jun. 2014
Impact of forest fragment size on the population structure of three palm species (Arecaceae) in the Brazilian Atlantic Rainforest
Impacto del tamaño del fragmento de bosque en la estructura de la población de tres especies de palmas del Bosque Atlántico Brasileño
Impacto del tamaño del fragmento de bosque en la estructura de la población de tres especies de palmas del Bosque Atlántico Brasileño
*Dirección para correspondencia:
Abstract
The main threats to natural populations in terrestrial ecosystems have been widly recognized to be the habitat fragmentation and the exploitation of forest products. In this study, we compared the density of the populations and the structure of three tropical palm species, Astrocaryum aculeatissimum, Euterpe edulis and Geonoma schottiana. For this, we selected five forest fragments of different sizes (3 500ha, 2 400ha, 57ha, 21ha and 19ha) where palms were censused in nine 30x30m plots. We tracked the palms survival from 2005 to 2007, and recorded all new individuals encountered. Each individual was assigned in one of the five ontogenetic stages: seedling, infant, juvenile, immature and reproductive. The demographic structure of each palm species was analyzed and compared by a generalized linear model (GLM). The analysis was performed per palm spe-cies. The forest fragment area and the year of observation were explanatory variables, and the proportion of individuals in each ontogenetic class and palm density were response variables. The total number of individuals (from seedlings to reproductives, of all species) monitored was 6 450 in 2005, 7 268 in 2006, and 8 664 in 2007. The densities of two palm species were not influenced by the size of the fragment, but the population density of A. aculeatissimum was dependent on the size of the fragment: there were more individuals in the bigger than in the smaller forest fragments. The population structure of A. aculeatissimum, E. edulis, and G. schottiana was not altered in the smaller fragments, except the infants of G. schottiana. The main point to be drawn from the results found in this study is that the responses of density and population structure seem not to be dependent on fragment size, except for one species that resulted more abundant in bigger fragments.
Key words: population structure, population density, Astrocaryum aculeatissimum, Euterpe edulis, Geonoma schottiana
Resumen
Uno de los principales riesgos que corren las poblaciones naturales de los ecosistemas terrestres es la fragmentación de hábitat y la explotación de los recursos forestales. Aquí hemos comparado la densidad de las poblaciones y el estado estructural de tres especies de palmeras, Astrocaryum aculeatissimum (Schott) Burret, Euterpe edulis Mart. y Geonoma schottiana Mart., en fragmentos de diferente tamaño. Las palmas fueron censadas en nueve cuadrantes de 30x30m en cinco fragmentos de bosque de diferente tamaño (3 500ha, 2 400ha, 57ha, 21ha y 19ha). Realizamos el seguimiento de la sobrevivencia de las palmas entre 2005-2007, y registramos todos los nuevos individuos encontrados. Cada individuo fue designado dentro de alguno de los siguientes cinco estadíos ontogenéticos: plántula, infantil, juvenil, inmaduro y reproductivo. La comparación de la estructura demográfica fue analizada usando Modelos Lineares Generalizados (GLM). Estos fueron utilizados para cada especie por separado, con el área del fragmento de bosque y el año como variables explicativas, así como proporción de individuos en cada estadío ontogenético y densidad de las palmeras como variables de respuesta. El número total de individuos monitoreado (desde semillas hasta adultos reproductivos, para todas las especies) fue 6 450 en 2005, 7 268 en 2006 y 8 664 en 2007. La densidad de población para dos especies de palmas no fue influenciada por el tamaño del fragmento, excepto para Astrocaryum aculeatissimum, cuya densidad dependió del tamaño del fragmento. Hubo más individuos en los fragmentos mayores que en los fragmentos menores. La estructura de la población de A. aculeatissimum, E. edulis y G. schottiana no se vio alterada en los fragmentos pequeños, excepto para G. schottiana en estado infante. El punto principal que se desprende de los resultados encontrados en este estudio es que la respuesta de la densidad y estructura de la población parece no ser dependiente del tamaño de fragmento, excepto para una especie que es más abundante en los fragmentos mayores.
Palabras clave: estructura de la población, densidad de la población, Astrocaryum aculeatissimum, Euterpe edulis, Geonoma schottiana.
One of the main threats to natural populations in terrestrial ecosystems is habitat fragmentation (Tabarelli & Gascon, 2005; Ewers & Didham, 2006; Melo, Arroyo-Rodríguez, Fahrig, Martínez-Ramos & Tabarelli, 2013). In the tropics, high deforestation rates have converted formerly continuous rainforest to anthropogenic landscapes, where forest habitats have been reduced to scattered forest fragments of varying sizes and quality (Ranta, Blom, Niemelã, Elina & Sitonem, 1998; Fischer & Lindenmayer, 2007). Both experimental and observational studies about the effects of forest fragmentation in plant populations, have demonstrated that the biotic and abiotic changes associated with fragmentation, can dramatically alter the fertility, growth, and survivorship of the remaining plant populations (Bruna, Fiske & Trager, 2009). Following forest fragmentation, the remaining forest usually suffers degradation by timber harvest, the extraction of non-timber products, like leaves, fruits, and palm heart, and weed invasion (Ewers & Didham, 2006; Melo et al., 2013). The ability of a population to tolerate a certain level of exploitation is strongly influenced by the parts of the plant that are harvested, and the species life history particularities (Rodríguez-Buriticá, Orjuela & Galeano, 2005). These alterations, habitat fragmentation, and the harvesting of non-timber products, frequently result in the loss of habitat quality and species. Such is the case of the Atlantic Rainforest in Southeastern Brazil, where most of the forest has been converted to other land uses, thereby endangering endemic flora and fauna (Giulietti, Harley, Queiroz, Wanderley & Van den Berg, 2005). The Brazilian Atlantic Rainforest has experienced intense urbanization since the 1960s, and today 70% of the Brazilian population lives in the region (Melo et al., 2013).
It is frequently hypothesized that plant populations in forest fragments will be reduced in size, exhibit an altered population structure, and show an increased probability of extinction by demographic, environmental, and genetic stochasticity (Jules, 1998). One current view in conservation biology is that the conservation of any particular species in nature will only be successful through the maintenance of its former populations (Primack, 1995). The extinction of a species may occur through the successive extirpations of its population, turned locally unsustainable after direct and indirect changes in habitat quality (Fahrig, 2002; Souza & Martins, 2004, Ewers & Didham, 2006).
Despite the recognized conservation value of forest remnants (Fischer & Lindenmayer, 2007; Santos, Kinoshita & Santos, 2007; Melo et al., 2013), and the well-established threats that fragmentation poses to natural populations, surprisingly few studies have devoted attention to the population ecology of long-lived plant species in tropical forest fragments (Silva-Matos, Freckleton, & Watkinson, 1999; Souza & Martins, 2002; Bruna & Kress, 2002; Bruna, 2003; Bruna & Oli, 2005; Souza 2007). Among other reasons, such studies are needed because long-lived, iteroparous plants may build up remnant populations under conditions where the completion of the whole life cycle is not possible (Eriksson, 1996). Such populations may respond favorably to habitat improvement under management policies, but are condemned if unfavorable conditions, represented by forest degradation, are prolonged for more than the lifetime of its individuals (Souza & Martins, 2004). Also, obtaining information on the consequences of fragmentation for populations persisting in fragmented tropical landscapes is necessary to derive effective conservation strategies for these species-rich regions (Laurance & Bierregaard, 1997).
Despite the use of population structure, some previous studies considered that it represents a poor predictor of population performance (Condit, Sukumar, Hubbell & Foster, 1998). In a conservation setting, where long-term demographic monitoring may not be feasible, a population’s stage structure can be the basis for immediate management decisions (Bruna & Kress, 2002). In fact, Schemske, Husband, Ruckelshaus, Goodwillie, Parker & Bishop (1994) suggested that a good starting point for the recovery of threatened plant species is an assessment of the current demographic structure of the populations of interest.
In this paper, we compared the density of the populations and the stage structure of three tropical palm species: Astrocaryum aculeatis-simum (Schott) Burret, Euterpe edulis Mart. (palmito Juçara), and Geonoma schottiana Mart. (“ouricana”). These species are endemic, except G. schottiana, and are palms that are abundant in the Brazilian Atlantic Rainforest. Specifically, we tested the following two predictions: a) densities of A. aculeatissimum, E. edulis and G. schottiana are reduced in the smaller forest fragments relative to larger ones, suggesting reductions in population sizes due to either episodic catastrophes or ongoing demographic decline; and b) the demographic structure of populations is altered in the smaller forest fragments, specially due to a negative impact on the seedlings.
Material and Methods
Study site: The study was carried out in five areas of lowland Atlantic Rainforest (“floresta pluvial baixo montana”, according to Rizzini, 1979), two reserves (big fragments): the National Biological Reserve of Poço das Antas (fragment 1–ca. 3 500ha) and the National Biological Reserve of União (fragment 2–ca. 2 400ha), and three private fragments (small fragments): Santa Helena (fragment 3–ca. 57ha), Estreito (fragment 4–ca. 21ha), and Afetiva-Jorge (fragment 5–ca. 19ha). They are located in the municipalities of Silva Jardim, Casimiro de Abreu, and Rio das Ostras, state of Rio de Janeiro, Southeastern Brazil (22°30’22.33”S - 42°15’42.19”W). Details on the vegetation structure of the fragments can be found in Carvalho and Nascimento (2009). The two reserves and the three fragments are surrounded by pasture, agricultural fields, and secondary forest. The climate is classified as Walter and Lieth’s Equatorial type (Walter, 1971), with an average annual temperature of 24.6°C and mean annual rainfall of ca. 2 100mm (1987-1997 data, Souza & Martins, 2004). There is no distinct dry season; however, despite large variations between years, a drier period normally occurs from May to August.
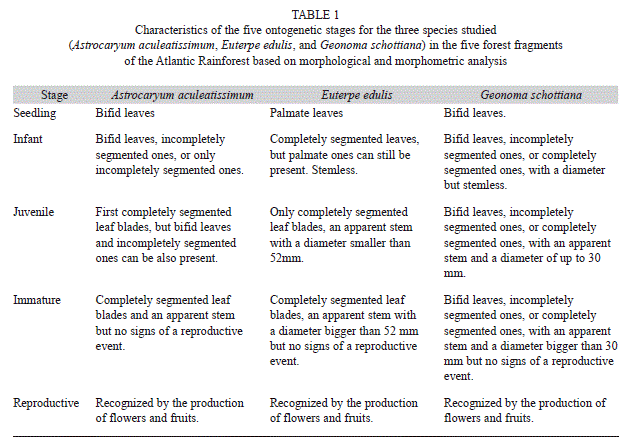
Study species: The three studied species were chosen because they are present in all five fragments. A. aculeatissimum is a monoecious, shade-tolerant, slow-growing, solitary or multi-stemmed palm with abundant black spines in the stem (Henderson, Galeano, & Bernal, 1995; Lorenzi, Souza, Medeiros-Costa, Cerqueira & Ferreira, 2004). It is typically 4-8m in height and 11-15cm in diameter (Henderson, Galeano, & Bernal, 1995; Lorenzi et al., 2004). This species is endemic to the Southeastern Atlantic Rainforest, occurring from Bahia to Santa Catarina (Henderson, Galeano, & Bernal, 1995; Lorenzi et al., 2004). It grows in the understory of lowland forest, and is rarely found in flooded sites and occasionally in the matrix surrounding forest fragments. E. edulis is a monoecious, solitary, shade-tolerant, and slow-growing palm. It is a subcanopy palm, can reach 20m in height and 10-15cm in diameter (Henderson, Galeano, & Bernal, 1995; Lorenzi et al., 2004). It occurs in forests primarily along the Atlantic coast of Brazil, reaching inland at least to Brasilia and just reaching Argentina and Paraguay (Henderson, Galeano, & Bernal, 1995). It occupies the crests or slopes of hills and flooded sites up to 1 000m elevation (Henderson, Galeano, & Bernal, 1995). This species is harvested for its palm heart and it is one of the most abundant and valuable non-timber forest products in the Atlantic Rainforest (Fantini & Guris, 2007). Intensive harvesting has led to the decline of the palm over much of the region; many of the surviving populations are small and fragmented (Galetti & Aleixo, 1998; Silva-Matos, Freckleton, & Watkinson, 1999). G. schottiana is a monoecious, solitary or rarely multi-stemmed species, and is a shade-tolerant and slow-growing palm (Henderson, Galeano, & Bernal, 1995; Lorenzi et al., 2004). It is typically 1-4m in height, 2.5-4cm in diameter, and occupies the forest understory of lowland forest (Henderson, Galeano, & Bernal, 1995; Lorenzi et al., 2004). This species occurs in Espírito Santo, Rio de Janeiro, São Paulo, Paraná, Santa Catarina, Minas Gerais and Goiás, in the Atlantic Rainforest and in Cerrado (Henderson, Galeano, & Bernal, 1995; Lorenzi et al., 2004). In the studied region, the leaves of G. schottiana are currently harvested for floral arrangements. There are two methods of harvesting the leaves: cutting the stem of the plant (causing mortality), or removing only the leaves (personal observation).
Based on morphological and morphometric analysis, we distinguished five stages for each palm species (Table 1). The meaning of the stage names for each studied species is described and discussed in Portela and Santos (2011).
Demographic plots and censuses: Palms were censused in nine 30x30m plots in each fragment, distributed systematically in three blocks, 100m apart to each other. Each block had three plots that were 50m separated from each other. A total of 0.81ha was censused in each fragment. One block was established in the middle of each fragment and the other two blocks were established on the sides of the first block. For the two National Biological Reserves, we used a previously established trail close to the center of the fragment; all other methods were the same. All individuals of the three palm species were tagged with a numbered aluminum label between June and September of 2005. The survivorship of the plants was subsequently monitored between June and September of 2006 and 2007. All new plants were also tagged. Each individual was assigned to one of the five abovementioned stages. The density of each species was estimated using the total number of individuals in the nine plots, a total of 0.81ha, for each year. During June-September of 2007, the number of E. edulis and G. schottiana individuals inside the plots was counted to estimate the intensity of harvesting in each fragment.
The comparison of the density and demographic structure of each palm was analyzed by Generalized linear models (GLM). The analysis was performed per palm species. The forest fragment area and year of observation were explanatory variables, and the proportion of individuals in each ontogenetic class and palm density were response variables. The population density of A. aculeatissimum, E. edulis, and G. schottiana was adjusted to a quasi-pois-son distribution. The proportion of seedlings and immatures of A. aculeatissimum, infants, juveniles, and reproductives of E. edulis, and infants, juveniles, immatures, and reproductives of G. schottiana was adjusted to a binomial distribution; the proportion of infants, juveniles, and reproductives of A. aculeatissimum, seedlings and immatures of E. edulis, and seedlings of G. schottiana was adjusted to a quasi-binomial distribution. Analyses were implemented in the software package R 2.15.1 (R Development Core Team, 2012).
Results
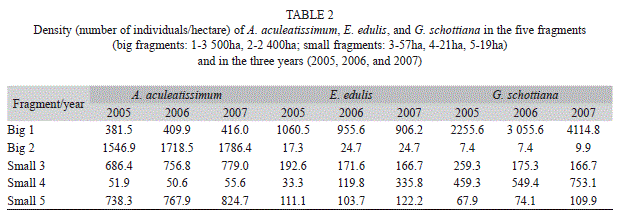
Plant density: The population density of A. aculeatissimum was dependent on the size of the forest fragment (t=-3.831, p=0.0028), but was not dependent on the year (t=1.234, p=0.24302). There were more individuals in the bigger fragments than in the smaller fragments (Table 2). The population density of E. edulis and G. schottiana was not dependent on the size of the forest fragment (t=-1.398, p=0.190; t=-0.704, p=0.496, respectively) or on the year (t=-0.321, p=0.754; t=0.753, p=0.467, respectively).
The density E. edulis and G. schottiana did not show a relationship with fragment size, but the two palm species had a higher density in at least one of the bigger fragments compared with the smaller ones (Table 2). None of the three species had a higher density in the smaller fragments.
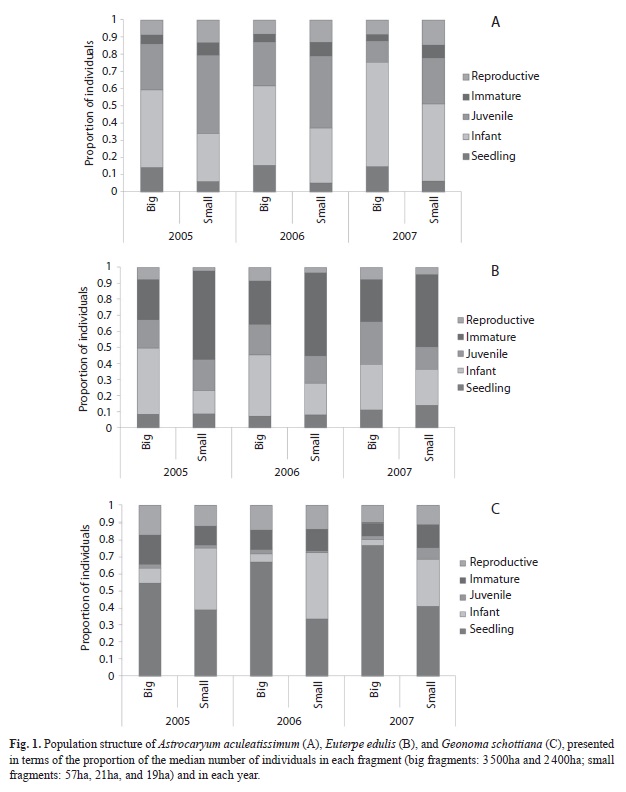
Population structure (Fig. 1): The proportion of A. aculeatissimum individuals in the seedling (Z=-0.722, p=0.4702), infant (Z=-0.368, p=0.720), juvenile (Z=0.686, p=0.507), immature (Z=0.211, p=0.833), and reproductive (Z=0.456, p=0.657) ontogenetic stages was not dependent on the size of the fragment. It was not dependent neither on the year: seed-lings (Z=0.091, p=0.9276), infants (Z=-0.582, p=0.572), juveniles (Z=-0.689, p=0.507), immatures (Z=-0.194, p=0.847), and reproductives (Z=-0.040, p=0.969).
Similar results were found for E. edulis for which the proportion of its individuals during the seedling (T=-0.803, p=0.439), infant (Z=-1.950, p=0.0511), juvenile (Z=1.570, p=0.1163), immature (Z=2.019, p=0.0686), and reproductive (Z=-0.163, p=0.871) onto-genetic stages was not dependent on the size of the fragment, and neither dependent on the year: seedlings (T=-0.229, p=0.823), infants (Z=-1.788, p=0.0738), juveniles (Z=1.742, p=0.0816), immatures (Z=0.367, p=0.7204), and reproductives (Z=-0.133, p=0.894).
For G. schottiana, the proportion of individuals in the seedling (T=-0.748, p=0.4700), juvenile (Z=-0.086, p=0.932), immature (Z=-1.470, p=0.14152), and reproductive (Z=-0.920, p=0.3578) ontogenetic stages was not dependent on the size of the fragment. However, the proportion of infants (Z=4.619, p=0.000) varied according to the size of the fragment, showing higher values in the smaller fragments. Additionally, the population structure was dependent on the year: infants (Z=-1.298, p=0.1943), juveniles (Z=-0.068, p=0.946), and reproductives (Z=-1.849, p=0.0644), except for seedlings (T=5.876, p=0.000) and immatures (Z=-2.677, p=0.00744), which varied, especially during the first year when compared with the other two years.
Harvesting: In the studied areas, only A. aculeatissimum individuals were not harvested; nevertheless, the palm heart of E. edulis (with the subsequent death of the individual) and the leaves of G. schottiana were under harvesting activities. We only found this harvesting in the three private fragments (Table 3). In fragment 4, we did not find any harvesting activity in the plots, but there were some harvested individuals outside the plots. For G. schottiana, harvesting of leaves rarely causes mortality, however, sometimes the whole crown is cut (Table 3), but usually, the oldest leaves were cut and the newest leaves were left. Figure 1
Discussion
The density of A. aculeatissimum was the only one affected by the size of the forest fragment; the other two species were not directly influenced. The population structure of A. aculeatissimum, E. edulis, and G. schottiana was not altered in the smaller fragments, except the infants of G. schottiana. Therefore, one may conclude that a neutral effect of fragment size on the population structure was found for these three palm species.
The densities of palms reported in the literature are much higher than those we have found. Sampaio and Scariot (2010, G. schot-tiana) and Reis et al. (2000, E. edulis), who studied the same species, found a much higher density. Rodríguez-Buriticá et al. (2005), who studied another Geonoma species that was lightly harvested, found a higher density than the one we found in our study. Souza and Martins (2004), who studied other palm species in the same region, found a similar density for some populations and a higher density for others.
The three palms had a higher density in at least one of the bigger fragments when compared to the smaller ones, although a statistically significant difference was only found for A. aculeatissimum. None of the three species had a higher density in one of the smaller fragments. A similar result was previously reported by Arroyo-Rodríguez, Aguirre, Benítez-Malvido & Mandujano (2007), who studied Astrocaryum mexicanum; they determined that smaller and more irregular fragments presented lower reproductive densities. Chazdon (1996) showed that, in addition to natural factors like soil, topography, and humidity, human activity and historical perturbation also influenced the distribution and the dynamics of palm populations. Additionally, the species responses to habitat fragmentation are governed by the traits of individual species, like dispersal abilities (Ewers & Didham, 2006).
Considering the size of the fragments, the two bigger fragments were Reserves, and no harvesting of E. edulis and G. schottiana was permitted. Fragment 1 had been a protected reserve since 1975, and fragment 2 since 1998. However, neither the fragment size nor the protected status, influenced the population size of these two palm species. On the other hand, A. aculeatissimum is not a harvested species and has clonal growth (more than 50% of the population are clonal, Portela & Santos, 2011), and its density is negatively affected by the reduction of habitat or modification of forest structure that comes with the fragmentation. Galetti, Donatti, Pires, Guimarães & Jordano (2006), also studying A. aculeatissimum, concluded that a large fraction of the Atlantic Rainforest palms that relied on scatter-hoarding rodents might become regionally extinct due to forest fragmentation and defaunation. The reduction in A. aculeatissimum population size in the smaller fragments could be a response of the absence of its seed dispersers and, consequently, a reduction in recruitment. The effect of the loss of seed dispersers can be especially pervasive for plants that rely on few frugivore species for seed dispersal, such as large-seeded palms like A. aculeatissimum. Other studies have also reported the absence of large-bodied dispersers in small fragments and the relatively high abundance of small seed predators (Chinchilla, 2009). The other two palm species also depend on animals to disperse theirs seeds, and some seed dispersers have been reported as birds, ranging from small to large animals (Galetti et al., 2013).
The population structure of A. aculeatis-simum, E. edulis, and G. schottiana was not altered in the smaller fragments and did not vary between the studied years. The Brazilian Atlantic Rainforest has been drastically devastated since the beginning of the European colonization (Dean, 1996). Also, according to reports from former residents of the studied region, obtained and described by Carvalho and Nascimento (2009), the fragments were part of a continuous forest until about a century ago. The studied palm species are forest-dependent species (Henderson, Galeano, & Bernal, 1995; Lorenzi et al., 2004); nevertheless, their population structure was not affected by forest fragmentation. These populations may be in equilibrium in the studied landscape, since the area has been fragmented since the last century. Despite Ewers and Didham (2006), in their revision about species’ responses to habitat fragmentation, reporting that time-scales of 50-90 years after fragmentation, may not be sufficient to ensure that species have reached equilibrium. Nevertheless, results on the population structure of the three palm species in an old fragmented area seem to suggest that they are in equilibrium. The main point to be drawn from the results of this study is that the density responses (except for a species that is highly dependent on large-bodied dispersers) and population structure, are not affected by the size of the forest fragment in an old fragmented landscape.
Acknowledgments
We thank Eraldo dos Santos Almeida, Alexandra dos Santos Pires, and Antonio Tavares de Oliveira for help in the field. Financial support was provided by FAPESP (Proc. No. 2005/60788-4), a CNPq fellowship to FAMS (308748/2010-7) and a CAPES fellowship to RCQP. We are grateful to IBAMA for permission to work in their protected areas and to private landowners for permission to work in the fragments.
References
Arroyo-Rodríguez, V., Aguirre, A., Benítez-Malvido, J., & Mandujano, S. (2007). Impact of rain forest fragmentation on the population size of a structurally important palm species: Astrocaryum mexicanum at Los Tuxtlas, Mexico. Biological Conservation, 138, 198-206. [ Links ]
Bruna, E. M. (2003). Are populations in fragmented habitats recruitment limited? Tests with an Amazonian herb. Ecology, 84, 932-947. [ Links ]
Bruna, E. M. & Kress, W. J. (2002). Habitat fragmentation and demographic structure of an Amazonian Under-story herb (Heliconia acuminata). Conservation Biology, 16, 1256-1266. [ Links ]
Bruna, E. M. & Oli, M. K. (2005). Demographic effects of habitat fragmentation on a tropical herb: life-table response experiments. Ecology, 86, 1816-1824. [ Links ]
Bruna, E. M., Fiske, I. J., & Trager, M. D. (2009). Habitat fragmentation and plant populations: is what we know demographically irrelevant? Journal of Vegetation Science, 20, 569-576. [ Links ]
Carvalho, F. A. & Nascimento, M. (2009). Estrutura diamétrica da comunidade e das principais populações arbóreas de um remanescente da floresta atlântica submontana Silva Jardim – RJ, Brasil. Revista Árvore, 33, [ Links ] 327-337.
Chazdon, R. L. (1996). Spatial heterogeneity in tropical forest structure: canopy palms as landscape mosaics. Trends in Ecology and Evolution, 11, 8-9. [ Links ]
Chinchilla, F. (2009). Seed predation by mammals in forest fragments in Monteverde, Costa Rica. Revista de Biología Tropical, 57, 865-877. [ Links ]
Condit, R., Sukumar, R., Hubbell, S. P., & Foster, R. B. (1998). Predicting population trends form size distributions: a direct test in a tropical tree community. The American Naturalist, 152, 495-509. [ Links ]
Dean, W. (1996). A ferr e fogo. A história e a devastação da Mata Atlântica Brasileira. São Paulo: Companhia das Letras. [ Links ]
Eriksson, O. (1996). Regional dynamics of plants: a review of evidence for remnant, source-sink and metapopulations. Oikos, 77, 248-258. [ Links ]
Ewers, R. M. & Didham, R. K. (2006). Confounding factors in the detection of species responses to habitat fragmentation. Biological Reviews, 81, 117-142. [ Links ]
Fantini, A. C. & Guries, R. (2007). Forest structure and productivity of palmiteiro (Euterpe edulis Martius) in the Brazilian Mata Atlântica. Forest Ecology and Management, 242, 185-194. [ Links ]
Fahrig, L. (2002). Effect of habitat fragmentation on the extinction threshold: a synthesis. Ecological Applications, 12, 346-353. [ Links ]
Fischer, J. & Lindenmayer, D. B. (2007). Landscape modification and habitat fragmentation: a synthesis. Global Ecology and Biogeography, 16, 265-280. [ Links ]
Galetti, M. & Aleixo, A. (1998). Effects of palm heart harvesting on avian frugivores in the Atlantic rain forest of Brazil. Journal of Applied Ecology, 35, 286-293. [ Links ]
Galetti, M., Donatti, C. I., Pires, A. S., Guimarães Jr, P. R., & Jordano, P. (2006). Seed survival and dispersal of an endemic Atlantic Forest palm: the combination effects of defaunation and Forest fragmentation. Botanical Journal of Linnean Society, 151, 141-149. [ Links ]
Galetti, M., Guevara, R., Côrtes, M. C., Fadini, R., Von Matter, S., Leite, A. B., Labecca, F., Ribeiro, T., Carvalho, C. S., Collevatti, R. G., Pires, M. M., Guimarães Jr., P. R., Brancalion, P. H., Ribeiro, M. C., & Jordano, P. (2013). Functional extinction of birds drives rapid evolutionary changes in seed size. Science, 340, 1086-1090. [ Links ]
Giulietti, A. N., Harley, R. M., Queiroz, L. P., Wanderley, M. G., & Van den Berg, C. (2005). Biodiversidade e conservação das plantas no Brasil. Megadiversidade, 1, 52-61. [ Links ]
Henderson, A., Galeano, G., & Bernal, R. (1995). Field guide to the palms of the Americas. New Jersey: Princeton University Press. [ Links ]
Jules, E. S. (1998). Habitat fragmentation and demographic change for a common plant: Trillium in old-growth forest. Ecology,79, 1645-1656. [ Links ]
Laurance, W. F. & Bierregaard, R. O. (1997). Tropical forest remnants: ecology, management and conservation of fragmented communities. Chicago, IL: University of Chicago Press. [ Links ]
Lorenzi, H., Souza, H. M., Medeiros-Costa, J. T., Cerqueira, L. S. C., & Ferreira, E. (2004). Palmeiras brasileiras e exóticas cultivadas. Nova Odessa: Plantarum. [ Links ]
Melo, F. P. L., Arroyo-Rodríguez, V., Fahrig, L., Martínez-Ramos, M., & Tabarelli, M. (2013). On the hope for biodiversity-friendly tropical landscapes. Trends in Ecology and Evolution, 1649, 1-7. [ Links ]
Primack, R. B. (1995). A primer of conservation biology. Sunderland: Sinauer Associates. [ Links ]
Portela, R. C. Q. & Santos, F. A. M. (2011). Caracterização dos estádios ontogenéticos de três espécies de palmeiras: uma proposta de padronização para estudos de dinâmica populacional. Revista Brasileira de Botânica, 34, 523-535. [ Links ]
Ranta, P., Blom, T., Niemelã, J., Elina, J., & Sitonem, M. (1998). The fragmented Atlantic rain forest of Brazil: size, shape and distribution of forest fragments. Biodiversity and Conservation, 7, 385-403. [ Links ]
Reis, M. S., Fantini, A. C., Nodari, R. O., Reis, A., Guerra, M. P., & Mantovani, A. (2000). Management and conservation of natural populations in Atlantic Rain Forest: The case study of palm heart (Euterpe edulis Martius). Biotropica, 32, 894-902. [ Links ]
Rizzini, C. T. (1979). Tratado de fitogeografia do Brasil, aspectos sociológicos e florísticos. São Paulo: HUCI-TEC/Ed. Da Universidade de São Paulo. [ Links ]
Rodríguez-Buriticá, S., Orjuela, M. A., & Galeano, G. (2005). Demography and life history of Geonoma orbignyana: an understory palm used as foliage in Colombia. Forest Ecology and Management, 211, 329-340. [ Links ]
Sampaio, M. B. & Scariot, A. (2010). Effects of stochastic herbivory events on population maintenance of an understory palm species (Geonoma schottiana) in riparian tropical forest. Journal of Tropical Ecology, 26, 151-161. [ Links ]
Santos, K., Kinoshita, L. S., & Santos, F. A. M. (2007). Tree species composition and similarity in semideciduous forest fragments of southeastern Brazil. Biological Conservation, 135, 268-277. [ Links ]
Schemske, D. W., Husband, B. C., Ruckelshaus, M. H., Goodwillie, C., Parker, I. M., & Bishop, J. G. (1994). Evaluating approaches to the conservation of rare and endangered plants. Ecology, 75, 584-606. [ Links ]
Silva-Matos, D. M., Freckleton, R. P., & Watkinson, A. R. (1999). The role of density dependence in the population dynamics of a tropical palm. Ecology, 80, 2635-2650. [ Links ]
Souza, A. F. (2007). Ecological interpretation of multiple population size structures in trees: the case of Arau-caria angustifolia in South America. Austral Ecology, 32, 524-533. [ Links ]
Souza, A. F. & Martins, F. R. (2002). Spatial distribution of an undergrowth palm in fragments of the Brazilian Atlantic Forest. Plant Ecology, 164, 141-155. [ Links ]
Souza, A. F. & Martins, F. R. (2004). Population structure and dynamics of a neotropical palm in fire-impacted fragments of the Brazilian Atlantic Forest. Biodiversity and Conservation, 13, 1611-1632. [ Links ]
Tabarelli, M. & Gascon, C. (2005). Lições da pesquisa sobre fragmentação: aperfeiçoando políticas e diretrizes de manejo para a conservação da biodiversidade. Megadiversidade, 1, 181-188. [ Links ]
Walter, H. (1971). Ecology of Tropical and Subtropical Vegetation. Edinburgh: Oliver and Boyd. [ Links ]
Bruna, E. M. (2003). Are populations in fragmented habitats recruitment limited? Tests with an Amazonian herb. Ecology, 84, 932-947. [ Links ]
Bruna, E. M. & Kress, W. J. (2002). Habitat fragmentation and demographic structure of an Amazonian Under-story herb (Heliconia acuminata). Conservation Biology, 16, 1256-1266. [ Links ]
Bruna, E. M. & Oli, M. K. (2005). Demographic effects of habitat fragmentation on a tropical herb: life-table response experiments. Ecology, 86, 1816-1824. [ Links ]
Bruna, E. M., Fiske, I. J., & Trager, M. D. (2009). Habitat fragmentation and plant populations: is what we know demographically irrelevant? Journal of Vegetation Science, 20, 569-576. [ Links ]
Carvalho, F. A. & Nascimento, M. (2009). Estrutura diamétrica da comunidade e das principais populações arbóreas de um remanescente da floresta atlântica submontana Silva Jardim – RJ, Brasil. Revista Árvore, 33, [ Links ] 327-337.
Chazdon, R. L. (1996). Spatial heterogeneity in tropical forest structure: canopy palms as landscape mosaics. Trends in Ecology and Evolution, 11, 8-9. [ Links ]
Chinchilla, F. (2009). Seed predation by mammals in forest fragments in Monteverde, Costa Rica. Revista de Biología Tropical, 57, 865-877. [ Links ]
Condit, R., Sukumar, R., Hubbell, S. P., & Foster, R. B. (1998). Predicting population trends form size distributions: a direct test in a tropical tree community. The American Naturalist, 152, 495-509. [ Links ]
Dean, W. (1996). A ferr e fogo. A história e a devastação da Mata Atlântica Brasileira. São Paulo: Companhia das Letras. [ Links ]
Eriksson, O. (1996). Regional dynamics of plants: a review of evidence for remnant, source-sink and metapopulations. Oikos, 77, 248-258. [ Links ]
Ewers, R. M. & Didham, R. K. (2006). Confounding factors in the detection of species responses to habitat fragmentation. Biological Reviews, 81, 117-142. [ Links ]
Fantini, A. C. & Guries, R. (2007). Forest structure and productivity of palmiteiro (Euterpe edulis Martius) in the Brazilian Mata Atlântica. Forest Ecology and Management, 242, 185-194. [ Links ]
Fahrig, L. (2002). Effect of habitat fragmentation on the extinction threshold: a synthesis. Ecological Applications, 12, 346-353. [ Links ]
Fischer, J. & Lindenmayer, D. B. (2007). Landscape modification and habitat fragmentation: a synthesis. Global Ecology and Biogeography, 16, 265-280. [ Links ]
Galetti, M. & Aleixo, A. (1998). Effects of palm heart harvesting on avian frugivores in the Atlantic rain forest of Brazil. Journal of Applied Ecology, 35, 286-293. [ Links ]
Galetti, M., Donatti, C. I., Pires, A. S., Guimarães Jr, P. R., & Jordano, P. (2006). Seed survival and dispersal of an endemic Atlantic Forest palm: the combination effects of defaunation and Forest fragmentation. Botanical Journal of Linnean Society, 151, 141-149. [ Links ]
Galetti, M., Guevara, R., Côrtes, M. C., Fadini, R., Von Matter, S., Leite, A. B., Labecca, F., Ribeiro, T., Carvalho, C. S., Collevatti, R. G., Pires, M. M., Guimarães Jr., P. R., Brancalion, P. H., Ribeiro, M. C., & Jordano, P. (2013). Functional extinction of birds drives rapid evolutionary changes in seed size. Science, 340, 1086-1090. [ Links ]
Giulietti, A. N., Harley, R. M., Queiroz, L. P., Wanderley, M. G., & Van den Berg, C. (2005). Biodiversidade e conservação das plantas no Brasil. Megadiversidade, 1, 52-61. [ Links ]
Henderson, A., Galeano, G., & Bernal, R. (1995). Field guide to the palms of the Americas. New Jersey: Princeton University Press. [ Links ]
Jules, E. S. (1998). Habitat fragmentation and demographic change for a common plant: Trillium in old-growth forest. Ecology,79, 1645-1656. [ Links ]
Laurance, W. F. & Bierregaard, R. O. (1997). Tropical forest remnants: ecology, management and conservation of fragmented communities. Chicago, IL: University of Chicago Press. [ Links ]
Lorenzi, H., Souza, H. M., Medeiros-Costa, J. T., Cerqueira, L. S. C., & Ferreira, E. (2004). Palmeiras brasileiras e exóticas cultivadas. Nova Odessa: Plantarum. [ Links ]
Melo, F. P. L., Arroyo-Rodríguez, V., Fahrig, L., Martínez-Ramos, M., & Tabarelli, M. (2013). On the hope for biodiversity-friendly tropical landscapes. Trends in Ecology and Evolution, 1649, 1-7. [ Links ]
Primack, R. B. (1995). A primer of conservation biology. Sunderland: Sinauer Associates. [ Links ]
Portela, R. C. Q. & Santos, F. A. M. (2011). Caracterização dos estádios ontogenéticos de três espécies de palmeiras: uma proposta de padronização para estudos de dinâmica populacional. Revista Brasileira de Botânica, 34, 523-535. [ Links ]
Ranta, P., Blom, T., Niemelã, J., Elina, J., & Sitonem, M. (1998). The fragmented Atlantic rain forest of Brazil: size, shape and distribution of forest fragments. Biodiversity and Conservation, 7, 385-403. [ Links ]
Reis, M. S., Fantini, A. C., Nodari, R. O., Reis, A., Guerra, M. P., & Mantovani, A. (2000). Management and conservation of natural populations in Atlantic Rain Forest: The case study of palm heart (Euterpe edulis Martius). Biotropica, 32, 894-902. [ Links ]
Rizzini, C. T. (1979). Tratado de fitogeografia do Brasil, aspectos sociológicos e florísticos. São Paulo: HUCI-TEC/Ed. Da Universidade de São Paulo. [ Links ]
Rodríguez-Buriticá, S., Orjuela, M. A., & Galeano, G. (2005). Demography and life history of Geonoma orbignyana: an understory palm used as foliage in Colombia. Forest Ecology and Management, 211, 329-340. [ Links ]
Sampaio, M. B. & Scariot, A. (2010). Effects of stochastic herbivory events on population maintenance of an understory palm species (Geonoma schottiana) in riparian tropical forest. Journal of Tropical Ecology, 26, 151-161. [ Links ]
Santos, K., Kinoshita, L. S., & Santos, F. A. M. (2007). Tree species composition and similarity in semideciduous forest fragments of southeastern Brazil. Biological Conservation, 135, 268-277. [ Links ]
Schemske, D. W., Husband, B. C., Ruckelshaus, M. H., Goodwillie, C., Parker, I. M., & Bishop, J. G. (1994). Evaluating approaches to the conservation of rare and endangered plants. Ecology, 75, 584-606. [ Links ]
Silva-Matos, D. M., Freckleton, R. P., & Watkinson, A. R. (1999). The role of density dependence in the population dynamics of a tropical palm. Ecology, 80, 2635-2650. [ Links ]
Souza, A. F. (2007). Ecological interpretation of multiple population size structures in trees: the case of Arau-caria angustifolia in South America. Austral Ecology, 32, 524-533. [ Links ]
Souza, A. F. & Martins, F. R. (2002). Spatial distribution of an undergrowth palm in fragments of the Brazilian Atlantic Forest. Plant Ecology, 164, 141-155. [ Links ]
Souza, A. F. & Martins, F. R. (2004). Population structure and dynamics of a neotropical palm in fire-impacted fragments of the Brazilian Atlantic Forest. Biodiversity and Conservation, 13, 1611-1632. [ Links ]
Tabarelli, M. & Gascon, C. (2005). Lições da pesquisa sobre fragmentação: aperfeiçoando políticas e diretrizes de manejo para a conservação da biodiversidade. Megadiversidade, 1, 181-188. [ Links ]
Walter, H. (1971). Ecology of Tropical and Subtropical Vegetation. Edinburgh: Oliver and Boyd. [ Links ]
*Correspondencia a:
Rita de Cássia Quitete Portela. Departamento de Ecologia, Universidade Federal do Rio de Janeiro, Caixa Postal 68020, Cep 21941-590, Rio de Janeiro, Brasil. Correo electrónico rita@quiteteportela.com.br
Flavio Antonio Maes dos Santos. Departamento de Biologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, 13083-970, Campinas, SP, Brasil. Correo electrónico fsantos@unicamp.br
1. Departamento de Ecologia, Universidade Federal do Rio de Janeiro, Caixa Postal 68020, Cep 21941-590, Rio de Janeiro, Brasil. Correo electrónico rita@quiteteportela.com.br
2. Departamento de Biologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, 13083-970, Campinas, SP, Brasil. Correo electrónico fsantos@unicamp.br
Received 14-v-2013. Corrected 10-Xii-2013. Accepted 14-i-2014.


{kind=link}
{kind=link}
{kind=link}
{kind=link}